
皆伐前后油松人工林下植物多样性变化及与土壤水分的关系
2020-03-11 05:32:50孙启越张卫强赵连清迟明峰罗桂生郑晓敏贾忠奎王清春
中南林业科技大学学报 2020年3期
孙启越,张卫强,赵连清,迟明峰,罗桂生,郑晓敏,贾忠奎,王清春
(1.北京林业大学 a.林学院;b.国家林业和草原局油松工程技术研究中心;c.生态与自然保护学院,北京 100083; 2.河北省林业调查规划设计院,河北 石家庄 050051;3.河北省平泉市国有黄土梁子林场,河北 平泉 067506)
植物物种多样性是植物群落在物种水平上的多样化,反映了植物群落与生态环境间的相互关系,能体现植物群落的结构类型、组织水平、发展阶段和稳定程度,是生态学中研究最为广泛的多样性层次[1-2]。林下植物是森林生态系统的重要组成部分,具有维持生态平衡和维护地力的重要功能,可以改善人工林的群落结构,改善土壤结构,增加土壤有机质,作为土壤养分库,有利于林分的养分循环,同时一些林下的豆科植物还具有固氮作用[3]。人工林由于群落结构单一和生物多样性低而导致土壤肥力衰退,发展林下植被可以有效缓解这种问题,有研究表明,林下植被对土壤微生物区系和数量有明显影响,可促进土壤酶活性的提高,改善土壤主要化学性质,也就有利于土壤肥力的提高[4],此外林下植物的凋落物较针叶人工林凋落物分解速度快,加快了林分营养元素的归还速度[5]。因此发展林下植物无论在生态上还是培育高质量的林分上都有重要价值。近年来关于植物多样性的研究多是比较不同林分类型的林下植物多样性[6-8],研究林分不同发育阶段林下植物多样性变化的较少;比较不同经营措施如间伐前后的林下植物变化的较多,研究皆伐后林下植物变化的较少。林下物种多样性与林分生长发育阶段密切相关,林下环境随着林分发育过程不断变化,林下植物也会伴随着林分生长发育、人为干扰和环境变化而不断演替[9]。了解不同发育阶段的林下植物多样性特点,通过对森林的科学经营,实施人工抚育以及进行自然恢复,可以保持森林的植物多样性和生态系统的稳定性,维持地力,实现森林的可持续经营[11]。
油松Pinus tabulaeformis为我国华北地区重要的造林树种,在水土保持、木材生产、荒山绿化等方面有重要功能。河北平泉地区是油松人工林的一个重要分布区,油松人工林占当地人工林总面积的61.6%,单层林多,结构简单,当地油松人工林皆伐收获后,依靠油松自身天然更新的能力,在皆伐迹地实现森林的恢复,将来可形成复层异龄林。油松人工林不同发育阶段及皆伐后随时间推移的地力变化是个值得关注的问题,而林下植物是指示地力状况的重要指标,因此有必要开展油松人工林不同发育阶段林下植物多样性动态研究。水是构建和维护生态系统功能的关键因素,物种丰富度与生产力有密切关系,而植物生产力常受水分的限制。土壤作为植物生长的重要基质,其理化特性会对植物产生不同效应,从而影响植物多样性[10]。然而土壤水分对林下植物群落结构和多样性的影响却鲜有人关注。
鉴于此,本研究在河北平泉选取了海拔、坡向、坡度、土层厚度等立地条件一致的处于不同发育阶段的油松人工林及由油松人工林皆伐后依靠天然更新形成的林地,开展林下植物调查,比较皆伐前不同发育阶段和皆伐后不同时间林分中灌木层和草本层植物物种组成和多样性指数的变化规律,用林分环境的阶段性解释林下植物多样性的变化原因,并探讨土壤水分条件与植物多样性的相关性,为更好地发挥油松人工林生态功能,实现林分结构长期稳定性提供理论依据和数据支撑。
1 研究区概况与研究方法
1.1 研究区概况
研究区设在河北省平泉市国有黄土梁子林场,位于河北省东北部,冀蒙交界区域,燕山山脉末端辽河上游,41°10′~41°17′N,118°39′~118°46′E,地形以中低山和丘陵为主,平均海拔660 m。属中温带大陆性干旱季风山地气候,年平均气温6.6 ℃,年平均降水量540 mm,降水主要集中在夏季。年日照2 000~2 900 h,年日照率65%,无霜期120~130 d。本地区土壤类型主要为棕壤土,母质大部分为花岗岩,土壤pH 值约为6.5~7.5。林场辖区森林覆盖率71%,植被主要由落叶阔叶林、针叶林、灌丛和亚高山草甸组成,乔木树种主要有油松、华北落叶松Larix principisrupprechtii、刺 槐Robinia pseudoacacia等,灌 木以胡枝子Lespedeza bicolor、土庄绣线菊Spiraea pubescens、木香薷Elsholtzia stauntonii、圆叶鼠李Rhamnus globosa为主,草本层主要有披针薹草Carex lanceolata、卷柏Selaginella sinensis、白莲蒿Artemisia sacrorum、委陵菜Potentilla aiscolor、龙芽草Agrimonia pilosa等,以旱生中生为主。
1.2 样地设置与调查
2018年5月在对研究区进行全面踏查后,选择地形和土壤条件基本一致的32、40、53年生人工林、伐后5、10、24 a 天然更新林作为研究对象,每种林地设置3个样地,样地面积20 m×20 m,共计18 块样地。其中样地Ⅰ、Ⅱ、Ⅲ为油松一代人工林,分别于1986、1978、1965年春造林,初植密度4 400 株/hm2;样地Ⅳ、Ⅴ、Ⅵ在皆伐前均为50年油松人工纯林,分别于2013、2008、1994年实施皆伐作业,皆伐面积均在5 hm2以上,伐后依靠天然更新在皆伐迹地上恢复成林。样地基本概况见表1。在样地内进行每木检尺,记录乔木株数、树高、胸径、林分郁闭度等,在每个样地四角及中心处设置5个5 m×5 m 的灌木样方,在每个灌木样方内设置1个1 m×1 m 的草本样方,于7月调查记录每个样方内林下灌木(除去油松天然更新)及草本植物的种类、数量、高度、盖度。于5、7、9月在每个样方内用环刀取0~ 20 cm 原状土壤,测定土壤含水量、容重、饱和持水量、毛管持水量、总孔隙度、毛管孔隙度,取3次测定结果平均值作为林地土壤水分数据。

表1 皆伐前后样地概况Table 1 General condition of sample plots
1.3 数据处理
1.3.1 重要值计算
重要值是以综合数值表示植物物种在群落中的相对重要性,其大小是确定优势种的重要依据。林下灌木和草本重要值的计算方法为:

式(1)中,相对多度=(某物种的个体数/所有物种的个体数)×100%;相对频度=(某物种在各样方出现的次数/所有物种出现的总次数)× 100%;相对盖度=(某物种在各样方的盖度之和)/(所有物种在各样方的盖度之和)×100%。
1.3.2 多样性指数和相似系数计算
很多蔬菜的种子表面甚至内部,可能会感染很多的病原菌,如茄子的褐纹病、黄萎病、绵疫病、立枯病、猝倒病;辣椒的绵疫病、立枯病、猝倒病、炭疽病、细菌性斑点病、病毒病;番茄的疮痂病、叶霉病、早疫病、萎蔫病、花叶病;瓜类炭疽病、细菌性角斑病、枯萎病;菜豆炭疽病、叶烧病、锈病、花叶病等。由于带菌的种子又会将病原菌传染给幼苗和成株,从而导致蔬菜病害的发生和蔓延,因此,在播种前进行种子消毒是非常必要的。
采用广泛使用的反映群落物种多样性的Margalef丰富度指数(R)、Simpson多样性指数(D)、Shannon-Wiener 多样性指数(H)及Pielou 均匀度指数(J),计算方法如下:

式(2)、(3)、(4)、(5)中,S为样方内物种数,N为样方内所有物种个体总数,Pi为样方内某个物种的相对多度。
植物群落相似性系数采用Jaccad 系数,计算方法为:

式(6)中,j为两个群落共有的物种数,a和b分别为两个群落各自含有的全部物种数。
1.3.3 数据分析
采用Excel 2010 进行数据的计算和作图,采用SPSS19.0 进行单因素方差分析(One-way ANOVA)、多重比较(LSD)和Pearson 相关性分析,检验不同林地林下植物多样性的差异,分析林下植物多样性与土壤水分物理性质间的相关性。
2 结果与分析
2.1 皆伐前后林下植物物种组成及重要值
2.1.1 灌木层植物
由表2 可以看出,6 种林分样地中共有灌木21 种,隶属14 科20 属,豆科和蔷薇科为优势科,在各样地出现次数较多,在群落发展中起重要作用,其中胡枝子Lespedeza bicolor在所有林分样地中均有出现,优势地位较明显。32年样地(I)林下仅有胡枝子1 种灌木;40年样地(II)林下灌木有7 种,隶属4 科7 属,土庄绣线菊Spiraea pubescens重要值达49.76%,为优势种,其次为胡枝子(24.58%);53年样地(III)林下灌木有14种,隶属13 科14 属,胡枝子(33.74%)为优势种,其次为土庄绣线菊、榆树Ulmus pumila等;伐后5年样地(IV)灌木层植物有6 种,隶属6 科6 属,优势种为胡枝子(66.05%);伐后10年样地(V)灌木层植物有11 种,隶属7 科11 属,胡枝子重要值相对较大,为26.33%,伴生物种有土庄绣线菊、山杨Populus davidiana、榆树等;伐后24年样地(VI)灌木层植物有5 种,隶属3 科5 属,胡枝子重要值达50.27%,为优势种。林下灌木层物种数在各样地表现为53 a >伐后10 a >40 a >伐后5 a >伐后24 a >32 a,不同林分样地灌木层物种组成不尽相同,53年人工林和伐后10年天然更新林样地灌木层物种数较多,且优势种的重要值低于40%,各物种分布较均匀。
2.1.2 草本层植物
由表3 可以看出,研究区内林下草本植物种类较丰富,6 种林分样地中共出现草本植物65 种,隶属28 科59 属,菊科物种数最多,出现植物18种,其次为蔷薇科和禾本科,出现植物6 种和4种。白莲蒿Artemisia sacrorum和委陵菜Potentilla chinensis在所有林分样地中均出现,披针薹草Carex lancifolia、马唐Digitaria sanguinalis、卷 柏Selaginella tamariscina、大丁草Gerbera anandria、龙芽草Agrimonia pilosa在5 种林分样地中出现,除卷柏外均为上述3 科所属物种,说明菊科、蔷薇科和禾本科为研究区草本植物优势科,分布较广、适应性较强。32年样地(I)林下共有草本植物16 种,隶属11 科15 属,以披针薹草为优势种,重要值达36.84%;40年样地(II)林下有草本植物14 种,隶属9 科14 属,优势种为卷柏(40.76%),其次为披针薹草(19.43%);53年样地(III)林下有草本植物34 种,隶属17科31 属,卷柏、小红菊Dendranthema chanetii、披针薹草等物种重要值较其他物种大,为相对优势种;伐后5年样地(IV)草本层植物有20 种,隶属11 科18 属,以沿阶草Ophiopogon bodinieri、白茅Imperata cylindrica、委陵菜等物种相对居多,重要值相对较大;伐后10年样地(V)草本层植物有30 种,隶属14 科28 属,披针薹草相对较多,其次为老鹳草Geranium wilfordii和卷柏;伐后24年样地(VI)草本层植物有31 种,隶属17 科29 属,以披针薹草和马唐为相对优势种,重要值分别为15.82%和12.10%。林下草本层物种数在各样地表现为53 a >伐后10 a >伐后24 a >伐后5 a > 40 a >32 a,皆伐前随林龄增加物种丰富度逐渐增加,皆伐后物种丰富度先降低后增加,这可能与油松天然更新实现植被恢复有关。不同林分样地草本层物种组成差异较大,53年人工林、伐后5年和伐后10年天然更新林样地草本层物种数较多,且优势种重要值均低于20%,优势不明显,说明物种间处于竞争阶段,没有物种占据绝对优势,物种分布趋于均匀。
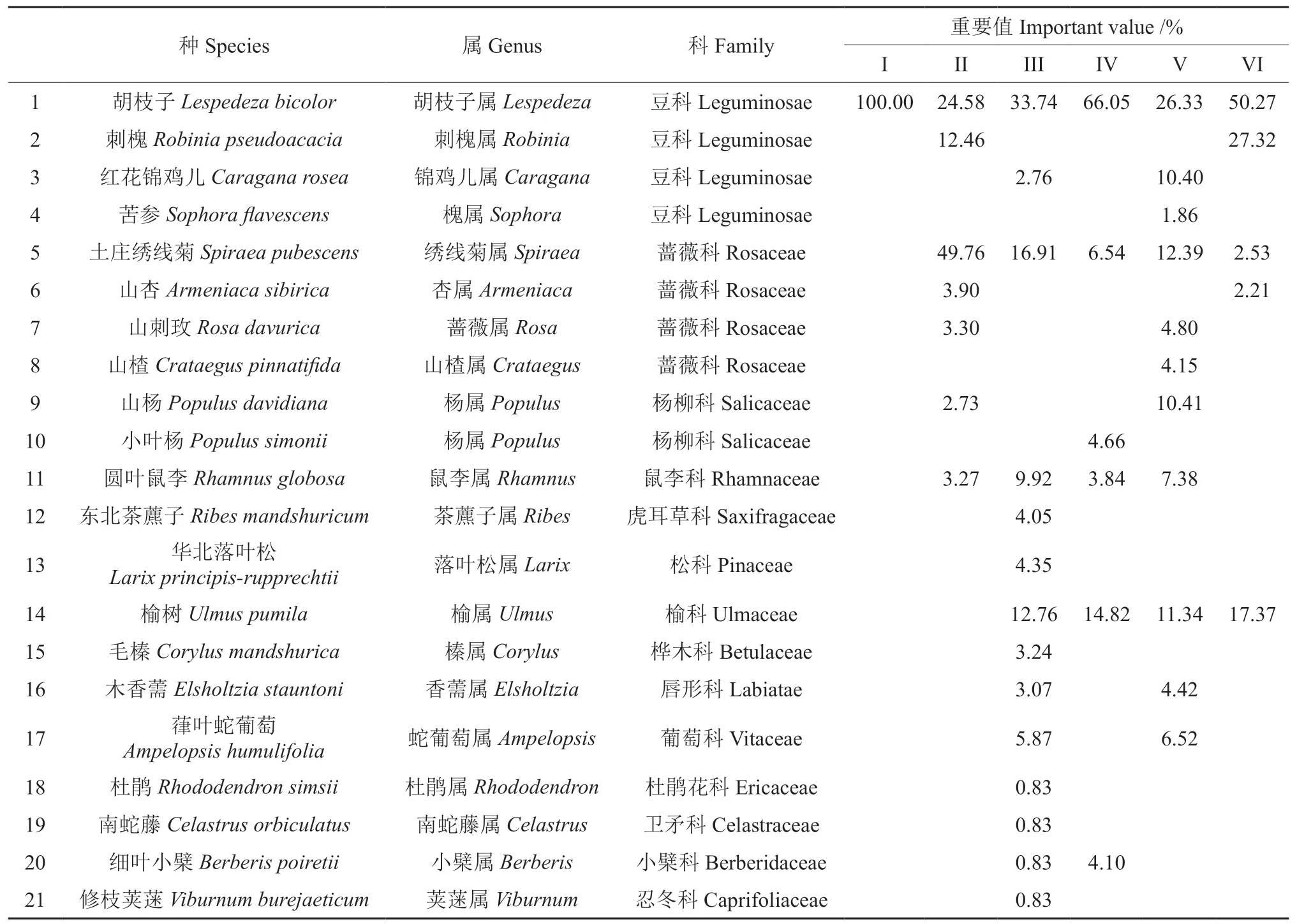
表2 皆伐前后林下灌木层物种组成及重要值Table 2 Species composition and important values of shrub layer before and after clearcutting

表3 皆伐前后林下草本层物种重要值Table 3 Species composition and important values of herb layer before and after clearcutting
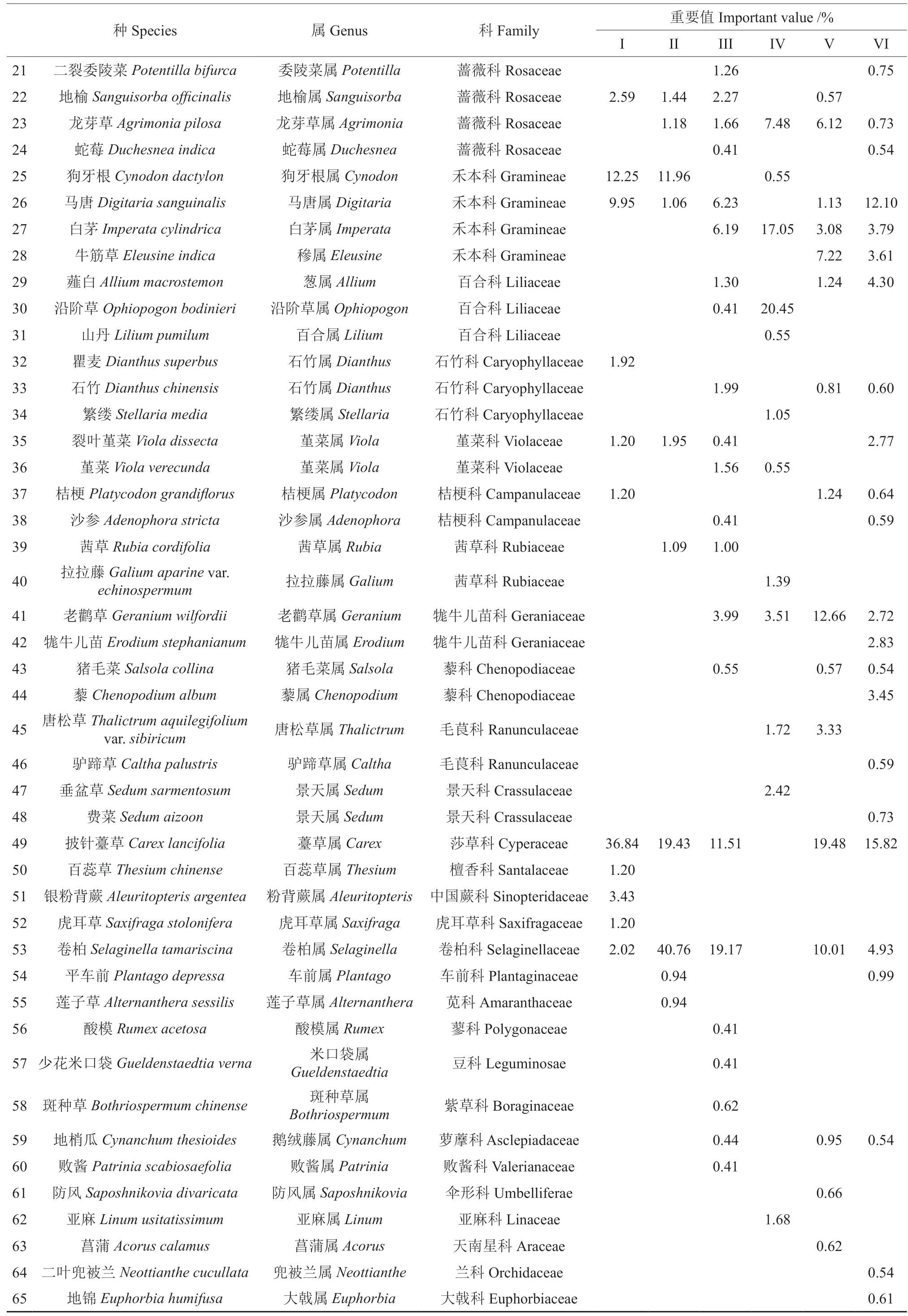
续表3Continuation of table 3
2.2 皆伐前后林下植物多样性
通过对6 种林分样地的4 种多样性指数进行方差分析可以看出(图1),不同林分样地林下植物多样性差异较大,灌木层与草本层的变化规律也不尽相同。32年灌木层仅有胡枝子1 种植物,物种丰富度较差,无法计算其多样性指数,其余样地灌木层中Margalef 丰富度指数、Simpson 多样性指数和Shannon-Wiener 多样性指数均表现为伐后10 a >53 a >40 a >伐后20 a >伐后5 a,且伐后10年和53年样地多样性指数显著大于其他林分样地(P<0.05)。灌木层的Pielou 均匀度指数表现为伐后10 a >40 a >53 a >伐后20 a > 伐后5 a,且伐后10年、40年和53年样地与其他样地差异显著(P<0.05)。伐后10年灌木层Margalef 丰富度指数、Simpson 多样性指数、Shannon-Wiener 多样性指数和Pielou 均匀度指数分别为1.40、0.61、1.24 和0.74,比伐后5年分别增长了240.9%、204.1%、275.8%和71.3%。
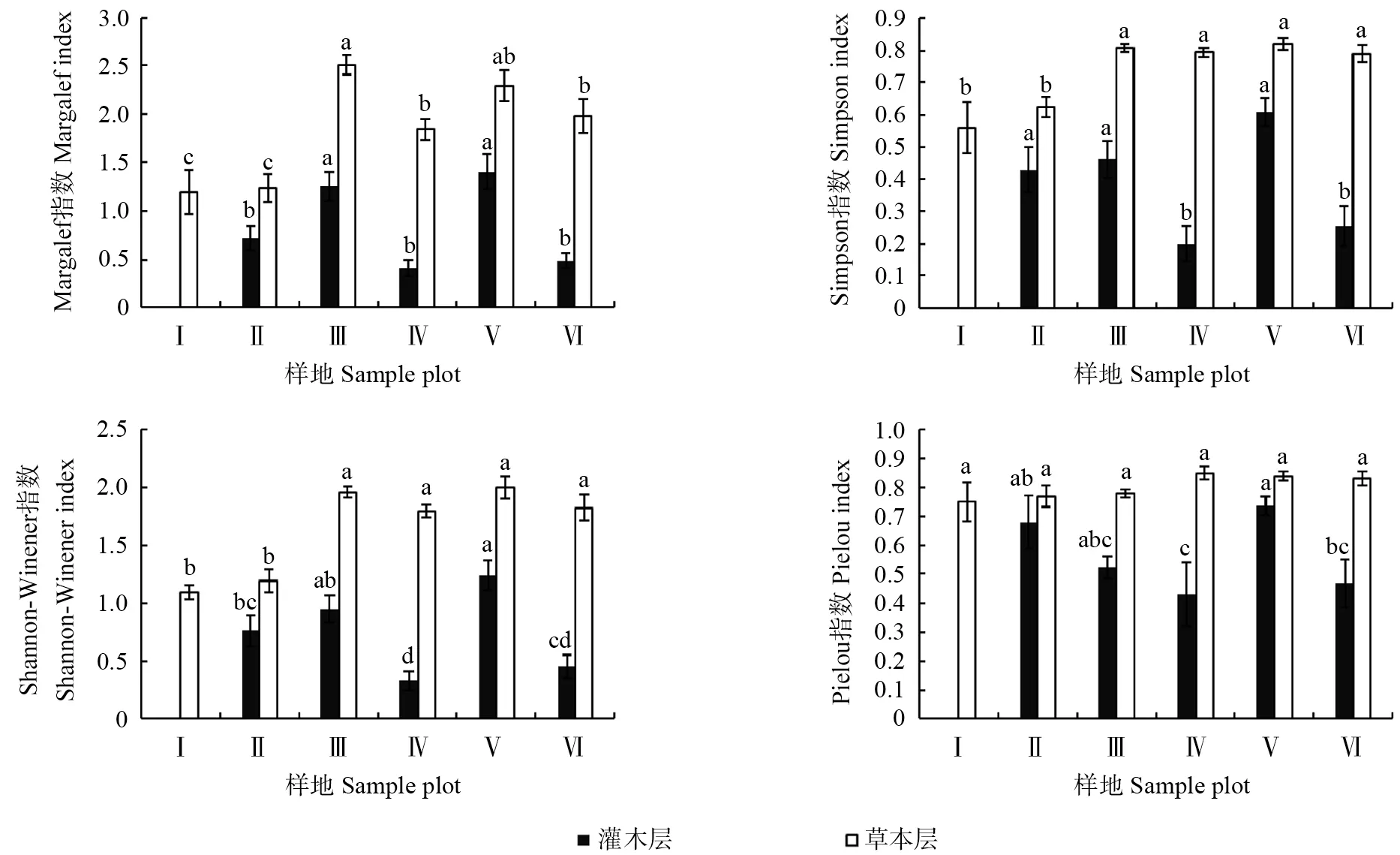
图1 皆伐前后林下植物多样性Fig.1 Understory plant diversity before and after clearcutting
草本层中不同林分样地植物多样性差异同样较为显著,且不同指数的变化规律不同:53年、伐后10年样地的Margalef 指数显著高于其他样地(P<0.05),分别达到2.51 和2.29。32年和40年样地的Margalef 指数最低,分别为1.19 和1.24。53年、伐后5年、伐后10年、伐后24年样地间Simpson 指数和Shannon-Wiener 指数差异均不显著(P>0.05),但均显著高于32年和40年样地 (P<0.05)。不同样地间草本层的Pielou 均匀度指数差异不显著(P>0.05)。
从上述分析可以发现,在皆伐前样地中53年样地多样性最好,32年和40年样地林下植物多样性相对较差;在实施皆伐后植物多样性指数显著降低,伐后10 a 与伐后5 a 相比多样性增长较大,并与53年样地间差异不显著(P>0.05),随时间推移至伐后24年林下植物多样性又出现一定程度降低。从32 a 到伐后24 a 大体呈现出先增加后降低再增加再降低的双峰变化,峰值出现在53 a和伐后10 a,且灌木层的变化幅度较大。此外不同林分样地灌木层与草本层之间林下植物多样性指数差异较明显,均表现为草本层>灌木层,说明草本层的物种更加丰富。
2.3 皆伐前后林下植物群落相似性
群落相似性可以通过分析两个植物群落共有的物种组成特征反映两个群落间的相似程度。由表4 可以看出,不同林地间植物群落相似性有较大差异,其中53年样地(III)与伐后10年(V)、伐后24年样地(VI)相似度较高,相似系数分别达到0.459、0.400,伐后10年(V)和伐后24年(VI)样地相似度也较高,达到0.400,其他样地间相似性较低。伐前样地间物种组成差异较大,可能是因为随着林龄增加林内环境不断发生变化,物种组成也发生较大变化,伐后5年(IV)与其他样地差异明显的原因可能与皆伐对林内环境造成的强度干扰有关。

表4 不同林地植物群落的相似系数Table 4 Similarity coefficient of plant communities before and after clearcutting
2.4 林下植物多样性与土壤水分的相关性
土壤对植物的生长起到重要作用,而土壤水分作为一个重要的环境因素对植物群落的组成和结果有直接影响。表5 为不同林分样地的土壤水分物理性质,可以看出不同林分土壤水分物理性质差异显著(P<0.05)。总体来看伐前32年(I)到53年(III)土壤水分物理性质逐渐改善,伐后5年(IV)到10年(V)仍保持较高水平,伐后24年(VI)出现一定下降。对4 种多样性指数与土壤水分物理性质进行了Pearson 相关性分析,结果表明不同林分林下植物多样性与土壤水分之间存在显著相关性(表6)。灌木层的Margalef指数与土壤水分物理性质各指标均有显著相关性 (P<0.05),Simpson 指数、Shannon-Wiener 指数、Pielou 指数与土壤容重、饱和持水量、毛管持水量、总孔隙度呈显著相关关系(P<0.05)。草本层的Margalef 指数、Simpson 指数、Shannon-Wiener 指数与土壤水分各指标达到极显著相关关系(P<0.01),Pielou 指数与土壤水分的相关性不显著(P>0.05)。总体来看土壤水分条件的差异对林下植物多样性产生了显著影响,较多的土壤水分可以促进植物多样性的提高,且草本层植物多样性对土壤水分条件差异的响应要大于灌木层,这可能与林下植被的根系作用有关。
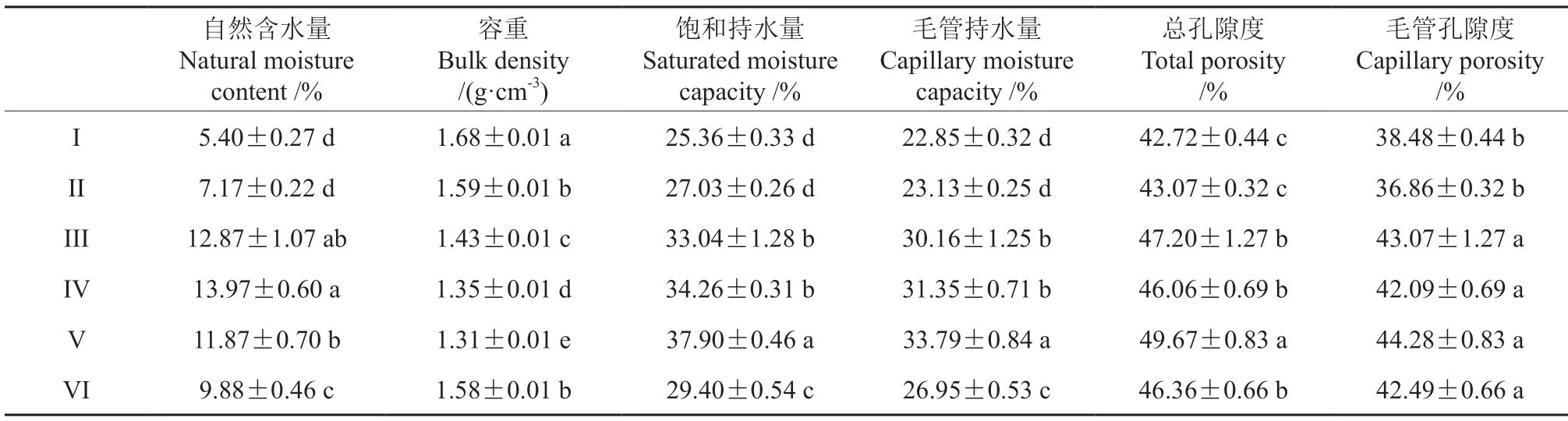
表5 皆伐前后土壤水分物理性质变化Table 5 Variation of soil moisture before and after clearcutting

表6 植物多样性与土壤水分间的相关性†Table 6 Correlations between plant diversity and soil moisture
3 结论与讨论
本研究中,伐前成熟林(53 a)林下物种数量远大于中龄林(32 a)和近熟林(40 a),说明随着林龄增加物种丰富度逐渐增加,这与不同发育阶段林分环境的差异有关。赵耀等[12]关于晋西黄土区不同林地植物多样性的研究表明林分郁闭度对林下植物多样性的影响十分显著,揭示出光照条件是决定林下植物发展的重要因素。本研究中龄林(32 a)林分密度较大,郁闭度高达70%,光照条件较差,林下植物发育也较差,灌木层仅有胡枝子一种植物。马履一等[13]和王凯等[14]关于间伐对油松人工林下植物多样性的影响研究表明,间伐可以明显改善各林龄林分林下植物状况,随间伐强度增大,林下植物多样性和生物量逐渐增加,但间伐的影响不具有长期性,确定合理的间伐周期有利于促进林分结构稳定、生物多样性丰富。本研究中,从中龄林(32 a)到近熟林(40 a)再到成熟林(53 a),林分密度从2 500株/hm2到1 500 株/hm2再到900 株/hm2,间伐强度较大,林分郁闭度保持稳定,调整了林木间关系,在一定程度上改善了光照等林分生长环境,满足了林下植物的生长需要,因此灌木层和草本层植物逐渐发展[15-16]。
皆伐后初期林下物种数相比成熟林显著降低,这是因为皆伐这种强度人为干扰对原有植物造成了较大破坏,皆伐后油松作为先锋树种率先天然更新,压制了其他物种的发育。随着时间推移林下植物种类逐渐增多,这可能是因为油松基本在皆伐迹地上实现天然更新,此时土壤养分状况较好,林地内没有高大乔木遮挡,形成了有利于植物发展的环境。本研究显示在皆伐后24年林下植物种类又出现减少,这种物种丰富度先增加后降低的现象与密度增加导致的物种竞争有关[17-18],油松天然更新密度增大会导致林分所需的营养空间减小,随之出现了较为严重的自然稀疏现象,灌木种类明显减少,灌木均匀度也出现下降。此外林分逐渐郁闭也不利于灌木层多样性的增加。丁继伟等[19]的研究表明灌木植物对林分密度的变化较为敏感,本研究皆伐后灌木层多样性随时间变化幅度较大,与其研究结果较一致。有研究认为采伐后林下尤其是草本层植物多样性会增加[20-21], 本研究中皆伐后草本层多样性一直保持在较高水平,可能是皆伐后林地环境较均一,且林地水热条件有利于草本植物发展。合理的森林恢复方式对植物多样性的提高是有利的,王芸等[22]的研究表明自然恢复的森林植物群落多样性更高,Boan等[23]的研究也认为采伐后自然更新的林分植物丰富度较高,本研究中皆伐后依靠天然更新恢复森林,在伐后10年林下植物多样性达到峰值,与成熟林差异不显著,随后由于物种竞争导致多样性出现下降。因此对于密度过大的林地要及时进行抚育,对于过疏林分还可人工补植阔叶树种,调整林分结构,改善林内环境,加速林下多物种稳定结构的形成[24]。
本研究中胡枝子在各个林地灌木层中均有分布,在皆伐后初期优势度明显,是该地区植物恢复过程中最先广泛自然定居的灌木种,且成熟林和采伐10 a以后林分林下出现刺槐和红花锦鸡儿,这些豆科植物根部常用根瘤菌共生,可有效改良土壤,增加土壤肥力。各个林地草本层中菊科拥有物种数量最多,分布最为广泛,说明菊科植物在研究区内具有较强的适应性,为各个植物群落的重要组成部分。皆伐后初期林地草本层以禾本科、百合科等单子叶植物为主,随后菊科、蔷薇科植物开始进入群落,草本层植物多样性逐渐发展。本研究中草本层出现的物种数量远大于灌木层,其物种组成的变化大于灌木层的变化,这与吴文等[25]的研究结果一致,原因可能是草本层植物对环境条件变化的响应更加敏感。有的研究表明不同林分类型林下植物多样性表现为灌木层>草本层[22],而有的研究表明不同林分林下多样性表现为草本层>灌木层[26],本研究6 种不同发育阶段林分林下植物多样性均表现为草本层>灌木层,这说明林下物种组成和多样性与树种组成、林分环境、人为干扰等因素有关,不同森林类型的灌木层和草本层植物多样性存在差异,不同群落演替过程表现出不同的林下物种组成和分布。
植物多样性可以较好地表征群落组成结构,同时也能对林内环境状况做出指示。本研究选取各发育阶段样地处于同一立地类型,海拔、坡向等林分因子一致,重点考虑土壤水分对林下植物多样性的作用。Deng 等[27]的研究表明植物多样性与土壤含水量呈显著正相关,郝文芳等[28]的研究也表明土壤含水量是影响植物群落组成和多样性最关键的生态因子。本研究中,土壤水分物理性质与灌木层植物多样性相关性显著,与草本层植物多样性相关性极显著,林下植物多样性变化与土壤水分密切相关,土壤水分含量越高,植物多样性越大,同时林下植被的根系可以疏松土壤,改善土壤结构、增加通气性和透水性[29],且丰富的植物种类增加了地表凋落物覆盖,可减少土壤水分的蒸发。因此土壤水分是该地区植被恢复的关键环境因子,良好的土壤水分条件与林下植物的发展可以相互促进。
本研究也存在一定局限性:最早于平泉种植的一代油松林于1994年皆伐收获,至调查时为伐后24年的天然更新林,随着时间推移天然更新林下植物多样性变化有待于进一步研究;影响物种多样性的环境因子比较复杂,本文重点分析了土壤水分变化的影响,由间伐等经营措施造成的林分环境异质性也可作为一个研究方向,如林内光照强度的变化等;此外林下植物可改善土壤肥力,可对林地土壤养分和生物活性进行长期监测,建立其与林下植物相互作用的关系。
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
山西林业(2021年2期)2021-07-21 07:29:32
少儿美术(2020年8期)2020-11-06 05:21:38
防护林科技(2020年6期)2020-08-12 13:34:40
绿色科技(2019年6期)2019-04-12 05:38:42
绿色科技(2019年6期)2019-04-12 05:38:42
现代园艺(2017年22期)2018-01-19 05:06:56
山西农经(2015年7期)2015-07-10 15:00:57
亚热带资源与环境学报(2015年3期)2015-01-22 07:05:36
