
獐子岛岩相潮间带大型海藻有机碳含量及δ13C值的季节变化特征
2020-03-09 00:56:22张靖凡蔡恒江赵玥茹陈文翰胡思琪刘长发
海洋科学 2020年2期
张靖凡, 蔡恒江, 2, 赵玥茹, 陈文翰, 胡思琪, 刘 远, 2, 刘长发, 2
獐子岛岩相潮间带大型海藻有机碳含量及13C值的季节变化特征
张靖凡1, 蔡恒江1, 2, 赵玥茹1, 陈文翰1, 胡思琪1, 刘 远1, 2, 刘长发1, 2
(1. 大连海洋大学 海洋科技与环境学院, 辽宁 大连 116023; 2. 辽宁省高校近岸海洋环境科学与技术重点实验室, 辽宁 大连 116023)
为研究岩相潮间带大型海藻有机碳含量及13C值的季节变化特征, 分别于2016年11月(秋)和2017年2月(冬)、5月(春)、8月(夏)对獐子岛岩相潮间带(39°01′E, 122°43′N)的大型海藻进行调查, 并对其有机碳含量和13C值进行分析。结果表明: 共鉴定出大型海藻3门49种, 其中红藻门24种, 占总数的48.98%; 褐藻门17种, 占总数的34.69%; 绿藻门8种, 占总数的16.33%。大型海藻种类数为春季(35种)>冬季(24种)=夏季(24种)>秋季(23种)。不同种类海藻体内的有机碳含量为15.54%~35.03%,13C值在–33.42‰~–7.43‰之间变动。不同季节海藻体内有机碳含量为冬季>春季>夏季>秋季,13C值为夏季>春季>冬季>秋季。
大型海藻; 种类; 季节变化; 有机碳;13C值
大型海藻是海洋潮间带生态系统中最重要的初级生产者之一, 是最具有经济价值的藻类[1], 为各种海洋生物提供栖息、索饵和产卵的场所, 同时对近岸海洋生态环境也存在着巨大的影响[2]。研究表明, 它们不仅对近岸水体的富营养化有一定的修复作用, 而且部分大型海藻还可以通过生物克生作用来防控有害赤潮生物的生长[3-5]。但也有些大型海藻会破坏近岸海洋生态环境, 如绿潮的爆发[6-7]。
近年来由于人类活动的影响, 导致了大气CO2浓度的不断升高, 对生态环境造成了一系列的影响, 如温室效应、海水酸化等[8]。由于大型藻类可通过光合作用将溶解无机碳和CO2转化为有机碳, 促使空气中CO2向海水中转移, 导致海水中CO2溶解量降低, 因此大型海藻的碳汇效应正逐渐受到关注[9-10]。目前大型海藻碳汇研究多集中在养殖或人工构建的藻场研究[11]。潮间带大型海藻种类繁多, 分布不均, 研究难度较大, 所以针对其碳汇研究少见报道。不同种类大型海藻光合作用的碳同化机制也存在差别, 外来碳源的途径主要有两种, 分别为CO2和HCO3–。研究表明, 大型海藻在光合作用中的碳同化机制可以通过分析其体内的13C值来确定[12-13]。
獐子岛地处北黄海辽东半岛南部, 岛屿海岸线迂回曲折, 多为岩石海岸, 是海藻生长栖息的良好场所, 已经成为黄海北部海藻资源最为丰富的海域之一[14]。獐子岛是我国海参、鲍鱼等经济生物的重要繁殖基地, 大型海藻即可作为海参、鲍鱼这些海珍品的饲料源, 也可作为食品、医药和工业等领域的重要原料。由于自然地理隔离及完善的管理, 岛上海岸线人为影响较少, 为对大型海藻的科学研究调查提供了良好的样本条件。
本文在对獐子岛岩相潮间带大型海藻群落结构调查的基础上, 对不同种类海藻体内有机碳含量和13C值季节变化进行分析, 旨在为大型海藻碳汇效应的估算及碳同化机制的研究提供依据。
1 材料和方法
1.1 采样点和采样方法
分别于2016年11月(秋)和2017年2月(冬)、5月(春)、8月(夏)大潮期间, 在獐子岛岩相潮间带(39°01′E, 122°43′N)采集完整的大型海藻植株。
将不易腐烂的海藻直接放入密封袋中, 叶片较薄、藻体容易被损坏的海藻直接放入盛有海水的采集瓶中, 4℃冷藏, 24 h内带回实验室进行种类鉴定和分析。
1.2 样品处理
已经鉴定的大型海藻用海水清洗, 去除泥沙和附生在表面的动植物, 用吸水纸吸干表面的水分, 使用分析天平测其湿质量(FW), 然后将新鲜样品放入1 mol的HCl蒸汽中进行熏蒸, 过夜, 去除碳酸盐。将熏蒸后的样品用蒸馏水洗净, 在烘箱中烘干(60℃)到恒重, 用分析天平测其干质量(DW), 将干样品研磨至细粉末状进行有机碳含量和13C值分析。
含水率(%)=(FW–DW)/FW
1.3 有机碳含量和δ13C值的测定
称取(1.50±0.50)mg海藻细粉末, 用锡纸进行包埋, 将包埋好的样品用元素分析仪(Vario PYRO)进行有机碳含量的测定; 以PDB(Pee Dee Belemnite)为标准, 利用同位素质谱仪(Isoprine 100)测定样品的13C值。
1.4 数据处理
采用SPSS 11.0 进行数据处理及统计分析。
2 结果
2.1 大型海藻的种类及群落结构
獐子岛岩相潮间带共发现大型海藻49种(表1)。其中红藻门24种(占总数的48.98%), 褐藻门17种(占总数的34.69%), 绿藻门8种(占总数的16.33%)。潮间带大型海藻的种类组成以红藻门为主, 其次为褐藻门, 绿藻门最少。

表1 獐子岛岩相潮间带大型海藻物种名录
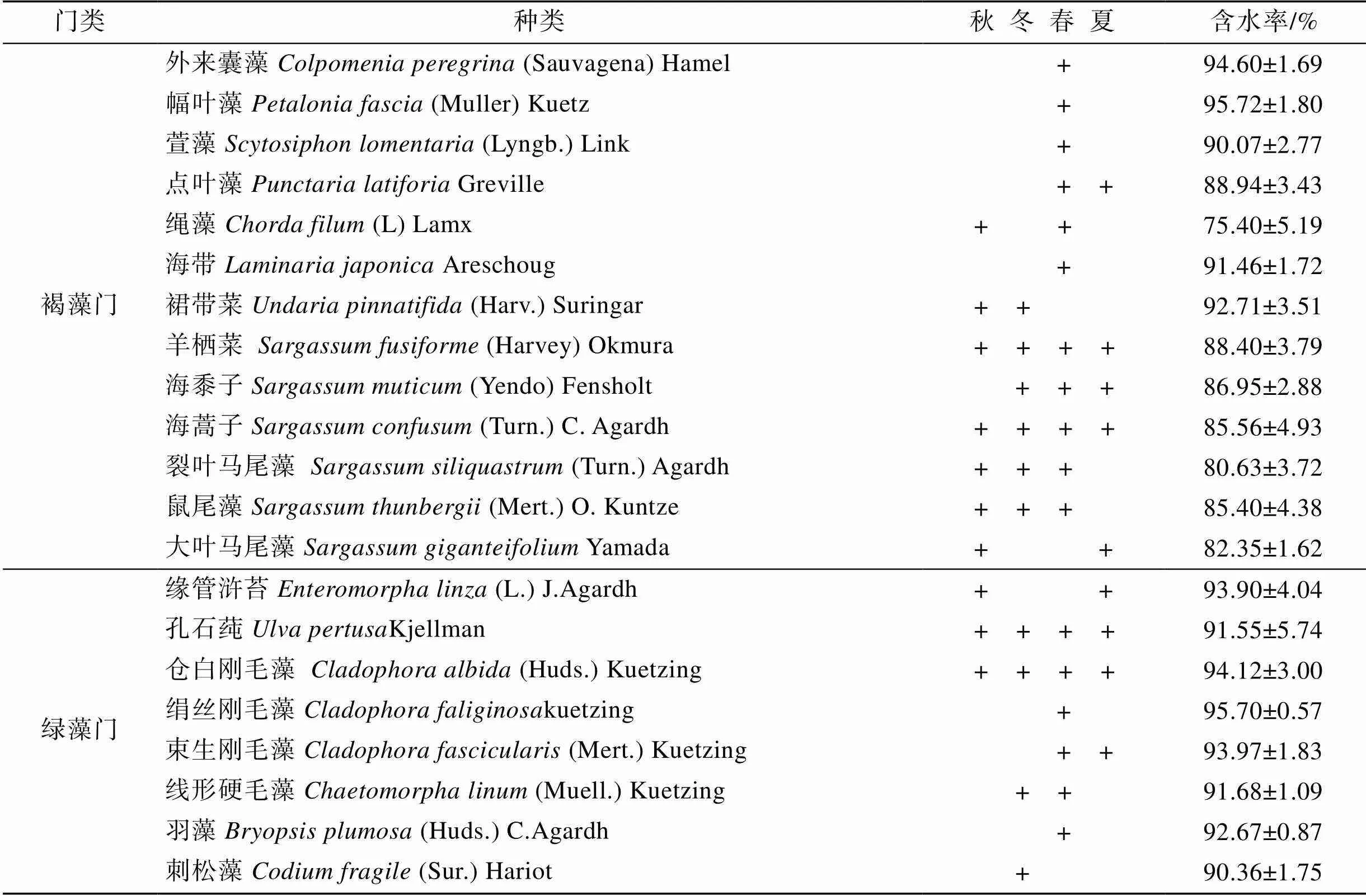
续表
注“+”代表该物种出现
从图1可以看出, 大型海藻种类数在一年四季中的变化, 秋季23种、冬季24种、春季35种、夏季24种。春季海藻种类最多, 其中红藻16种(占总数的45.71%), 褐藻13种(占总数的37.14%), 绿藻6种(占总数的16.22%)。秋季海藻种类数最少, 其中红藻12种(占总数的52.17%), 褐藻8种(占总数的34.78%), 绿藻3种(占总数的13.43%)。无论何季均以红藻门种类数相对最多, 绿藻门相对最少。一年中均能采集到的海藻种类共计12种, 其中红藻门7种(鸡毛菜、小珊瑚藻、扇形拟伊藻、角叉菜、绒线藻、被芯松节藻和鸭毛藻), 褐藻门3种(囊藻、羊栖菜和海蒿子), 绿藻门2种(孔石莼和仓白刚毛藻)。
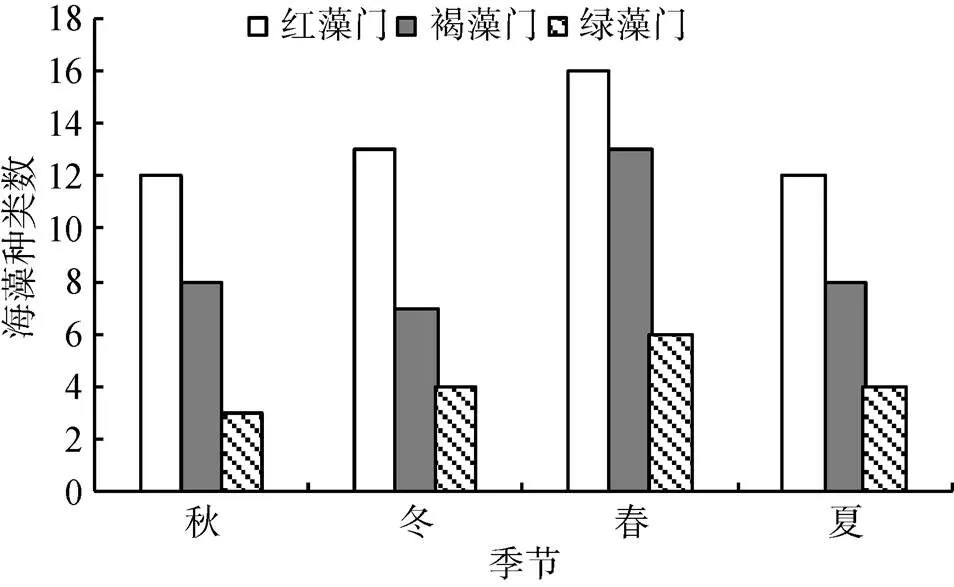
图1 獐子岛岩相潮间带不同季节大型海藻的种类数
2.2 不同门类大型海藻有机碳含量和δ13C值的变化
表2可见, 獐子岛岩相潮间带大型海藻有机碳含量和13C值的变化, 有机碳含量为15.54%~ 35.03%,13C值为–33.42‰~–7.43‰。不同门类大型海藻的体内的有机碳含量有着明显的不同(图2)。红藻门的种类体内有机碳含量多为25%~30%, 以及大于30%; 褐藻门的种类体内有机碳含量多为25%~ 30%; 绿藻门的种类体内有机碳含量在秋季多为25%~30%, 而其他三个季节藻体内有机碳含量在秋季多为大于30%。从图3可以看出不同门类大型海藻的体内的13C值的变化。红藻门的种类体内13C值在夏、秋季节多为–20‰~–10‰, 而在冬季转变为多为–30‰~–20‰; 褐藻门和绿藻门的种类体内13C值多为–20‰~–10‰。
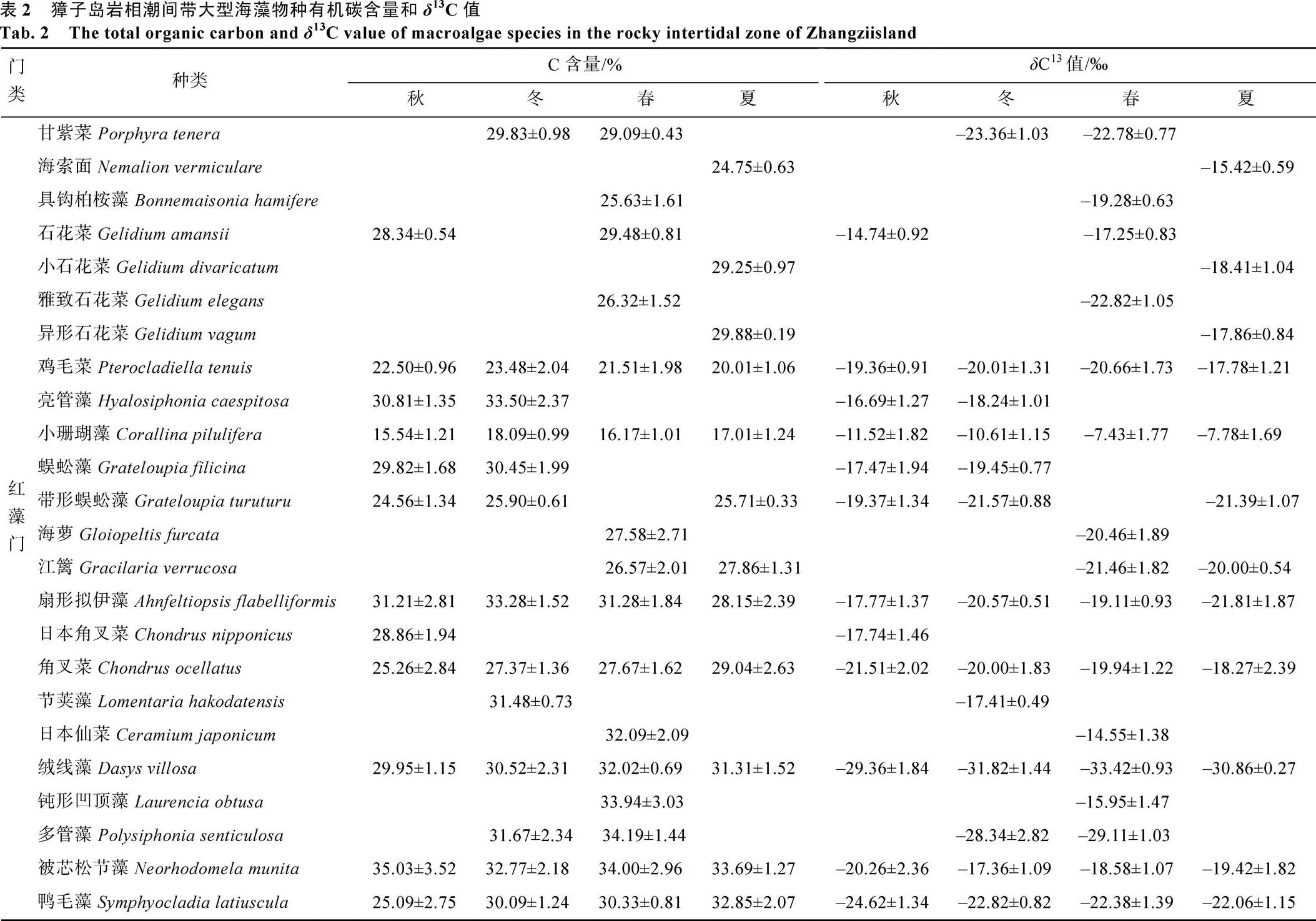
图2 獐子岛岩相潮间带不同门类大型海藻体内有机碳含量统计
Fig. 2 Statistics of the total organic carbon contents of macroalgae in the rocky intertidal zone of Zhangzi island

图3 獐子岛岩相潮间带不同门类大型海藻体内δ13C值统计
2.3 不同季节大型海藻藻体有机碳含量和δ13C值的变化
在獐子岛岩相潮间带地区, 全年均采集到的大型海藻共12种, 以此为材料, 对不同季节海藻体内有机碳含量和13C值变化进行分析。从图4看到, 不同季节大型海藻体内的有机碳含量, 绿藻门最高, 褐藻门最低。海藻体内有机碳含量为冬季>春季>夏季>秋季, 但绿藻门、褐藻门和红藻门海藻体内有机碳含量季节变化并不一致。不同季节大型海藻体内13C值有明显的变化(图5),13C值为夏季>春季>冬季>秋季。红藻门和褐藻门海藻体内13C值在秋冬季节最低, 而绿藻门海藻体内13C值秋季达到最低。

图4 獐子岛岩相潮间带不同季节大型海藻体内有机碳含量变化
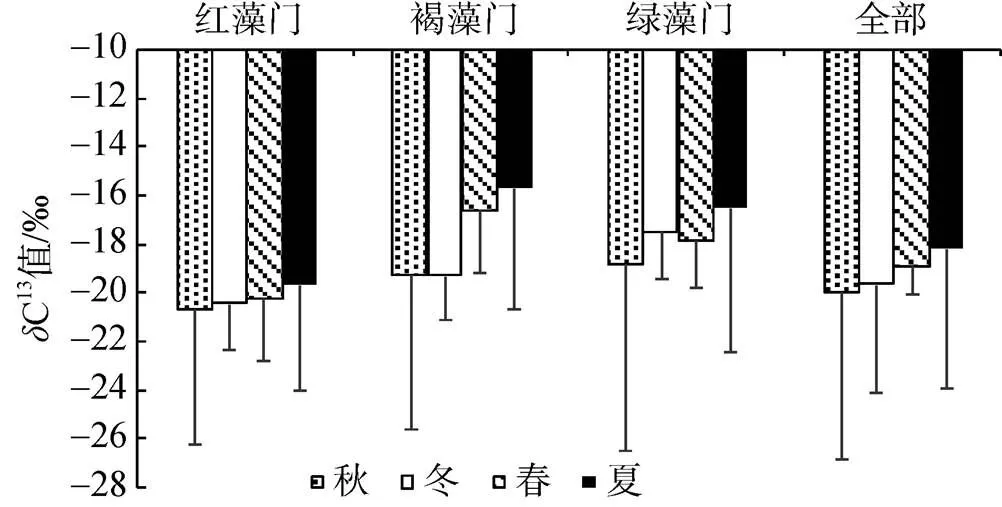
图5 獐子岛岩相潮间带不同季节大型海藻体内δ13C值变化
3 讨论
3.1 大型海藻的群落结构的季节变化
温度作为决定大型海藻分布的重要生态因子, 直接影响海藻的生长、发育和形态特征, 所以会对潮间带大型海藻的群落结构有很大的影响。獐子岛属于温带大陆性季风气候区, 温度在四季有着明显的变化, 因此, 潮间带的大型海藻的种类组成随着季节的变化也出现了明显的改变。为了保护本次调查点的海藻生存环境, 本次调查只进行了定性研究, 共计发现大型海藻49种, 其中12个种类的大型海藻在潮间带终年可见, 它们主要是一些多年生和一年中能繁殖几代的种类, 这些海藻生长能力相对较强, 适应温度等生态环境因子能力强。余下的37种大型海藻除少部分属于偶见种(如异形石花菜)外, 均属于随季节变化的种类。不同种类的大型海藻在不同季节所处的生活史阶段不同, 出现的配子体和孢子体的形态、大小和生活期限等方面也存在较大差异, 这也是造成大型海藻种类季节变化的主要原因之一。一些海藻属于冷水性种类, 在气温较低的冬季和春季比较繁盛, 如: 甘紫菜、多管藻、幅叶藻、羽藻等。而在夏季由于气温较高, 这些冷水性海藻会消失, 而一些暖水性的种类大量出现, 如: 海索面、鞭状索藻、叉开网翼藻等。田丽斯等[15]于2005—2007年的调查结果表明, 獐子岛潮间带大型海藻种类的变化趋势是春季最多, 到了夏季随着水温的上升和持续高温的出现逐渐减少, 秋季达到最小值, 秋后冬初随着气温和水温的降低, 海藻种类又有增加的趋势不过幅度不大, 入冬以后增长幅度逐渐加大。本次调查的结果也呈现出这种趋势, 潮间带大型海藻种类的季节变化为春季35种、冬季24种、夏季24种、秋季23种, 其中春季海藻的种类数量明显高于冬、夏和秋季。另外, 本次调查中还发现, 各门类大型海藻的种类数在不同季节均为红藻门>褐藻门>绿藻门。红藻门种类虽然最多, 但是由于个体相对较小, 生物量并不高; 而褐藻门由于种类个体相对较大, 所以生物量较高。特别是在春季和夏季, 一些褐藻(如海蒿子、海黍子等)会成为优势种, 地毯式的大片覆盖在潮间带岩石的表面, 虽然种类数较为单一, 但是生物量巨大。
3.2 大型海藻体内有机碳含量和δ13C值的变化
海洋作为地球上最大的碳库, 可以大量的吸收由于人类活动而释放出的CO2, 在调节气候变化中起着重要的作用。海洋的固碳机制是现今的研究热点之一, 海洋固碳主要依靠溶解度泵和生物泵两个驱动过程, 这些过程对调节大气中 CO2的浓度和全球碳循环的系统过程起着非常重要的影响[16]。溶解度泵与海洋溶解CO2的能力密切相关, 这一过程会使碳酸盐平衡体系的波动, 导致海洋pH值下降[17]。海洋生物泵是以一系列海洋生物为介质将大气中的碳输运到海洋深层的过程, 是海洋碳循环的重要组成部分以及未来的研究重点[18]。研究表明, 潮间带大型海藻会通过光合作用将无机碳转化为有机碳, 进而进入海洋生物泵的过程, 这些无机碳主要来源于空气中的CO2及水中溶解的CO2和HCO3–[19-20]。周伟男[21]对硇洲岛岩相潮间带50种大型海藻的固碳作用进行分析表明, 各种类海藻有机碳含量的变化范围为 4%~42%, 平均含量为27.57%。本研究表明, 獐子岛岩相潮间带不同藻类体内的有机碳含量在15.54%~35.03%之间变动, 平均含量为27.70%。獐子岛与硇洲岛岩相潮间带大型海藻体内有机碳含量变化范围存在差异, 主要是由于两地大型海藻的种类存在很大的差别, 导致有机碳含量的变化范围不同, 但两地大型海藻体内有机碳的平均含量基本一致。獐子岛岩相潮间带多数的大型海藻(78.30%)体内有机碳含量大于25%, 仅有少量的大型海藻体内有机碳含量较低。在12种终年可见种中, 9种海藻体内的含有机碳量大于25%, 其中扇形拟伊藻、绒线藻、被芯松节藻和仓白刚毛藻含有机碳量均大于30%; 而小珊瑚藻体内有机碳含量最低, 仅为16.70%±1.10%。
大型海藻类除了能够利用CO2作为光合作用外在无机碳源外, 许多种类的大型海藻还能够利用HCO3–作为光合作用外在碳源。HCO3–的利用主要为胞外碳酸酐酶(CA)催化的HCO3–与CO2的相互转化及HCO3–的直接吸收两种方式[19]。研究表明, 大型海藻体内13C值可以指示大型海藻光合作用的碳源利用途径[12-13]。当藻类的13C≤–30‰时, 藻类通常仅依赖于CO2扩散进行光合作用; 相反, 当13C≥–10‰时, 物种光合作用通常利用的是HCO3–;13C值在–30‰和–10‰之间时, 藻类光合即可利用CO2也可以利用HCO3–作为C源[22, 23]。在獐子岛岩相潮间带所采集的大型海藻体内13C值在–7.43‰~ –33.42‰之间。Jesús等[22]在西班牙Cádiz湾调查发现, 大型海藻13C值在–6.08‰~–33‰之间, 这与本次调查结果一致。从结果可以看出小珊瑚藻体内13C值较高, 在春夏两季13C≥–10‰, 表明其利用HCO3–进行光合作用; 而绒线藻内13C值较低, 除秋季外13C值均≤–30‰, 表明其依赖于CO2扩散进行光合作用。本研究还发现, 红藻门13C值(–19.97‰)低于褐藻门(–18.31‰)和绿藻门(–18.26‰), 而且海藻体内13C值会出现季节变化(夏季>春季>冬季>秋季)。Wang和Yeh对台湾澎湖岛大型海藻体内13C值进行分析, 也发现了红藻门13C值低于其他门类的现象[13]。已有研究表明, 大多数红藻只具有很弱的HCO3–利用能力, 其HCO3–利用能力远远不如褐藻和绿藻[19, 22], 这可能是红藻门13C值低于褐藻门和绿藻门的主要原因。Raven等[24]研究也发现不同季节海藻体内13C值的变化, 主要原因是由于在春夏季节温度升高, 特别是处于正午退潮期间的潮池会显著变暖, 导致大型海藻利用的CO2减少, 这些环境条件的变化会促使藻体内13C值升高。
[1] 孙利芹, 王书亚, 孙海林, 等. 山东省大型经济海藻养殖现状及发展对策[J]. 齐鲁渔业, 2017, 34(6): 29- 33. Sun Liqin, Wang Shuya, Sun Hailin, et al. Development countermeasures of large-scale economic seaweed farming in Shandong Province[J]. Shandong Fisheries, 2017, 34(6): 29-33.
[2] 曾宴平, 马家海, 陈斌斌, 等. 浙江省枸杞岛潮间带大型底栖海藻群落的研究[J]. 浙江农业学报, 2013, 25(5): 1096-1102. Zeng Yanping, Ma Jiahai, Chen Binbin, et al. Survey on the community of benthic macro-algae in Gouqi island of Zhejiang Province[J]. Acta AgriculturaeZhejiangensis, 2013, 25(5): 1096-1102.
[3] 黄银爽, 欧林坚, 杨宇峰. 广东南澳岛大型海藻龙须菜与浮游植物对营养盐的竞争利用[J]. 海洋与湖沼, 2017, 48 (4): 806-813. Huang Yinshuang, OuLinjian, Yang Yufeng. Nutrient competition between macroalgaeand phytoplankton in coastal waters of Nan’ao island, Guangdong[J]. Oceanologia et Limnologia Sinica, 2017, 48(4): 806-813.
[4] Glibert P M, Burkholder J A M. Harmful algal blooms and eutrophication: “strategies” for nutrient uptake and growth outside the Redfield comfort zone[J]. Chinese Journal of Oceanology and Limnology, 2011, 29(4): 724-738.
[5] Glibert P M, Wilkerson F, Dugdale R, et al. Pluses and minuses of ammonium and nitrate uptake and assimilation by phytoplankton and implications for productivity and community composition, with emphasis on nitrogen-enriched conditions[J]. Limnology and Oceanography, 2016, 61(1): 165-197.
[6] 宋晓丽, 黄蕊, 苑克磊, 等. 山东半岛东部沿海绿潮灾害的发生特点[J]. 海洋环境科学, 2015, 34(3): 391-395. Song Xiaoli, Huang rui, Yuan Kelei, et al. Characteristics of the green tide disaster of east Shandong Peninsula offshore[J]. Marine Environmental Science, 2015, 34(3): 391-395.
[7] 刘帆, 费鲜芸, 王旻烨, 等. 黄、东海海域浒苔时空分布变化特征研究[J]. 海洋环境科学, 2017, 36(3): 416-421. Liu Fan, Fei Xianyun, Wang Minye, et al. Research on the Characteristics ofin Yellow Sea and East Sea[J]. Marine Environmental Science, 2017, 36(3): 416-421.
[8] Mou S, Zhang Y, Li G, et al. Effects of elevated CO2and nitrogen supply on the growth and photosynthetic physiology of a marine cyanobacterium,PCC7002[J]. Journal of Applied Phycology, 2017, 29(4): 1755-1763.
[9] 何培民, 刘媛媛, 张建伟, 等. 大型海藻碳汇效应研究进展[J]. 中国水产科学, 2015, 22 (3): 588-595. He Peimin, Liu Yuanyuan, Zhang Jianwei, et al. Research progress on the effects of macroalgae on carbon sink[J]. Journal of Fishery Sciences of China, 2015, 22(3): 588-595.
[10] Chung I K, Beardall J, Mehta S, et al. Using marine macroalgae for carbon sequestration: a critical appraisal[J]. Journal of Applied Phycology, 2011, 23(5): 877- 886.
[11] 权伟, 应苗苗, 康华靖, 等. 中国近海海藻养殖及碳汇强度估算[J]. 水产学报, 2014, 38(4): 509-514. Quan Wei, Ying Miaomiao, Kang Huajing, et al. Marine algae culture and the estimation of carbon sink capacity in the coastal areas of China[J]. Journal of Fisheries of China, 2014, 38(4): 509-514.
[12] Andrew P M, Glenn A H, Matheus C C, et al. Physical and biogeochemical correlates of spatio-temporal variation in the13C of marine macroalgae[J]. Estuarine, Coastal and Shelf Science, 2015, 157: 7-18.
[13] Wang W L, Yeh H W.13C values of marine macroalgae from Taiwan[J]. Bot Bull Acad Sin, 2003, 44(2): 107- 112.
[14] 冯天威, 刘正一, 付晚涛, 等. 辽宁省黄海沿岸人工岸线潮间带大型海藻调查与分析研究[J]. 海洋科学, 2013, 37(12): 17-27. Feng Tianwei, Liu Zhengyi, Fu Wantao, et al. Investigation of artificial shoreline seaweed along the Yellow Sea coast in Liaoning Province[J]. Marine Sciences, 2013, 37(12): 17-27.
[15] 田丽斯, 李莹, 张明, 等. 獐子岛潮间带底栖海藻资源及其季节性变化[J]. 水产科学, 2009, 28(3): 142- 145. Tian Lisi, Li Ying, Zhang Ming, et al. The benthic marine alga resources and seasonal changes in intertidal zone at Zhangzi island[J]. Fisheries Science, 2009, 28(3): 142-145.
[16] 李纯厚, 齐占会, 黄洪辉, 等. 海洋碳汇研究进展及南海碳汇渔业发展方向探讨[J]. 南方水产, 2010, 6(6): 81-86. Li Chunhou, Qi Zhanhui, Huang Honghui, et al. Review on marine carbon sink and development of carbon sink fisheries in South China Sea[J]. South China Fisheries Science, 2010, 6(6): 81-86.
[17] Ferreira A F, Ortigueira J, Alves L, et al. Biohydrogen production from microalgal biomass: energy requirement, CO2emissions and scale-up scenarios[J]. Bioresour Technol, 2013, 144: 156-164.
[18] 孙军, 李晓倩, 陈建芳, 等. 海洋生物泵研究进展[J]. 海洋学报, 2016, 38(4): 1-21. Sun Jun, Li Xiaoqian, Chen Jianfang, et al. Progress in oceanic biological pump[J]. Haiyang Xuebao, 2016, 38(4): 1-21.
[19] 邹定辉, 高坤山. 大型海藻类光合无机碳利用研究进展[J]. 海洋通报, 2001, 20(5): 83-90. Zou Dinghui, Gao Kunshan. Progress in Studies on Photosynthetic Inorganic Carbon Utilization in Marine Macroalgae[J]. Marine Science Bulleten, 2001, 20(5): 83-90.
[20] 张继红, 方建光, 唐启升. 中国浅海贝藻养殖对海洋碳循环的贡献[J]. 地球科学进展, 2005, 20(3): 359- 365. Zhang Jihong, Fang Jianguang, Tang Qisheng. The contribution of shellfish and seaweed mariculture in China to the carbon cycle of coastal ecosystem[J]. Advance in Earth Sciences, 2005, 20(3): 359-365.
[21] 周伟男. 硇洲岛岩礁带底栖生物的群落结构及大型海藻的碳汇作用[D]. 湛江: 广东海洋大学, 2013. Zhou Weinan. The macrobenthos community structure and effects on the carbon sink of macroalgae in the intertidal rocky zone in Naozhou island[J]. Zhanjiang: Guangdong Ocean University, 2013.
[22] Jesús M M, Santos C B, Lucas P, et al. Carbon isotopic fractionation in macroalgae from Cádiz Bay (Southern Spain): Comparison with other bio- geographic regions[J]. Estuarine Coastal & Shelf Science, 2009, 85(3): 449- 458.
[23] María M, Antonio D H, José A F, et al. Photosynthetic use of inorganic carbon in deep-water kelps from the Strait of Gibraltar[J]. Photosynth Res, 2016, 127: 295- 305.
[24] Raven J A, Johnston A M, Kubler J E, et al. Seaweeds in cold seas: evolution and carbon acquisition[J]. Annals of Botany, 2002, 90(4): 525-536.
Seasonal variation in the total organic carbon contents and the13C values of macroalgae in the rocky intertidal zone of the Zhangzi island
ZHANG Jing-fan1, CAI Heng-jiang1, 2, ZHAO Yue-ru1, CHENG Wen-han1, HU Si-qi1, LIU Yuan1, 2, LIU Chang-fa1, 2
(1. College of Marine Technology and Environment, Dalian Ocean University, Dalian 116023, China; 2. Key Laboratory of Nearshore Marine Environmental Research, Dalian 116023, China)
To study the seasonal variation in the total organic carbon contents and13C values of macroalgae in the rocky intertidal zone of Zhangzi island (39°01′E, 122°43′N). The macroalgae was investigated in November (Autumn) 2016, February (Winter), May (Spring), and August (Summer) 2017. At the same time, the total organic carbon contents and13C values in the collected macroalgae were analyzed. A total of 49 species from 3 phyla of macroalgae were collected. Of these, 24 species (48.98%) were Rhodophyta, 17 species (34.69%) were Phaeophyta, and 8 species (16.33%) were Chlorophyta. The number of macroalgae species were 35 in spring, 24 in winter, 24 in summer, and 23 in autumn. The total organic carbon content of the different species of macroalgae was 15.54%– –35.03%, and the13C value was −33.42%–−7.43%. The total organic carbon contents of macroalgae in different seasons were winter > spring > summer > autumn, and the13C values were summer > spring > winter > autumn.
macroalgae; species; seasonal variation; total organic carbon;13C value
Aug. 29, 2019
[National Natural Science Foundation of China, No.41306104; Scientific Research Project of Liaoning Provincial Education Department, No.JL201906; The Wetland Program of Pink Beach, No.PHL-XZ- 2017013-002]
Q918.82
A
1000-3096(2020)02-0056-10
10.11759/hykx20190829002
2019-08-29;
2019-12-12
国家自然科学基金项目(41306104); 辽宁省教育厅科学研究项目(JL201906); 红海滩湿地项目(PHL-XZ-2017013-002)
张靖凡(1994-), 女, 辽宁沈阳人, 硕士研究生, 研究方向: 海洋生态学, 电话: 17612495978; E-mail: 497117939@qq.com; 蔡恒江(1978-),通信作者, 副教授, 研究方向: 海洋生态学, E-mail: caihj@dlou.edu.cn
(本文编辑: 康亦兼)
猜你喜欢
百科探秘·海底世界(2024年2期)2024-02-27 14:37:15
法人(2022年5期)2022-05-23 02:30:11
科学与财富(2021年35期)2021-05-10 14:54:31
海洋信息技术与应用(2020年4期)2021-01-18 06:21:36
海洋通报(2020年3期)2020-10-20 09:26:36
党员生活·中(2019年12期)2019-08-13 07:45:54
核农学报(2019年1期)2019-01-09 08:04:44
石油化工建设(2018年1期)2018-07-10 09:49:44
经理人(2018年1期)2018-02-03 17:56:04
科学家(2017年23期)2018-01-11 20:54:58
