
黄土高原半干旱区典型草地生态系统CO2交换对刈割的响应
2020-03-13 03:16:08张燕江邱莉萍高海龙魏孝荣张兴昌
生态学报 2020年1期
张燕江,邱莉萍,,*,高海龙,刘 建,魏孝荣,张兴昌
1 西北农林科技大学资源环境学院, 杨凌 712100 2 西北农林科技大学黄土高原土壤侵蚀与旱地农业国家重点实验室, 杨凌 712100 3 西北农林科技大学动物科技学院, 杨凌 712100
草地生态系统与大气的碳交换决定着草地的生产力,并对碳平衡有重要影响,包括土壤呼吸(Rs, soil respiration),生态系统呼吸(Re, ecosystem respiration)和光合3个过程。土壤呼吸是土壤产生并向大气释放CO2的代谢作用[1],是陆地碳收支中最大的通量,约占全球CO2交换量的25%[2]。净生态系统CO2交换(NEE, net ecosystem carbon exchange)是生态系统总初级生产力(GEPG, gross primary productivity)和生态系统呼吸平衡的结果[3],约为总初级生产力的20%[4]。随着气候变化和人为活动的加剧,生态系统碳平衡被打破,导致净生态系统CO2交换降低[5],土壤呼吸释放出更多的CO2[6]。草地作为世界上分布最广的植被类型之一,大约储存着全球碳总量的1/5[7],在调节气候变化中扮演着重要角色,已成为减缓全球气候变化的主要研究内容[8]。目前,草地生态系统碳通量各组分季节性变化特征及其与环境因子之间关系的研究已受到广泛关注[9- 11],但在刈割条件下,黄土高原半干旱区典型草地生态系统碳循环过程及其影响因子的研究较少。
黄土高原半干旱区典型草地是我国主要的草地生态系统之一,受过度放牧及人类生产活动的干扰,草地退化十分严重[12- 13]。近年来,随着草地封育与植被重建措施的实施,该区退化草地得到明显恢复,面积已达6×107hm2,占该区土地面积的32.6%[14]。黄土高原属典型的干旱及半干旱气候,降雨年际差异大、季节性分布不均,该区草地生态系统碳交换对气候变化的响应非常敏感。为提高草地生产力,实现草地资源合理利用,保障生态效益最大化,选择适当的草地管理措施已成为促进草地生态系统可持续发展的关键。刈割作为草地生态系统合理利用的主要措施之一,对生态系统的结构及功能具有一定的调节作用[15- 17]。有研究表明刈割可使高草草原土壤呼吸降低19%—49%[18- 20],而Jia等[21]和Han等[22]发现刈割对我国黄土高原和内蒙古草地的土壤呼吸没有影响,这表明刈割对土壤呼吸的影响与气候条件有关,因此,需要深入分析刈割和气候条件的交互作用。此外,作为生态系统碳通量的重要组成部分,GEP、Re和NEE对刈割响应方面的研究比较少,限制了我们对草地生态系统碳循环过程及其对气候变化反馈的认识。
本研究以宁夏回族自治区固原市云雾山国家自然保护区围封30年的典型草地为对象,设计刈割和对照处理,对土壤呼吸、总初级生产力、生态系统呼吸和净生态系统CO2交换进行了3年的测定,以确定刈割对生态系统碳通量各组分的影响及其与温度和水分的关系,为黄土高原典型草地碳循环的预测及草地生态系统的管理提供科学依据。
1 材料及方法
1.1 研究区概况及试验设计
研究区位于宁夏回族自治区固原市云雾山国家级自然保护区(106°24′—106°28′ E, 36°13′—36°19′ N),海拔1800—2148 m,面积约为6000 hm2。该区为黄土高原典型的半干旱草地生态系统,属暖温带半干旱季风气候,冬季寒冷干燥,夏季炎热少雨,多年平均降雨量为425 mm,7—9月降水占年总量的65%,年均气温6.9℃,月均温最高可达22℃(7月),最低为-14℃(1月)。土壤类型为黄土母质上发育而成的黑垆土和山地灰褐土,土层分布均匀且深厚。植被以旱生草本植物为主,建群种包括长芒草 (Stipabungeana)、大针茅 (S.grandis)、铁杆蒿 (Artemisiavestita)、百里香 (Thymusmongolicus),伴生物种有猪毛蒿 (A.scoparis)、赖草(Leymussecalinus)、星毛委陵菜 (Potentilla)等[23- 24]。
在30年禁牧草地内选取地势相对平坦且植被类型相对一致的地段,设计刈割和未刈割2种处理,每种处理3次重复,共6个小区完全随机排列,小区面积为20 m2(4 m × 5 m),间隔为2 m。刈割是在每年秋季植物枯萎后将小区内所有植物齐地面收割并移出小区,未刈割为对照。
1.2 测定项目和方法
碳通量采用LI- 840红外线分析仪(LI-COR, Lincoln, USA)结合密闭式气室法测定。测定前在每个小区随机永久布置2个直径20 cm的PVC环和2个0.5 m × 0.5 m的PVC框作为测定基座,将其一端砸入土壤5 cm,另一端露出地表7 cm。在2013年7—9月、2014和2015年5—9月,选择晴朗的天气,在9:00—11:00进行Rs、Re、NEE的测定,每个月测定3—5次。Rs观测时,将直径20cm,高50 cm气体收集罩置于PVC环上,保持整个空间密闭,待数据稳定后,开始记录并保存数据,测定时长120 s。NEE的测定是在Rs测定结束并将气室更换为0.5 m × 0.5 m × 0.5 m的透明有机玻璃箱后进行。将玻璃箱放置在PVC框上,保持空间密闭,连续测定CO2浓度,待CO2浓度趋于稳定变化后,记录并保存120 s内的测定数据。NEE测定结束后,抬起箱子使其内部气体与外界大气充分交换,将其再次放置在PVC框上并迅速用不透明黑布遮盖箱体以阻挡光线,当CO2浓度开始稳定上升时,开始记录数据且持续120 s,此时测定结果为Re。GEP根据NEE和Re两者的差值进行计算。两气室内顶端均安装两个小风扇和一个测定箱内气体温度的温度传感器。
太阳辐射、空气温度、0—10 cm土壤温度和土壤水分由微气象系统直接观测,2秒1次,每10 min计算1个均值,所有数据保存在数据采集器(CR1000, Campbell Scientific Inc., Logan, USA)中。根据样地气象站观测数据(图1),该区2013—2015年降雨量分别为410 mm、450 mm和230 mm,与多年平均降雨量(425 mm)相比,2013年和2014年为正常降雨年,而2015年为干旱年。
采用以下公式对生态系统碳通量各组分(Rs,NEE和Re)进行计算,具体方法见[25- 26]。
(1)
式中,F为CO2通量组分 (Rs,NEE和Re, μmol m-2s-1);V是气室体积 (m3);Pav为大气压强 (kPa);Wav是水汽分压 (mmol/mol);R为理想气体常数 (8.314 J mol-1K-1);S表示气室表面积 (m2);Tav为箱室内气温 (℃); dc/dt为气室内CO2浓度随时间变化的斜率。
总初级生产力(GEP)计算公式为:
GEP=-NEE+Re
(2)
1.3 数据分析
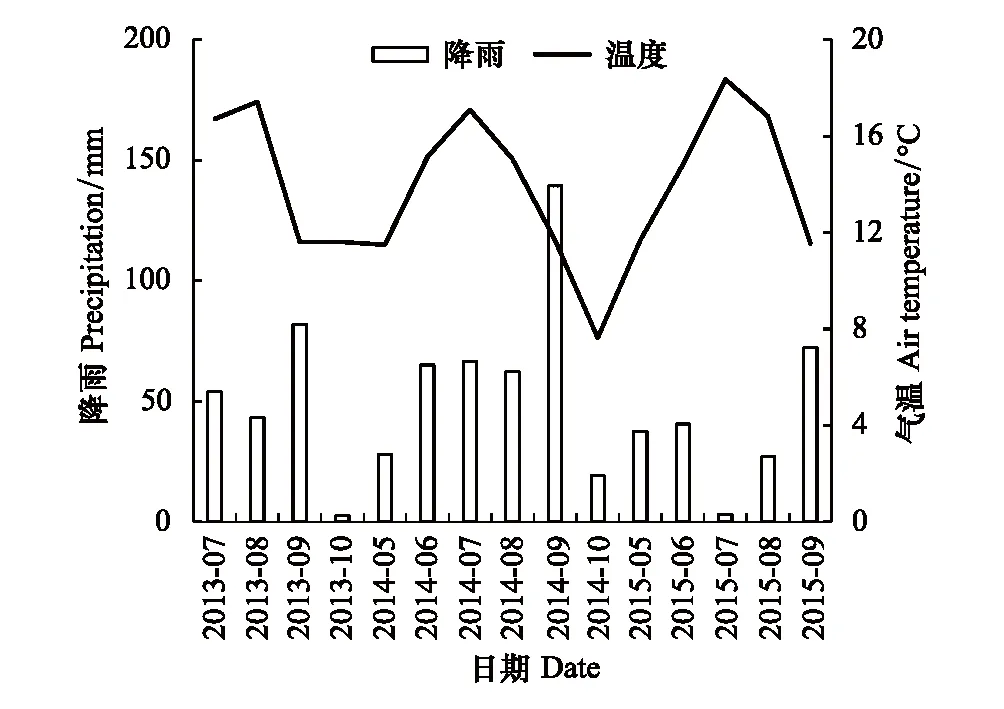
图1 实验期间降雨量(柱)及月平均气温(线)的变化 Fig.1 Variations in monthly precipitation (columns) and mean air temperature (solid line) during experimental period
采用Excel 2016和SPSS 20.0软件对数据进行分析和作图。
2 结果
2.1 土壤温度和水分的变化
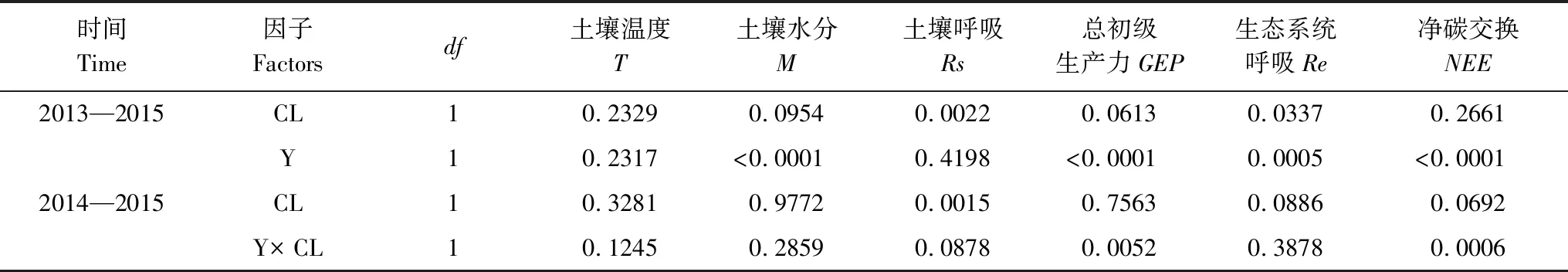
受大气温度的影响,两种处理土壤温度在各生长季均呈显著的单峰模式,以夏季最高,春秋季节最低(图2),但其峰值出现的时间与大小在年际间有所不同,2013和2014年其峰值均出现在8月中旬,其值分别为(24.6±1.0)℃和(23.6±1.0)℃,而2015年峰值出现在7月下旬((21.6±1.5)℃)。刈割、测定年度和两者交互作用对土壤温度的影响不显著(表1)。土壤水分季节性波动非常显著(图2):在2013年7月中旬最高,之后呈缓慢下降的趋势;2014年4月下旬和10月上旬分别出现2个峰值(20.4%±3.7%和24.8%±2.1%),5—10月呈现出波动上升的趋势;2015年全年最高值出现在5月(22.2%±1.9%),而7—9月受长期干旱及间歇性大降雨事件的影响,土壤水分在较高的范围内剧烈波动。刈割对土壤水分的影响与测定年度有关,2013年刈割使土壤水分显著提高了30%(表2),而在2014—2015年土壤水分不受刈割的影响,但受测定年度的影响(表1),与2014年相比,在2015年土壤水分显著降低了17.68%。
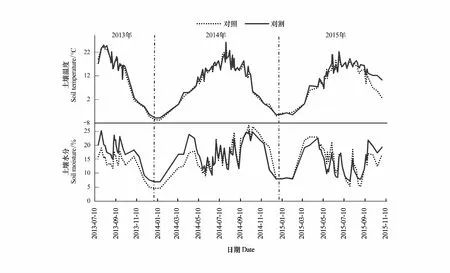
图2 2013年7月至2015年10月间0—10 cm日均土壤温度及水分的季节性变化Fig.2 Seasonal variation in daily mean soil moisture and temperature at 0—10 cm depth under different treatments from July 2013 to October 2015
表1 刈割与测量时间(年)对土壤温度、土壤水分及生态系统CO2通量各组分影响的双因素方差分析结果
Table 1 A two-ANOVA results of clipping and year effects on soil temperature, soil moisture, and ecosystem CO2flux components
时间Time因子Factorsdf土壤温度T土壤水分M土壤呼吸Rs总初级生产力GEP生态系统呼吸Re净碳交换NEE2013—2015CL10.23290.09540.00220.06130.03370.2661Y10.2317<0.00010.4198<0.00010.0005<0.00012014—2015CL10.32810.97720.00150.75630.08860.0692Y× CL10.12450.28590.08780.00520.38780.0006
CL:刈割 Clipping;T:土壤温度 Soil temperature;M:土壤体积含水量 Contents of volumetric moisture;Rs:土壤呼吸 Soil respiration;GEP:总初级生产力Gross primary productivity;NEE:净生态系统CO2交换(负值表示固定CO2) Net primary productivity;Re:生态系统呼吸 Ecosystem respiration;Y:年 Year
2.2 生态系统碳通量各组分的变化
草地生态系统碳交换通量呈现出显著的季节变化特征(图3)。生态系统Rs、GEP、Re和NEE在整个生长季均呈现出单峰曲线的变化模式,以夏季最高,春秋季节最低,但其峰值出现的时间因碳交换组分的不同而异(图3)。土壤和生态系统呼吸在3个测定年度均以7月下旬最高,其值在2013、2014和2015年分别为(4.0±0.1)和(6.7±1.0)、(4.8±0.9) μmol m-2s-1和(9.9±0.8)、(3.6±0.3)和(8.7±0.5) μmol m-2s-1。GEP和NEE在生长季峰值出现的时间则与测定年度有关,GEP的峰值在3个年度分别出现在8月中旬((14.2±3.8) μmol m-2s-1)、7月上旬((18.6±3.8) μmol m-2s-1)和6月中旬((13.6±3.5) μmol m-2s-1),而NEE的峰值则分别出现在9月中旬((-7.9±3.8) μmol m-2s-1)、6月上旬((-11.5±1.6) μmol m-2s-1)和6月下旬((-7.5±4.1) μmol m-2s-1)。

表2 刈割及对照对土壤温度水分及生态系统CO2 通量各组分的影响
每列不同小写字母表示处理之间具有显著性差异,每行不同大写字母表示年际间具有显著性差异(P< 0.01)

图3 2013年7月至2015年10月间生态系统CO2通量各组分的季节性变化Fig.3 Seasonal variation in ecosystem CO2 flux through July 2013 to October 2015 of each treatments Rs:土壤呼吸 Soil respiration;GEP:总初级生产力Gross primary productivity;NEE:净生态系统CO2交换(负值表示固定CO2) Net primary productivity;Re:生态系统呼吸 Ecosystem respiration
本研究中,土壤呼吸不受测定年度的影响,在2014、2015年的平均值分别为(2.3±0.6)μmol m-2s-1和(2.5±0.6) μmol m-2s-1(表2),但受刈割的影响(P=0.0015),而且刈割与测定年度之间的交互作用接近显著水平(P=0.0878)。刈割使土壤呼吸在2014年度增加最大(40%,P=0.032),在2013和2015年度增加较小(5%和8%,P=0.5080和0.2291)。生态系统呼吸在2014年度显著高((6.4±1.3) μmol m-2s-1)于2015年度((5.2±1.5) μmol m-2s-1)(P=0.0005)。刈割降低了生态系统呼吸(P=0.0886),而且这种降低在2014年度较大(12.4%),在2015年度较小(5%)。
生态系统总初级生产力和净气体交换呈现出显著的年际变化特征(P<0.0001),而且受测定年度和刈割交互作用的影响(P=0.0052和0.0006)。总初级生产力和净气体交换量在正常降水年(2014)显著高于干旱年(2015),其值分别为(13.3±3.0)和(-6.9±2.4)、(9.9±2.7)和(-4.7±1.8) μmol m-2s-1。刈割后总初级生产力和净气体交换量在正常降水年显著降低,但是在干旱年显著增加。如刈割后总初级生产力和净气体交换量在2013年分别降低了33%(P<0.001)和38%(P<0.001),在2014年分别降低了10%(P=0.2422)和8%(P=0.5253),在2015年则增加了22%(P=0.013)和63%(P=0.0002)。
2.3 生态系统碳通量各组分与土壤温度和水分的关系
整个实验期间,草地生态系统土壤温度与Rs、GEP和Re的相关性均显著(P<0.01),而与NEE的相关性不显著(P=0.676)。利用线性模型和指数模型分别对不同处理条件下各组分与土壤温度的关系进行拟合(表3),发现指数模型对Rs和Re与温度的拟合关系较好,土壤温度分别解释了Rs和Re季节性变化的31%—46%和39%—56%。基于线性和指数模型对土壤温度与GEP和NEE的拟合不显著,决定系数均在20%以下。土壤水分与Rs、GEP、Re和NEE的二次函数和线性拟合也不显著,决定系数在10%左右(未列出)。

Table 3 生态系统通量各组分与土壤温度的回归分析
RMSE:均方根误差 Root mean square error;Q10:温度敏感性Temperature sensitivity (Q10);SE (Q10): 标准误 Standard error forQ10,SE(Q10)=Q10×10×SE(b)
3 讨论
3.1 生态系统碳通量各组分的年季变化及其影响因素
生态系统碳通量各组分均呈显著的季节性变化特征,这与大多数的研究结果一致[27- 29]。土壤温度和水分是引起碳通量季节性变化的主要原因[8,22]。本研究中,与土壤水分相比,土壤温度很好地解释了Rs和Re的季节性变化,表明在该草地生态系统中土壤温度是引起Rs和Re季节性变化的主要原因。与Re和Rs不同的是,土壤温度和水分对NEE和GEP季节性变化的解释度均不高,其原因可能是NEE和GEP的季节性变化不仅与土壤温度和水分有关,同时也受到光合有效辐射和叶面积指数等多种因子的影响[30- 31]。因此,该生态系统碳通量季节性变化的影响因子与碳通量组分有关,土壤呼吸和生态系统呼吸的季节性变化主要由土壤温度决定。
Rs和Re的峰值出现在7—8月,这与大多数的研究结果一致[19,21],且与土壤温度峰值出现的时间大致相吻合,证明了土壤温度是调节Rs和Re季节变化的主要因子。有研究表明NEE最大值发生在植物生长旺盛期(8月)且与GEP显著相关[32]。本研究中GEP和NEE峰值出现的时间并不同步,且在不同生长季差异很大。适宜的气象条件是植被生长的关键,2013年降雨主要集中在8月,较好的水热条件有利于植物快速生长,使GEP达到峰值,但NEE的最大值却滞后于GEP。2014和2015年生长季前期发生的大降雨事件有利于植物快速返青并积累有机物,使GEP和NEE的峰值提前出现,在干旱年更加明显。这些结果表明降雨的季节分布是引起GEP和NEE季节性变化的主要原因。这与Pierre和Smith[33]发现的由于植物在不同生长阶段对水分的需求不同,生长季特定时期的降水量对生态系统过程的影响大于年降雨量这一观点基本吻合。此外,在干旱和半干旱草地生态系统,降水量和土壤水分有效性是植被生产力的主要限制因子[34],降雨引起生物量变化驱动的RE、GEP和NEE与年降雨量有关[35],干旱导致GEP和NEE降低[36]。如在本研究中,2015年是典型的干旱年,降水量仅为2014年的50%,生长季长期缺水限制了植物的生长,降低了生物量,从而使RE、GEP和NEE在干旱年显著低于正常降水年。
3.2 刈割对生态系统碳通量各组分的影响
刈割是草地生态系统的主要管理措施之一,可通过降低枯落物向微生物的底物供应和光合产物向根系的输送[37],改变群落结构[15]、植物群落补偿性生长[38]和土壤环境等多种直接和间接因素影响生态系统碳通量过程。
本研究结果与郭明英等[39]、Bremer等[18]发现的刈割使土壤呼吸降低这一结论不一致。这种差异可能与不同生态系统土壤温度和水分对刈割的不同响应有关。土壤温度是影响酶和微生物活性、植物生长和有机质分解的关键,对土壤呼吸具有重要作用[40]。增温可激发微生物及其酶活性、加快凋落物和有机质的分解[41],促进地上生物量的生长[42],从而增大土壤呼吸。土壤水分亦可通过改变土壤中溶解性有机碳的含量和迁移能力,影响微生物的利用效率,进而改变土壤呼吸[43]。在受水分限制的生态系统中,随着土壤水分的增加,土壤呼吸显著增加[44]。因此,本研究中,刈割后较高的土壤温度和水分可能是引起土壤呼吸增大的主要原因。此外,在2014年,刈割对土壤呼吸的影响最为显著,其原因可能是:在降雨充沛年,刈割可极大地促进植物的生长,增加组成群落的物种数量[15],甚至引起部分植物的超补偿生长[38],使土壤呼吸进一步提高。
刈割对GEP和Re的影响共同决定了NEE对刈割的响应。本研究中,NEE与GEP显著正相关(对照:NEE=1.48-0.63×GEP,R2=0.78,P<0.0001;刈割:NEE=1.18-0.62×GEP,R2=0.47,P<0.0001),而与Re相关性不显著(R2为0.11—0.06),表明在两种处理下NEE的变化主要受到GEP的直接调控,这一结果与张素彦等[45]发现的典型草地生态系统NEE主要取决于GEP的结论相吻合,而与初小静和韩广轩[46]对中国湿地草地生态系统NEE受ER影响的观点不符,这种差异可能与气候、生态系统类型和土地利用方式有关。在测定期间,刈割增大了Re,但其对GEP和NEE的影响则与降雨年型有关,干旱年为增,正常降雨年为减。这可能与降雨年型和刈割对植被群落结构组成及生长的交互作用有关。不同功能群物种对GEP和NEE贡献差异很大[47],刈割可通过去除土壤表层覆盖物改变土壤环境,使对土壤水分、温度和光照需求较低的物种类型显著增大[15],从而改变GEP和NEE的变化方向和尺度。正常降雨年长芒草主导了碳交换过程,刈割抑制了长芒草的生长,而干旱年长芒草的生长受到限制,其主导的GEP和NEE降低,刈割样地内耐旱性植被具有较高的水分利用性,其在干旱年生长旺盛,成为主导碳交换的植被类型,进而使GEP和NEE对刈割的响应出现年际差异。因此,刈割对GEP和NEE的影响主要取决于不同植被类型对降雨模式的不同响应。
4 结论
本文在不同降雨年型下研究了刈割对生态系统碳通量各组分的影响。结果表明,降雨年际变化是引起GEP、Re和NEE年际差异的主要原因,降雨越多,其值越大。刈割显著提高了土壤呼吸,而刈割对GEP和NEE的影响与降水年型有关,在干旱年增加,正常降雨年减小。
猜你喜欢
农业灾害研究(2022年1期)2022-05-07 01:31:04
辽宁林业科技(2017年4期)2017-06-22 10:23:36
高原山地气象研究(2016年2期)2016-11-10 06:06:27
浙江大学学报(工学版)(2016年2期)2016-06-05 09:20:50
大气科学(2015年5期)2015-12-04 03:04:44
化工进展(2015年6期)2015-11-13 00:26:29
塔里木大学学报(2014年3期)2014-03-11 18:47:27
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:06
自然资源遥感(2014年2期)2014-02-27 11:56:12
自然资源遥感(2014年2期)2014-02-27 11:56:11
