
uc.477+在小鼠B淋巴瘤细胞中的表达研究
2020-03-13 01:50郭俊荣花逸婷孙敏敏朱明明张艳青
实用临床医药杂志 2020年2期
郭俊荣, 张 欣, 花逸婷, 孙敏敏, 朱明明, 华 婧, 张艳青
(扬州大学医学院 江苏省非编码RNA 基础与临床转化重点实验室, 江苏 扬州, 225001)
在人类基因组中,已经发现了481个超保守区域(UCRs), 长度范围为200~779个碱基对。这些UCRs在人类、大鼠和小鼠基因组中高度保守[1-2]。鉴于UCRs的高度保守的特征,推测这些UCRs对哺乳动物细胞具有重要的生物学功能,如一些转录后的超保守区与染色质重构、基因转录及转录后处理有关[3-4], 且发现转录后的超保守RNA在肿瘤组织中差异表达[5]。目前,研究[6-7]报道超保守RNA uc.477+在结直肠癌、前列腺癌中表达异常。此外,研究[8-9]发现长链非编码RNA(lncRNA)与MYC基因家族在肿瘤中存在相互作用。
淋巴瘤是起源于淋巴造血系统的恶性肿瘤,根据瘤细胞分为非霍奇金淋巴瘤(NHL)与霍奇金淋巴瘤(HL), 非霍奇金淋巴瘤占90.0%, 其中临床大多数NHL为B细胞型,占总数70.0%~85.0%。近年来研究[9]表明,原癌基因c-myc过表达是B细胞淋巴瘤的重要分子特征。前期转录组芯片检测发现,当激活含有Myc-雌激素(MycER)融合基因的小鼠B淋巴瘤细胞的Myc时,细胞内uc.477+的表达发生变化,但uc.477+对小鼠B淋巴瘤的影响尚不清楚。本研究探讨uc.477+对B细胞淋巴瘤的作用,现报告如下。
1 材料与方法
1.1 细胞与主要试剂仪器
小鼠B淋巴瘤细胞38B9细胞株、人胚肾细胞293T由本实验室提供,小鼠正常B细胞分选自C57BL/6野生型小鼠脾脏。RPMI、DMEM 高糖培养基、10% 胎牛血清均购自Hyclone 公司。分选B细胞CD19磁珠抗体购自美国BD公司, TRIzol 和LipofectamineTM 2000 均购自Invitrogen 公司,引物和polybrene来自上海生工生物工程技术服务有限公司。PI购自碧云天公司,逆转录试剂盒、荧光定量PCR试剂盒购自TakaRa公司,荧光定量8连管和PCR仪购自罗氏公司。荧光显微镜购自日本Olympus公司, BD FACSVerse流式细胞仪器购自碧迪医疗器械有限公司。细胞培养箱购自美国Thermo公司。
1.2 引物设计
所用引物由Pubmed数据库的Primer-BLAST设计,由上海生工生物工程合成。实时荧光定量PCR引物序列(5′to 3′)如下: ① β-actin上游引物: ACCACACCTTCTACAATGAG, 下游引物: ACGACCAGAGGCATACAG; ② uc.477+上游引物: GACTACAAGACCACCATCTG, 下游引物: TCGGGATGTCCTAGCCATTT。
1.3 磁珠分选小鼠脾脏B细胞
取1只C57BL/6野生型小鼠,颈椎脱臼致死取出脾脏,研磨均匀后经70目尼龙网过滤,离心后加1 mL红细胞裂解液吹打,静置裂解红细胞,洗脱红细胞,加3 mL PBS重悬细胞,再加20 μL CD19磁珠抗体,混匀后4 ℃避光孵育20 min。利用磁珠分选架收集CD19阳性细胞。取200 μL CD19阳性细胞悬液加1 μL CD19流式抗体室温避光孵育20 min, 补加300 μL PBS后流式检测分选率。分选的小鼠B细胞备用。
1.4 荧光定量检测uc.477+在38B9 细胞和
正常B细胞中的表达
TRIzol 提取细胞总RNA, 取1 μg 38B9 细胞和1 μg B细胞的RNA 分别逆转录成cDNA, 5倍稀释待用。cDNA 2 μL, SYBR Ⅱ10 μL, primer F和primer R各0.5 μL, RNase free H2O 7 μL加入8连管,每个样本加3个复孔, β-actin做内参。条件: 变性为95 ℃下100 s, 退火延伸为95 ℃下15 s, 60 ℃下30 s, 40个循环; 溶解曲线为95 ℃下15 s, 60 ℃下30 s, 95 ℃下15 s; 冷却为37 ℃下30 s。
1.5 建立uc.477+过表达38B9细胞株和空载
体38B9细胞株
将uc.477+基因片段构建到pMSCV-PIG逆转录病毒表达载体上,用LipofectamineTM2000 将重组的uc.477-pMSCV-PIG和pMSCV-PIG质粒分别转染至293T 细胞,收36、48、60 h病毒,病毒液用0.45 μm过滤器过滤后置于-80 ℃冰箱备用。病毒液加1‰ polybrene感染38B9细胞,培养48 h后采用0.6 μg/mL嘌呤霉素筛选,获得稳定感染细胞株,荧光显微镜和流式细胞术检测感染率。
1.6 细胞计数和流式检测uc.477+过表达后
38B9细胞株和空载体38B9细胞株的增殖
及细胞周期变化
细胞计数: 将处于对数生长期的uc.477+过表达38B9细胞和空载体38B9细胞分成两组,每组取1×105个细胞接种于60 mm培养皿中,每组取3个复孔,分别计数24、48、72 h每组3个复孔的细胞数,绘制生长曲线,比较增殖趋势。
细胞周期: 2组分别收集1×106个细胞,每组取3个复孔, 75%酒精固定48 h后,加入100 μL RNase、100 μL质量浓度为100 μg/μL的PI, 37 ℃避光孵育30 min, 1 h内流式细胞仪分析细胞周期。
1.7 统计学处理

2 结 果
2.1 磁珠分选小鼠脾脏B细胞
细胞磁珠分选后,将分选的阳性细胞用CD19流式抗体孵育标记,流式细胞仪检测结果显示阳性细胞比例达95.2%, 说明B细胞分选成功。见图1。
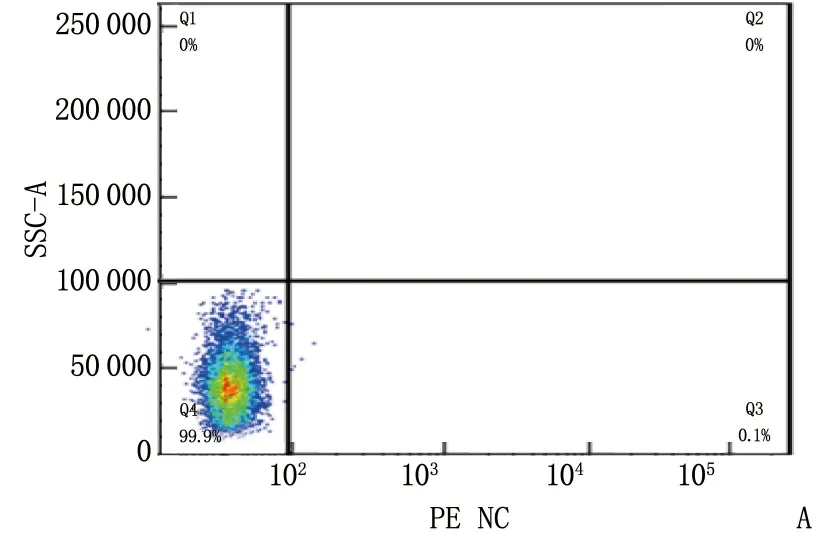
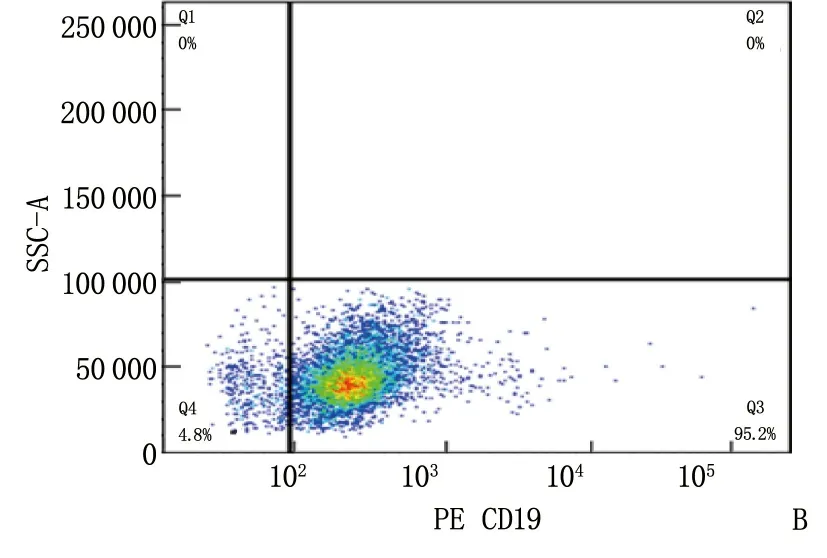
A: B细胞CD19(-); B: B细胞CD19(+)。
图1 流式验证磁珠分选B细胞的分选率
2.2 小鼠B淋巴瘤细胞中uc.477+表达水平
高于小鼠正常B细胞
RT-PCR结果显示,小鼠B淋巴瘤细胞中uc.477+表达水平,高于小鼠正常B细胞,差异有统计学意义(P<0.01)。见图2。
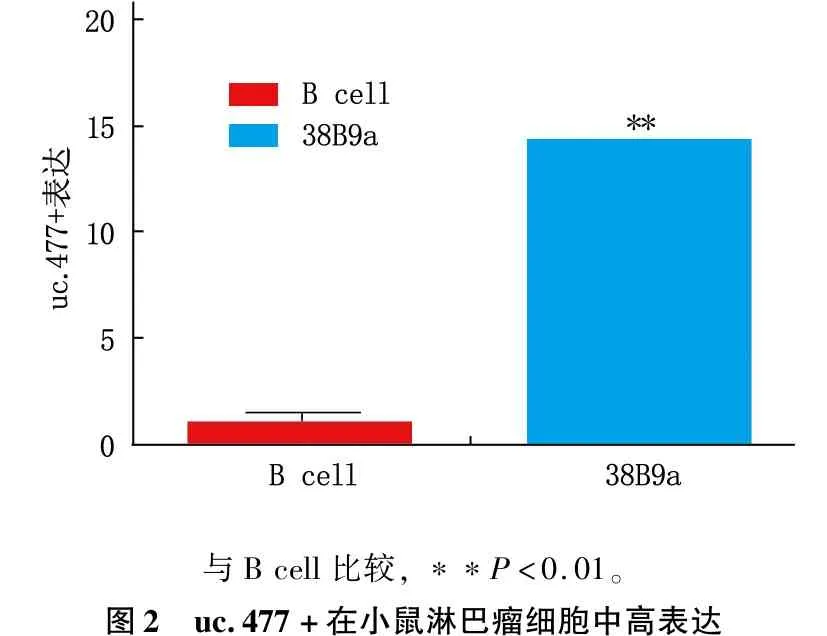
与Bcell比较,∗∗P<0.01。图2 uc.477+在小鼠淋巴瘤细胞中高表达
2.3 逆转录病毒感染38B9细胞感染效率鉴定
将重组的477-pMSCV-PIG和pMSCV-PIG质粒感染38B9细胞,荧光显微镜可见绿色蛋白荧光,见图3; 流式检测感染率分别达91.2%和93.7%, 见图4; 感染后的RT-PCR检测uc.477+的表达量增加,说明成功构建稳转细胞株,见图5。
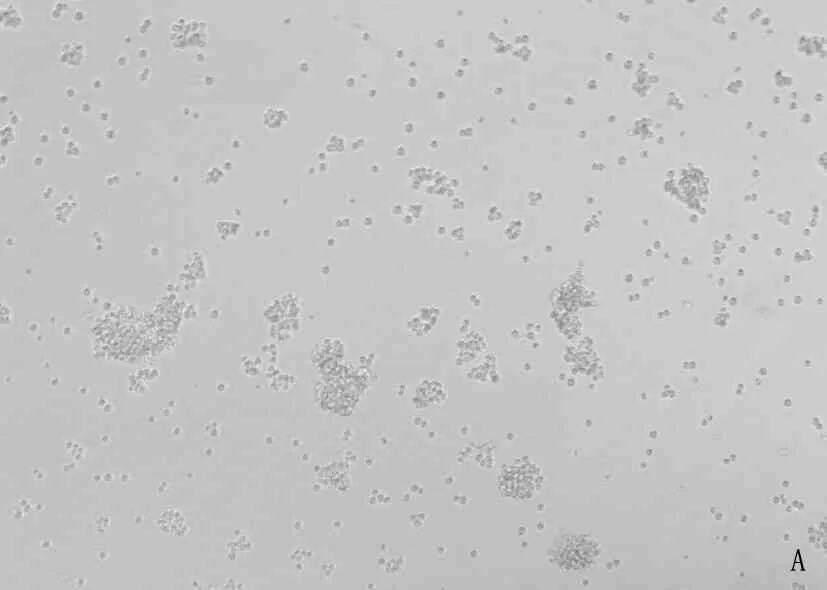
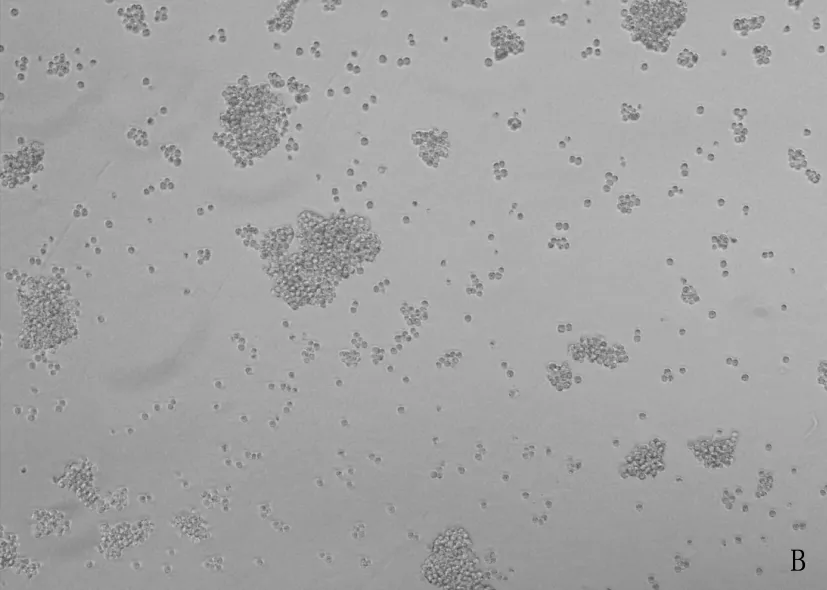
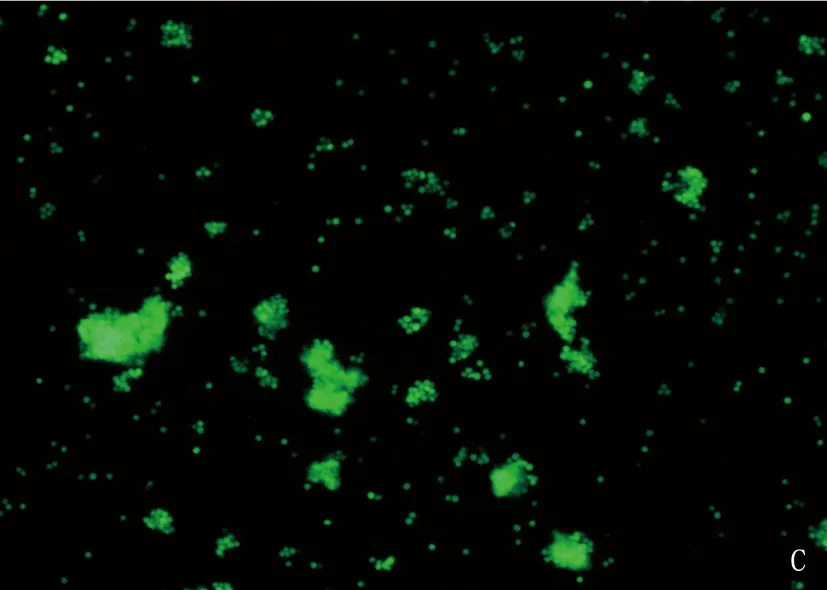
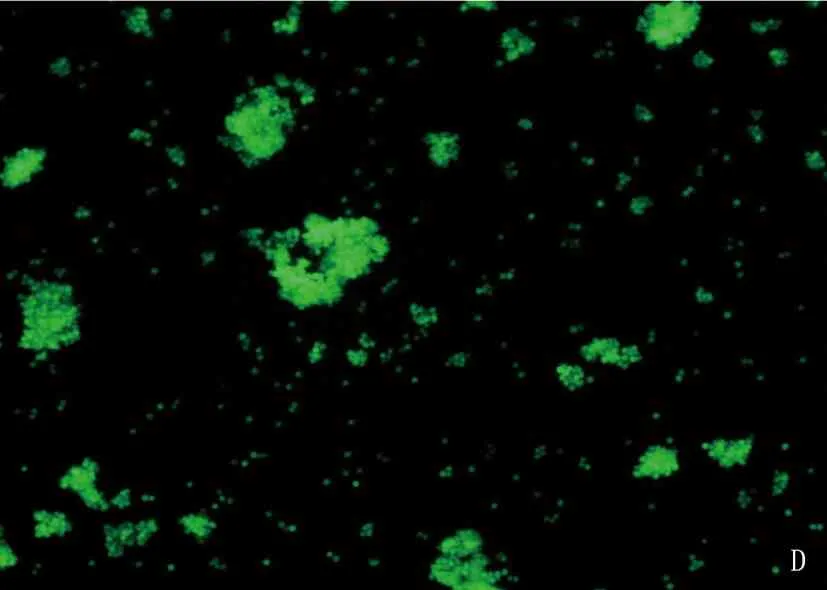
A: 38B9-pMSCV白光图; B: 38B9-uc.477+白光图; C: 38B9-pMSCV荧光图; D: 38B9-uc.477+荧光图。
图3 倒置荧光显微镜观察病毒感染率(放大倍数10倍)
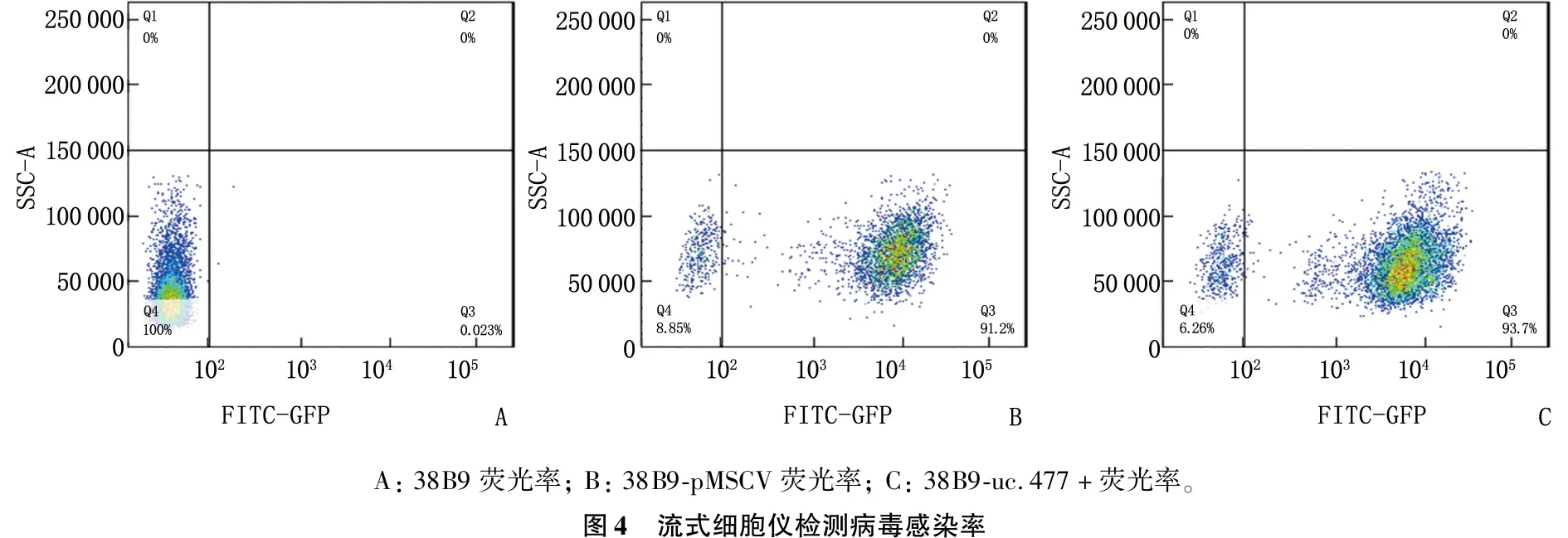
A:38B9荧光率;B:38B9-pMSCV荧光率;C:38B9-uc.477+荧光率。图4 流式细胞仪检测病毒感染率
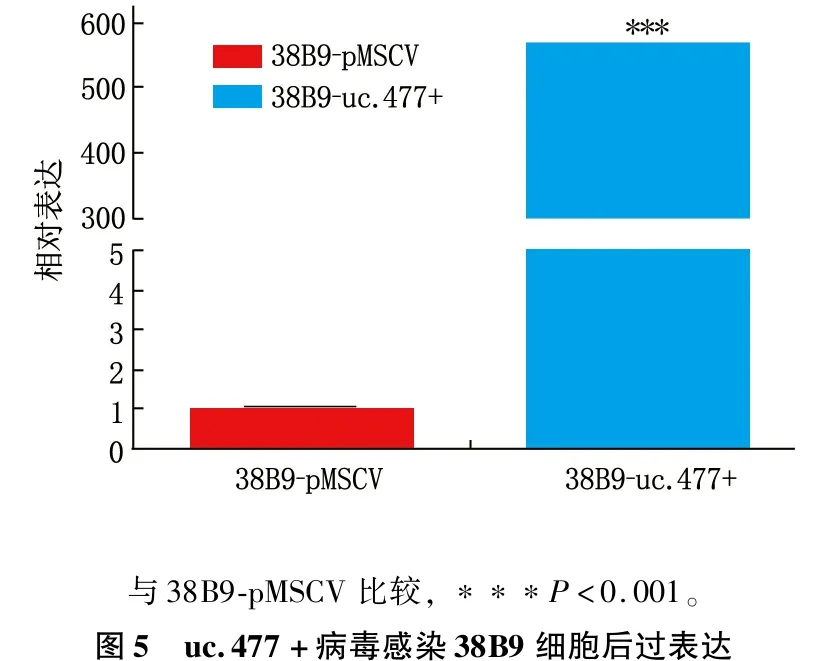
与38B9-pMSCV比较,∗∗∗P<0.001。图5 uc.477+病毒感染38B9细胞后过表达
2.4 uc.477+对38B9细胞增殖及细胞周期的影响
细胞计数实验发现过表达uc.477+的38b9细胞增殖速度比感染空载体38b9增殖速度显著增快(P<0.05或P<0.01)。细胞周期实验发现过表达uc.477+组细胞位于S期和G2期多于MSCV组,说明过表达uc.477+能促进38b9增殖。见图6。
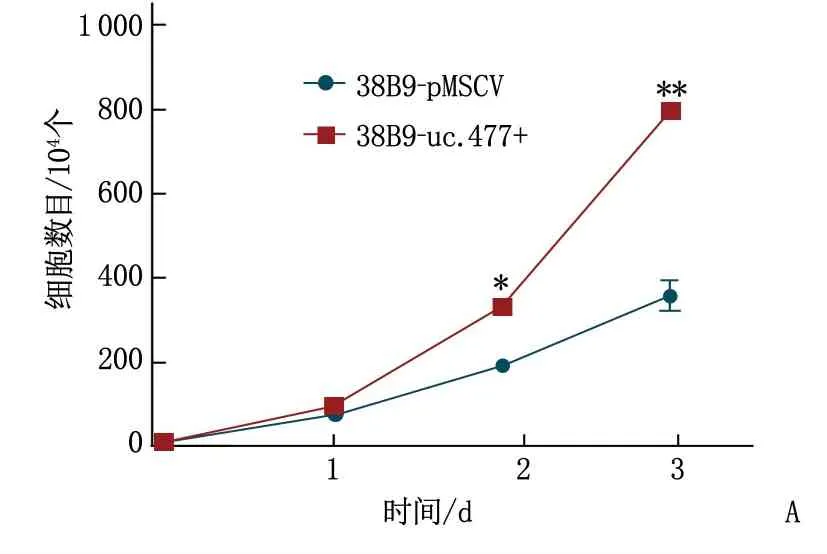

A: 过表达uc.477+后促进38B9细胞生长,与38B9-uc.477+比较,*P<0.05, **P<0.01;B: 过表达uc.477+后细胞S期和G2期比例增加,与38B9-MSCV比较, *P<0.05。
图6 uc.477+对38B9细胞增殖及细胞周期的影响
3 讨 论
研究[10-11]表明,非编码RNA(包括miRs、ucRNAs、lncRNAs、snoRNAs)异常表达会引起基因组的扩增、缺失、突变和表观遗传沉默。因此,非编码RNA在肿瘤发生、发展中起到很关键的作用。超保守区域通常位于癌基因相关的脆弱位点,研究报道T-UCRs的表达水平在人类的胃癌[12]、膀胱癌[13]、肝癌[14]、肺癌[15]、淋巴细胞白血病[16]等多种疾病中均发生显著改变。本研究的超保守RNA uc.477+在结直肠癌[7]、前列腺癌[6]中表达量显著增高。本研究首次检测uc.477+在小鼠普通B细胞与B淋巴瘤细胞中的表达,发现在B淋巴瘤细胞中uc.477+的表达显著增加,提示其可能在B淋巴瘤细胞的发生、发展中发挥一定的作用。
最新研究[17]发现, myc在B细胞淋巴瘤的发生、发展过程中起到非常重要的作用,尤其是源于生发中心的B细胞淋巴瘤,MYC的表达量与生发中心B细胞的分裂能力呈正比,当myc被激活过表达时,会导致B细胞突变成为B细胞淋巴瘤细胞。前期转录组芯片检测发现,当激活含有MycER融合基因的小鼠B淋巴瘤细胞的Myc时,细胞内uc.477+的表达发生变化。通过RT-PCR证实uc.477+在小鼠B淋巴瘤中高表达,这种差异表达提示其可能参与B细胞淋巴瘤的发展,作者通过构建稳定过表达uc.477+的B细胞淋巴瘤细胞系,观察细胞生长及检测其细胞周期变化,证实了uc.477+能促进B细胞淋巴瘤细胞的增殖。但是, uc.477+这种促B细胞淋巴瘤细胞增殖的作用是否参与c-Myc对细胞增殖的调控并不清楚。同时, uc.477+促细胞增殖的效应是通过何种功能蛋白发挥作用也需要进一步研究。
猜你喜欢
现代建筑电气(2022年6期)2022-12-16
计算机时代(2021年9期)2021-10-08
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
中西医结合肝病杂志(2020年2期)2020-10-27
电子技术与软件工程(2020年16期)2020-02-02
商品与质量(2019年27期)2019-12-02
载人航天(2017年1期)2017-07-18
法医学杂志(2016年6期)2017-01-12
医学研究杂志(2015年9期)2015-07-01
