
干旱区荒漠植物初级生产力及碳储量与功能性状之间的关系∗
2020-03-06 02:21:54吕光辉蒋腊梅李周康
新疆大学学报(自然科学版)(中英文) 2020年1期
张 磊,吕光辉,蒋腊梅,李周康,张 仰
(1.新疆大学资源与环境科学学院,新疆乌鲁木齐830046;2.新疆大学干旱生态环境研究所,新疆乌鲁木齐830046;3.绿洲生态教育部重点实验室,新疆乌鲁木齐830046)
0 引言
植物功能性状(plant functional traits)近年来已成为生态学和全球变化研究的热点[1,2].它不仅能反映植物在获取、利用和保存资源方面的能力与植物的生长对策、植株生物量及植物利用资源的关系密切相关[3],还能将环境、植物个体和生态系统结构、过程与功能紧密地联系起来[4].同时,通过植物功能性状来指示生态系统功能已成为当前新的研究视角[5].
群落生物量作为生态学研究中一种重要的数量特征,能够衡量植被的动态和生态系统功能的变化[6].生态系统中植被的碳主要来源于植物通过光合作用固定大气中的CO2,有机碳含量反映了植物的光合碳同化能力,是植物碳储量的一种度量[7].同时,植被碳储量的估算也是揭示植被恢复对陆地生态系统影响的核心内容[8].由于野外工作难度大,关于植物生物量及其碳储量的研究多基于结合资料估算等手段[9,10].根据实际测量结果,不同植物碳转换系数不尽相同,且地上部分与地下部分碳转换系数存在差异,这会造成对植物碳储量估算的偏差[11].基于此,本文采用实测方式来探讨影响植物碳储量的因素.在植物碳含量的研究中,不同植物固碳能力有所差异,近年来研究得出的结果也不同[12].本文以不同生活型荒漠植物为研究对象,在不同组织层次上讨论其固碳能力.
植物生物量及其碳储量与生态系统的物质周转和养分循环具有密切的联系,主要受植物生长型[13]、植物构件[14]、生境[15]、演替时间[16]等因素影响.近年来,植物功能性状与生态系统功能关系的研究越来越受到人们的关注.性状作为研究的解释变量是功能生态学研究的主要内容之一,植物性状对生态系统功能的变化有显著的指示作用[17].Mokany[18]认为在生态系统服务中,物种的组成与其所具有的功能性状对生态系统的功能起着重要作用,因此,群落的功能性状决定了生态系统的功能.目前,大部分研究只是从环境土壤角度与植物生产力及固碳能力等方面展开工作[19],有关植物功能性状与初级生产力及植被碳储量方面的研究相对较少且结论各异,有学者[20]从元素角度发现,植物体内的C、P通过对养分的限制和在器官中的分配,从而影响光合作用,与植物碳密度显著相关;对植物表型性状而言,冠幅、叶长、叶片数量、叶面积对地上生物量贡献率达到72.93 %[21];对植物光合能力来说,比叶面积、干物质含量、叶绿素等功能指标与森林生态系统碳储量密切相关[22].基于此,本文从植物功能性状的角度来探讨对植物生产力及固碳水平的影响.
干旱区是最脆弱的生态系统类型之一,水资源极为短缺[23],在全球范围内干旱区分布面积约占陆地面积的1/3以上[24].当前,由于全球变化和不合理的开发,干旱荒漠区的植物在生长繁殖及多样性维持方面遭受到严重威胁.艾比湖流域是典型的干旱荒漠生态系统,现有的研究主要针对荒漠植物的群落特征[25]、植物生理[26]等方面开展工作,而针对干旱区荒漠植物的生产力及碳储量的研究甚少.本文以艾比湖流域荒漠植物为研究对象,研究不同生活型荒漠植物功能性状的差异及其碳储量与群落初级生产力二者之间的关系,以期为干旱区荒漠生态系统植被碳固存潜力对区域生态系统的恢复和碳平衡提供重要理论和实践参考.
1 研究区概况
新疆艾比湖湿地国家级自然保护区地跨新疆博州精河县,范围位于艾比湖湿地作为准噶尔盆地的最低凹地,独特的地理位置使其成为了水盐汇集中心,其水资源主要来源于河流及地下水补给.艾比湖地区干旱少雨,蒸发量大,空气干燥,属典型的温带大陆性气候,日照时数约2 800 h,日平均气温为68 ℃,积温3 000∼3 500 ℃,年平均降水量90.9 mm.区系成分以干旱植物种类为主,主要植被类型除梭梭(Haloxylon ammodendron)、胡杨(Populus euphratica)外,在平原低地还有铃铛刺(Halimodendron halodendron)、新疆绢蒿(Seriphidium kaschgaricum)、白刺(Nitraria tangutorum)、盐爪爪(Kalidium foliatum)、骆驼刺(Alhagi sparsifolia)、小獐毛(Aeluropus pungens)、罗布麻(Apocynum venetum)、对节刺(Horaninowia ulicina)、盐地碱蓬(Suaeda salsa)、芦苇(Phragmites australis)、盐节木(Halocnemum strobilaceum)、沙拐枣(Calligonum mongolicum)、花花柴(Karelinia caspia)等.
2 材料与方法
2.1 样地设置及采样
2016年7月,在艾比湖流域,距阿其克苏河5 km,沿阿其克苏河垂直方向,东西相距100 m设置3条样带,长约1.62 km,每条样带南北间隔40 m,设置1.5 m×1.5 m小样方,小样方共计120个.记录每个样方的地理坐标、海拔和植被类型,调查样方内所有物种的个数,对样方内的每一种植物选择最大、最小、中等形态的个体进行表型测量(株高、冠幅),每一种植物随机选取5个完整的叶片,对叶面积进行拍照后,分类装袋带回实验室,用于后期比叶面积的测定,使用Sigmas4.1计算叶片面积.对出现在样方内的植物,采集20 g左右完整且成熟的叶片,按“同样方同物种”的原则放入自封袋,自然晾干后带回实验室,65 ℃下烘干48 h后粉碎,过筛(100目),用于后期叶片养分的测定.同时,采集样方内全部植物的地上、地下部分,分类装袋标记,自然风干,带回实验室处理.
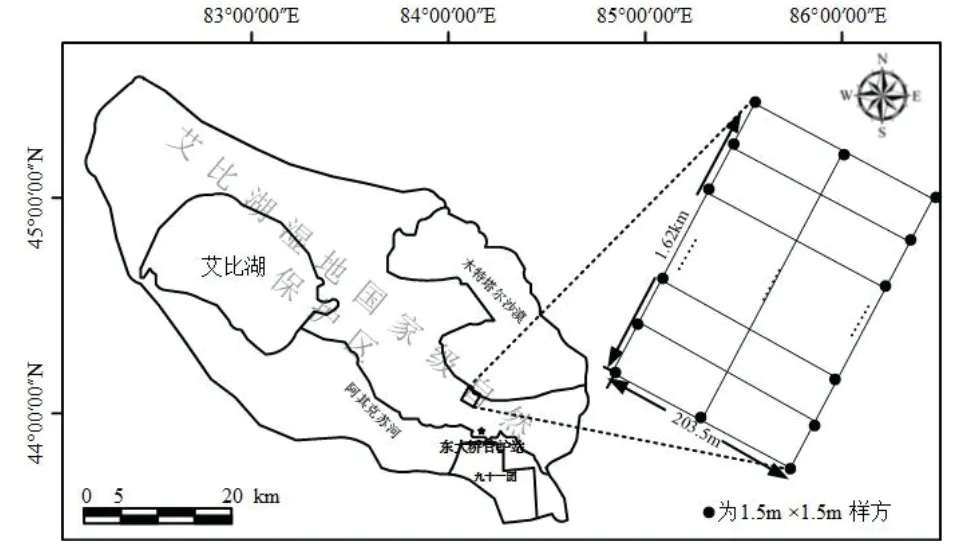
图1 研究区样地布设图Fig 1 Layout of the study area
2.2 实验方法
将分类好带回的植物地上、地下部分用蒸馏水冲洗掉表面所存微量泥土,65 ℃下烘干72 h直至重量恒定,称量其干重.植株地上、地下部分有机碳的测定使用重铬酸钾法.植物功能性状指标的选取、测定及其生态学意义如表1所示.
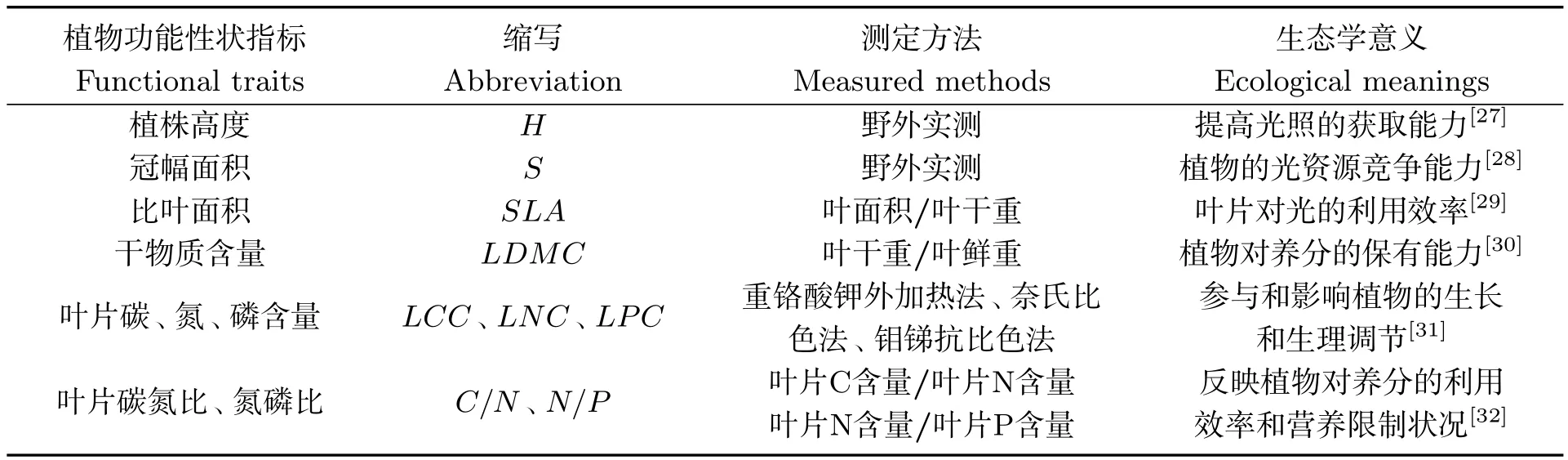
表1 植物功能性状指标的测定方法及生态学意义Tab 1 The measured methods and ecological meanings of plant functional traits
2.3 数据处理
将研究区的植物按照植物生活型划分为一年生草本(AG,annual grass)、多年生草本(PG,perennial grass)和灌木(S,shrub)3种类型;首先对样方数据进行正态分布检验,利用Bartlett检验样本总体方差齐性,通过单因素方差分析(ANOVA)比较不同生活型植物功能性状、生物量及其碳储量的差异性,采用SPSS17.0软件分析数据差异显著性;每个样方中不同植物的功能性状通过性状群落权重均值(CWM,community weighted mean)来表示;每个样方的植物群落碳含量由每种植物的碳含量乘以其占样方生物量的比重后加权获得;植物群落功能性状之间的相关分析在R3.5.0 Library(corrgram)程序包中完成;植物初级生产力及碳储量对植物功能性状的响应趋势通过冗余分析来判断,对植物功能性状进行RDA(redundancy analysis)筛选,同时利用蒙特卡洛置换检验(Monte-Carlo permutation test)定量评价9个植物功能性状对植物初级生产力及碳储存特征变化的独立解释量.
2.3.1 群落植物功能性状的计算
性状群落权重均值(CWM),即就某一性状而言,群落内所有该性状的值与其相对丰度积的总和.性状群落权重均值表示群落内某功能性状的均值,常用来评估群落动态和生态系统特征[33].

式中,Pi是物种i在群落中的重要值(重要值=相对多度+相对盖度+相对频度),traiti是物种i的性状值.
2.3.2 植物碳储量的计算

式中,VCD(vegetation carbon density)为植被碳密度(g·m−2),BECi为植被各组分(地上层、地下层)生物量,VCi为各植被层含碳率.
3 结果分析
3.1 不同生活型植物功能性状的差异性
从表2可以看出,不同植物生活型,其功能性状之间存在显著差异(P<0.05).研究区多年生草本、灌木的株高和冠幅面积均明显大于一年生草本植物;其次,多年生草本具有最大的比叶面积(71.65 ± 2.59 cm2·g−1),相比一年生草本和灌木植物,分别高于36.3 %和38.4 %;植物叶片干物质含量、叶片碳含量均表现为:多年生草本>灌木> 一年生草本;灌木植物叶片氮含量均高于一年生草本植物及多年生草本植物,分别高于34.0 % 和20.0 %;对于叶片磷含量指标,多年生草本植物显著高于灌木,高出了9.5 %;不同生活型植物叶片碳氮比均没有表现出显著差异(P <0.05);一年生及多年生草本的叶片氮磷比均显著低于灌木,分别低于29.9%和24.1%.
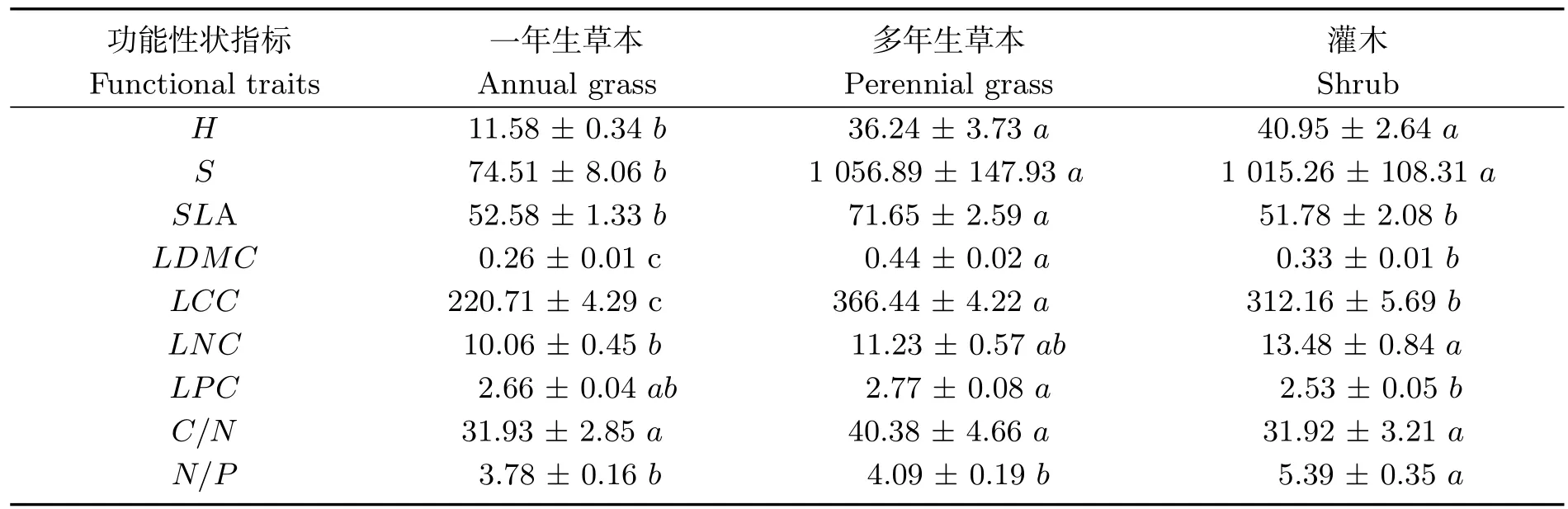
表2 不同生活型植物功能性状的差异性分析Tab 2 Difference analysis of functional traits of different life forms of plants
3.2 不同生活型植物生物量及碳储量的差异性
研究区不同生活型植物在不同组织的生物量及其碳储量(图2)具有较大差异(P <0.05).对于植物地上组织而言,多年生草本具有最大的生物量,相比一年生草本和灌木植物,分别高于76.12 %和46.56 %;植物地上部分碳储量为多年生草本(9.08 ± 1.55 g·m−2)>灌木(5.66 ± 0.81 g·m−2)> 一年生草本(3.16 ± 0.28 g·m−2);对于植物根系的生物量及其碳储量,均呈现出:灌木> 多年生草本> 一年生草本的规律;多年生草本、灌木植物总生物量均显著大于一年生草本植物,分别高于120.8 %和115.5 %;灌木植物具有最大的植株总碳储量,分别是一年生草本和多年生草本植物的4.54倍和1.16倍.根据不同生活型植物地上、地下碳储量的比例(图3)可知,不同生活型植物地下碳储量比重依次为:灌木(72.2 %)> 多年生草本(48.5 %)> 一年生草本(29.6 %),由此可知,不同生活型植物组织碳储量比重存在明显差异.
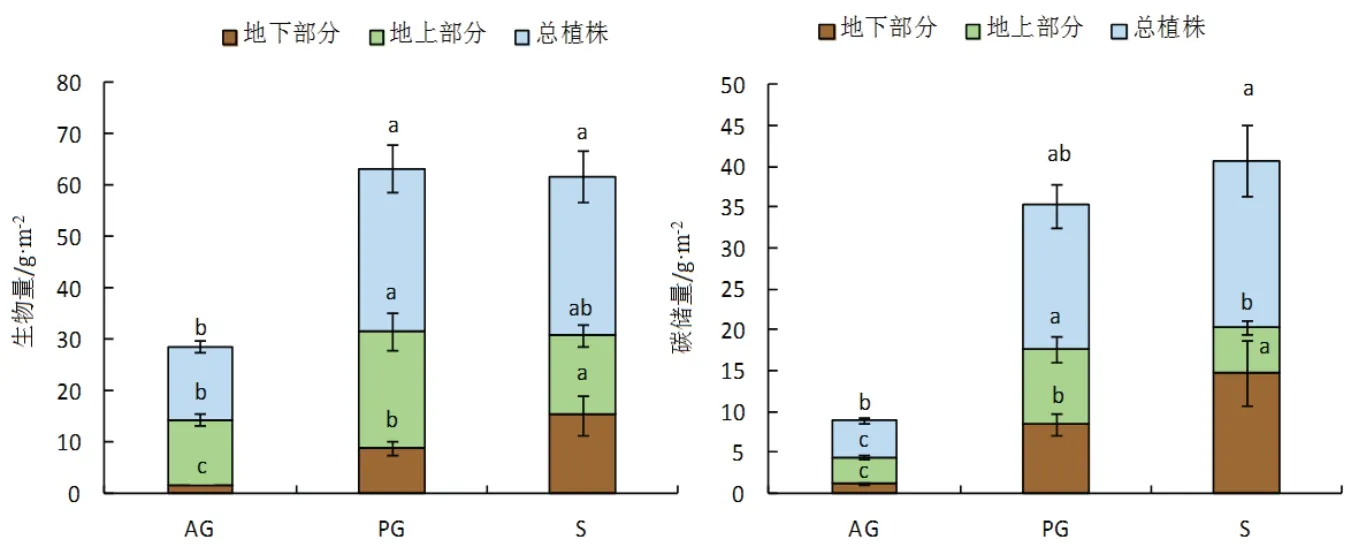
图2 不同生活型植物生物量及碳储量的差异性分析Fig 2 Difference analysis of plant biomass and carbon storage in different life types
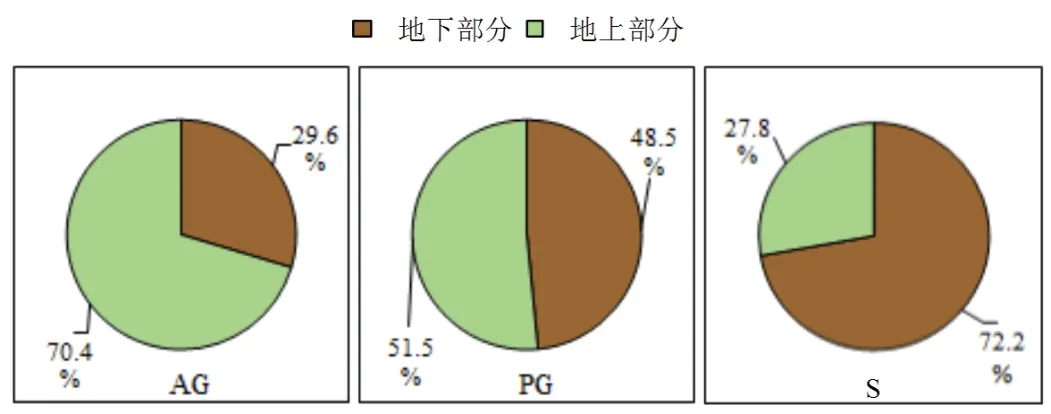
图3 不同生活型植物地上、地下碳储量的比例图Fig 3 Proportion map of above-ground and underground carbon storage of different living plants
3.3 群落植物功能性状的相关性
植物功能性状之间的相关性分析见图4.比叶面积、植株高度、冠幅面积、叶片干物质含量、叶片碳含量在群落水平上表现出极显著的两两正相关关系(P <0.01),其中叶片干物质含量和叶片碳含量、植株高度和冠幅面积的相关程度最高,分别为0.85和0.80;叶片氮含量与株高和冠幅面积显著正相关(P <0.05);叶片磷含量与冠幅面积、叶片干物质含量、叶片碳含量呈显著正相关(P <0.05),与叶片氮含量呈极显著正相关(P <0.01);叶片碳氮比、氮磷比分别与比叶面积、植株高度呈正相关趋势(P <0.05).

图4 植物功能性状间的相关性分析Fig 4 Correlation analysis of plant functional traits
3.4 植物功能性状与初级生产力及其碳储量的关系
对研究区荒漠植物初级生产力及其碳储量和9个植物功能性状进行RDA分析,得到二维排序图(图5).结果如图5a、5b所示:9个植物功能性状因子对荒漠植物初级生产力及其碳储量的解释量分别是31.7 %和22.3 %.植株高度、冠幅面积、叶片碳含量、叶片碳氮比、叶片氮磷比箭头连线相对较长,可知这5个植物功能性状指标是影响荒漠植物初级生产力及其碳储量的关键因素,且叶片碳氮比作为主要抑制因子作用于荒漠植物生长及固碳能力;比叶面积对荒漠植物初级生产力及碳储量呈现出负相关趋势;就植物组织器官而言,叶片氮磷比、叶片氮含量与地下根系的生物量及碳储量相关性更强,植株高度和冠幅面积则与植株地上部分趋势更密切;由图5b可知,叶片碳含量、叶片干物质含量与植株地上部分碳储量呈现极显著相关性.
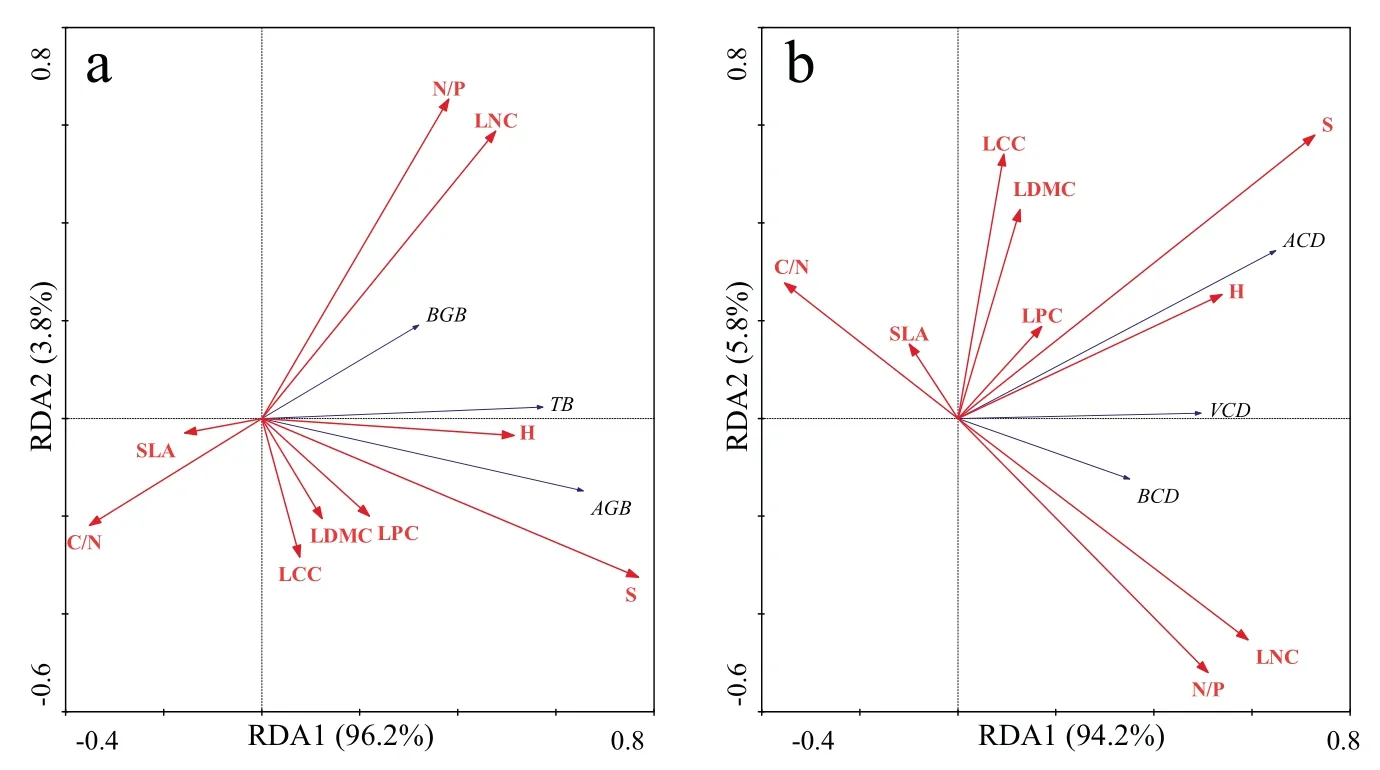
图5 植物功能性状与植物初级生产力和碳储量的二维排序图Fig 5 Two dimensional RDA ordination diagram of plant primary productivity and carbon storage with plant functional traits

表3 荒漠植物初级生产力的功能性状解释重要性排序和显著性检验结果Tab 3 The importance ranking and significance test results explained by functional traits of desert plants primary productivity
对9个植物功能性状和荒漠植物初级生产力及其碳储量进行蒙特卡洛检验,结果如表3、表4所示:影响荒漠植物初级生产力的植物地上功能性状因子重要性排序由大到小依次为:冠幅面积(39.9 %)>植株高度(17.8%)>叶片氮含量(16.2 %)>叶片氮磷比(11.0 %)> 叶片碳氮比(8.3 %);影响荒漠植物碳储量的植物地上功能性状因子排序由大到小依次为:冠幅面积(32.2 %)>叶片氮含量(21.1 %)>植株高度(17.2 %)>叶片氮磷比(16.1 %)>叶片碳氮比(7.5 %);其中冠幅面积、植株高度、叶片氮含量、叶片氮磷比对荒漠植物初级生产力及其碳储量影响极显著(P <0.01),叶片碳氮比表现出显著负驱动力(P <0.05).

表4 荒漠植物碳储量的功能性状解释重要性排序和显著性检验结果Tab 4 The importance ordering and significance test results explained by functional traits of desert plants carbon storage interpretation
4 讨论
4.1 不同生活型植物功能性状的差异
植物通过改变高度、叶面积等功能性状来适应外界环境条件的变化[34].研究区多年生草本和灌木植物的株高(H)和冠幅面积(S)无显著差异且远大于一年生草本,作为多年生植物而言,对自身株高的投入,来增强对光的捕获效率,同时能吸引授粉,使得地上生物量分配增加,固碳能力也随之加强,进而延长植物生活史及提高繁殖能力[35].而一年生草本在干旱的条件下,选择株高变得更为矮小,从而缩短了根系水分到达叶片的距离,提高了水分运输效率[36],顺利完成相对较短的生活史.多年生草本植物的比叶面积(SLA)显著高于一年生草本和灌木,这可能是由于植物在垂直空间分布的差异,使得不同生长型的植物捕获光资源的能力不同造成的.Daz等学者[37]认为高SLA植物拥有较高的光合作用和生长速率,而低SLA植物能更好地适应水分、养分缺乏的环境.研究区多年生草本以构件大的花花柴、芦苇、绢蒿等为主,相比低矮灌木植物(罗布麻、骆驼刺),其光合竞争能力更强,可迅速地获取外部资源,而生长较慢的一年生草本,则具有较低的SLA值,其倾向于更好地保存内部资源[38].研究区植物叶片碳含量(LCC)、干物质含量(LDMC)均表明:多年生草本> 灌木>一年生草本;陈婵等[39]对植物叶片C含量研究发现,叶片C的吸收主要是通过植物光合作用,进而使植物体内的C含量保持在稳定水平.多年生草本具有更大的SLA(71.65 ± 2.59 cm2·g−1),光合作用能力更强于灌木植物,同时综合研究区多年生草本和灌木植物的表型性状(H、S)更优于一年生草本,具有更强的光捕获能力.一年生草本植物的生长主要与环境条件密切相关,而多年生草本比一年生草本有更强的抗逆境和保持种群稳定的能力[40].
4.2 不同生活型植物生物量及碳储量的差异
由于植物光合产物的生产和累积能力不同,干旱区荒漠植物群落组成中,不同生活型植物生物量及碳储量的变化存在着显著的差异.多年生草本植物的地上部分生物量高于一年生草本和灌木.本文中一年生草本、多年生草本、灌木的地上部分所占群落的比例依次是0.25、0.45和0.30;这表明多年生草本植物对干旱缺水的荒漠生态系统具有较强的适应性,且植被地上生产力较大,因此,多年生草本的地上部分碳储量较高.另外,李善家对荒漠植物叶片功能性状的研究发现,SLA大的植物具有较高的生产力,而生物量的转化又与植物碳的同化密切相关[41],本文支持这一结论.灌木地下部分的生物量及碳储量显著高于一年生、多年生草本植物,说明在干旱地区,灌木植物通过增加地下生物量的策略来获取更多水分和养分以供生长所需,研究结果与王鑫[42]等对新疆干旱区常见灌丛生物量碳特征的结论基本一致.综合植株总碳储量来看,相比草本植物来说,灌木根系对植物碳固存具有重要的贡献作用,根系是土壤有机质最主要的来源,直接决定了生态系统碳循环中碳素的周转速率[43].
从植物地上、地下碳储量比例结果发现,一年生、多年生草本地上部分固碳量均大于植物根系,地上部分分别占总碳储量的70.4 %和51.5 %,可知,草本植被碳储量主要分布在地上部分.这与杨阳等学者发现森林生态系统中草本植物地上部分含碳量大于地下部分结果相同[44].灌木植物地下碳储量为14.72 ± 3.94 g·m−2,占总碳储量的72.2 %,这反映了灌木植物向地下部分分配了较多的生物量,以致灌木根系相比草本更为发达,水分是限制灌木生长的关键因子[45],发达的根系有助于该类植被从更深的区域获取水分,这是对干旱胁迫的一种适应[46].
4.3 群落植物功能性状的相关性
群落内植物功能性状之间的相关性是普遍存在的,性状组合的权衡是植物群落构建的重要机制之一,进而影响着生态系统的功能与过程[47].量化植物功能性状之间的关系,特别是多个物种性状间的关系,提供了不同物种间主要变异程度的信息,显示了对生态系统功能的作用能力[48].本文结果表明,植株高度(H)、冠幅面积(S)、比叶面积(SLA)、干物质含量(LDMC)、叶片碳含量(LCC)以及叶片氮含量(LNC)、叶片磷含量(LPC)等性状之间存在极显著正相关关系,这反映了荒漠植物对养分的吸收能力及对光的捕获能力相应的性状具有趋同的综合适应特征[49].
4.4 植物功能性状与初级生产力及其碳储量的关系
植物的功能性状能够为植物生长和适应环境的能力提供重要信息[50].本文以干旱区荒漠植物为研究对象,分析了植物功能性状对群落生产力及其碳储量的影响,结果显示,植株的表型性状即株高(H)和冠幅面积(S)对群落生产力的贡献率超过了50 %,从生态系统的角度出发,个体的矮小化将直接导致植被覆盖度降低,生产力下降.这与李江文等研究发现荒漠草原优势种的株高、冠幅对植物地上生物量的贡献大结果一致[51].而有学者[52]对森林固碳能力与群落功能性状之间的相关分析结果表明,冠幅面积与其呈显著负相关,这可能是森林群落郁闭度较大,植株间枝叶遮阴程度大,植物对光、养分等竞争强度加剧[53],在这种情况下,植物的呼吸消耗并不减弱,从而使净光合速率变低,群落固碳能力变低.而本文研究区群落覆盖度低,植物对光的竞争不强,能很好地适应并快速利用充足的光照,从而使得光合速率变大.氮元素作为构建植物光合器官的重要营养元素,决定着植物生长和植被生态系统的发育.光合器官的氮含量对植物适应干旱环境具有重要的作用[54].已有研究表明[55]植物叶片氮含量对植物的生产力和适应干旱环境的能力具有重要影响.植物通过增加叶片氮含量,提高整体光合能力,从而促进个体生长[56].碳氮代谢是植物生长发育过程中最基本的代谢途径,两大代谢共同调节植物的生长发育进程,影响着植物的光合产量[57].任胜茂等[58]研究发现,叶片光合速率与叶片碳氮比呈极显著负相关,本研究区位于准噶尔盆地,日平均日照时数为6.9 h[59],较强的光照条件下植株具有较高的碳氮同化能力[60],故植物光合速率呈相反趋势,从而抑制了植物光合产物的积累,影响植物生产力及碳储量.叶片的氮、磷含量反映了植物对土壤养分的吸收特征.Güsewell[61]研究认为,氮磷比大于20和小于10作为植物氮磷限制的评价指标,即氮磷比大于20说明植物生长受磷素限制,小于10说明植物生长受氮素限制.本研究区植物叶片氮磷比在3.78∼5.39之间,说明植物受氮素限制,叶片氮磷比的增加对植物生长及固碳能力起到促进作用.
综合上述研究结果,干旱区荒漠生态系统中,植株高度(H)、冠幅面积(S)、叶片氮含量(LNC)、叶片碳氮比(C/N)、叶片氮磷比(N/P)对群落初级生产力及固碳能力具有良好的指示作用.
5 结论
(1)艾比湖流域不同生活型植物功能性状存在差异.多年生草本的比叶面积、叶片干物质含量、叶片碳含量均大于一年生草本和灌木;灌木的叶片氮磷比显著大于草本,且具有最小的叶片磷含量;草本和灌木在叶片碳氮比上没有显著差异.
(2)不同生活型荒漠植物地上固碳量依次为:多年生草本(9.08± 1.55 g·m−2)> 灌木(5.66± 0.81 g·m−2)>一年生草本(3.16 ± 0.28 g·m−2);地下固碳量依次为:灌木(14.72 ± 3.94 g·m−2)> 多年生草本(8.54 ± 1.33 g·m−2)> 一年生草本(1.33 ± 0.10 g·m−2).其中,根系所占植物总固碳比重依次是:灌木(72.2 %)> 多年生草本(48.5 %)> 一年生草本(29.6 %).
(3)荒漠植物对养分的吸收能力及对光的捕获能力相应的性状具有趋同的综合适应特征.叶片碳氮比、氮磷比分别与比叶面积、植株高度显著相关.
(4)冠幅面积、植株高度、叶片氮含量、叶片氮磷比对荒漠植物初级生产力及碳储量具有显著的正效应,叶片碳氮比对荒漠植物初级生产力及碳储量具有负驱动力.
猜你喜欢
中华建设(2024年3期)2024-03-16 08:04:52
中华建设(2023年10期)2023-10-07 05:23:04
小哥白尼(趣味科学)(2022年5期)2022-08-15 08:34:46
中国石化(2021年8期)2021-11-05 07:00:14
少儿美术(2020年8期)2020-11-06 05:21:38
绿色中国(2019年14期)2019-11-26 07:11:34
乡村科技(2018年8期)2018-02-13 12:16:18
诗潮(2017年2期)2017-03-16 10:42:30
湖南林业科技(2017年1期)2017-02-06 05:29:17
文史春秋(2016年3期)2016-12-01 05:42:20
