
花后弱光对江汉平原稻茬小麦的产量及碳、氮分配效应的影响
2020-03-05 03:55:44李刘龙库旭灿王小燕
麦类作物学报 2020年11期
李刘龙,库旭灿,李 赟,王小燕
(长江大学农学院/主要粮食作物产业化湖北省协同创新中心,湖北荆州 434025)
江汉平原为湖北省冬小麦主产区之一,是长江中下游平原重要的组成部分。历史气象数据表明,在江汉平原小麦灌浆期(4-5月),阴雨寡照天气频发,2019年度更有持续超过20 d的极端连阴雨天气出现,长时间的阴雨天气限制了小麦籽粒产量的提高[1-2]。长江流域冬小麦种植面积占全国总面积的16.4%,产量占全国总产量的25%[3-4],因而开展小麦对弱光的响应机制研究及对弱光适应性强的品种进行筛选,对长江流域小麦生产具有重要意义[5]。前人研究表明,弱光会降低小麦的干物质生产[6-7],随着遮光程度的加重,小麦干物质生产量下降幅度加大[8],但不同器官的干物质积累量对弱光的响应因品种而异[9],而且开花前贮藏在营养器官中的非结构性碳水化合物对粒重的贡献率变大[10]。小麦对光能的利用和对氮素的吸收与光照强度紧密相关[11]。已有研究报道,灌浆期各阶段弱光均不利于小麦籽粒中蛋白质的积累[12-13],花后吸收氮素对籽粒氮素的贡献率随遮光程度的加大而增大[14]。也有研究认为,弱光导致植株对氮素吸收强度及积累量减少,分配到叶片和茎鞘的氮素增加[15],分配到穗部的氮素减少[8]。弱光对小麦产量的影响结果不一致,可能因遮光强度和试验材料不同而存在差异。有研究发现,遮光降低了小麦产量,而且产量下降幅度随遮光强度增大而增大[16-17]。遮光后小麦千粒重下降和败育小穗数增多,导致籽粒产量下降[18],但也有人认为,遮光对小麦籽粒产量有促进作用[19],千粒重是籽粒产量发生变化的主要原因[17]。以往研究多集中在遮光对小麦产量[16-19]、干物质[6-8]、光合特性[8,16,19]等方面影响的分析,但是供试材料较为单一,关于江汉平原耐荫品种筛选以及遮光对不同抗荫性小麦品种干物质和氮素的分配与转运的研究未见报道。本研究于2017-2018年度对长江中下游流域适宜推广的48个小麦品种开展灌浆期全程遮光试验,初步筛选出两个弱光敏感品种(扶麦1228和生选6号)和两个弱光钝感品种(襄麦55和扬麦158),2018-2019年以此4个品种和江汉平原主推品种郑麦9023为供试材料,分析开花期至成熟期遮光对小麦干物质积累与分配、植株氮素转运及籽粒产量的影响,以期为江汉平原小麦稳产、高产栽培提供参考。
1 材料与方法
1.1 试验设计
试验于2017-2019年度在湖北省荆州市长江大学农高区科技产业园(30°36′N,112°08′E)进行。土壤为黄黏土,pH值为8.2,有机质含量为14.79 g·kg-1,碱解氮含量为42.79 mg·kg-1,速效磷含量为7.26 mg·kg-1,速效钾含量为75.7 mg·kg-1。两年度试验地降水、日照时数和气温情况见图1。

图1 2017-2018、2018-2019年度小麦生长季气候特征
2017-2018年度对长江中下游流域适宜推广的48个小麦品种开展了灌浆期全程遮光试验,初步筛选出两个弱光敏感品种(扶麦1228和生选6号)和两个弱光钝感品种(襄麦55和扬麦158)。2018-2019年以此4个品种和江汉平原主推品种郑麦9023为供试材料,进一步开展试验。
试验采用裂区设计,遮光(AS)和自然光照(CK)为主区,品种为副区,每个处理重复3次。小区面积12 m2(2 m×6 m),行距25 cm。遮光处理从开花期开始直至成熟期,选用黑色聚乙烯遮光网遮去冠层自然光强的45%(表1)。遮光网距离地面1.8 m,遮光面积超过小区四周,以保证遮光小区完全被覆盖。播前各处理底施纯N 90kg·hm-2、P2O5105 kg·hm-2和K2O 105 kg·hm-2,拔节期追施纯N 90 kg·hm-2。氮肥为尿素(含N 46%),磷肥为过磷酸钙(含P2O512%),钾肥为硫酸钾(含 K2O 60%)。基本苗为225万株·hm-2,其他管理同一般大田栽培。试验地前茬作物为水稻,小麦播种时间为2018年10月31日,襄麦55、扬麦158、郑麦9023、扶麦1228和生选6号分别于2019年4月12日、12日、14日、14日和15日开花,分别于5月19日、19日、21日、21日、21日成熟。

表1 CK和遮光处理的照度值
1.2 研究项目与测定方法
1.2.1 干物质积累量的测定
于开花期、灌浆中期、成熟期每小区分别取10株小麦,按穗、旗叶、其余叶和茎秆分样并分别保存,105 ℃杀青30 min,60 ℃烘干至恒重,用于干物质积累量的测定,同时计算干物质转运特征参数[17,20]。另外自开花期始至收获,每小区每隔7 d取30个同一天开花、长势均匀的麦穗,烘干脱粒后测定粒重,并计算出各阶段的灌浆速率。
花前贮藏干物质转运量=开花期干物质积累量-成熟期干物质积累量;花前贮藏干物质转运效率=花前贮藏干物质转运量/各营养器官开花期干物质积累量×100%;花前贮藏干物质转运量对籽粒产量的贡献率=花前贮藏干物质转运量/成熟期籽粒干物质积累量×100%;花后干物质积累量=成熟期籽粒干物质积累量-花前贮藏干物质转运量;花后干物质积累量对籽粒产量的贡献率=花后干物质积累量/成熟期籽粒干物质积累×100%。
1.2.2 植株器官含氮量的测定
用凯式定氮仪测定各样品全氮含量,同时计算氮素积累与转运特征参数[9,21]。花前贮藏氮素转运量=开花期氮素积累量-成熟期氮素积累量;花前贮藏氮素转运效率=花前贮藏氮素转运量/各营养器官开花期氮素积累量×100%;花前贮藏氮素转运量对籽粒氮的贡献率=花前贮藏氮素转运量/成熟期籽粒氮素积累量×100%;花后氮素积累量=成熟期籽粒氮素积累量-花前贮藏氮素转运量;花后氮素积累量对籽粒氮的贡献率=花后氮素积累量/成熟期籽粒氮素积累×100%。
1.2.3 产量及其构成要素的测定
小麦成熟时每小区选有代表性的1 m2调查穗数,并随机选取30穗考察穗粒数。每小区收获2 m2,脱粒、晒干并计产,之后测定千粒重。
1.3 数据处理
用Microsoft Excel 2016软件对数据进行处理和绘图,用DPS 7.5和SAS 9.2统计分析软 件对数据进行差异显著性检验(LSD法,α= 0.05)。
2 结果与分析
2.1 花后弱光对不同小麦品种籽粒产量的影响
灌浆期遮光显著降低了各品种籽粒产量(表2)。2017-2018年度各品种遮光后籽粒产量范围为1 300~1 900 kg·hm-2,平均值仅为 1 685.6 kg·hm-2,与CK相比平均降低 67.0%;2018-2019年度减产趋势与2017-2018一致,5个品种籽粒产量降低幅度为32.0%~ 48.2%。其中襄麦55和扬麦158的减产幅度相对较小,扶麦1228和生选6号的减产幅度较大,郑麦9023介于两类品种中间。2017-2018年度产量降低幅度大于2018-2019年度。

表2 遮光对小麦产量及产量构成要素的影响
遮光对小麦的千粒重产生显著负效应,遮光处理较CK下降24.9%~38.9%。遮光对穗数和穗粒数影响均不明显。这说明千粒重降低是灌浆期遮光产量下降的主要原因。收获指数与籽粒产量变化趋势一致,遮光处理的收获指数显著低于CK。
2.2 花后弱光对小麦籽粒灌浆动态的影响
灌浆期弱光均导致各品种灌浆早期(花后 0~7 d)、中早期(花后7~14 d)、中期(花后14~21 d)、中后期(花后21~28 d)、后期(花后28~ 35 d)粒重的增长速率减小,粒重增长最快时期由中期后移到中后期(图2)。其中,扶麦1228和生选6号粒重增长受影响较大,襄麦55和扬麦158受影响较小,郑麦9023居中,这与产量对弱光的响应趋势一致。
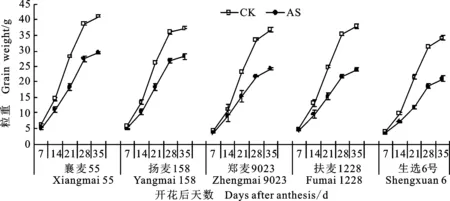
图2 遮光对花后小麦籽粒灌浆动态的影响
2.3 花后弱光对小麦干物质积累及分配的影响
由图3可知,遮光后,5个小麦品种灌浆中期和成熟期干物质积累量均显著降低,并且成熟期干物质下降幅度大于灌浆中期,其中成熟期干物质变化趋势与籽粒产量一致。品种间比较,扶麦1228和生选6号成熟期干物质积累量降低幅度较大,而襄麦55和扬麦158的降低幅度较小,郑麦9023居中。

相同品种和时期图柱上不同字母表示处理间差异显著(P<0.05)。
遮光导致茎鞘、叶片、颖壳、籽粒等器官干物质积累量均较CK均显著降低,分配比例亦发生显著变化(表3),其中籽粒的干物质积累量降低幅度最大,5个品种平均降低了35.6%,其次为叶片和茎鞘,颖壳降低幅度最小。各器官干物质积累量降低趋势也与籽粒产量一致。其中弱光敏感品种生选6号的叶片和茎鞘干物质积累量降低幅度远大于其他品种,其叶片降低幅度高达36.3%(襄麦55和扬麦158仅为10.5%左右),其茎鞘降低幅度亦高达18.3%(襄麦55和扬麦158仅为11%左右)。CK条件下营养器官(叶+茎鞘+颖壳等)干物质总分配比例为53.1%~58.0%,籽粒为42.0%~46.9%;遮光后营养器官总分配比例增加到60.8%~66.2%,籽粒仅为33.8%~39.2%。这表明灌浆期遮光不利于植株中干物质向籽粒中分配。

表3 遮光对不同品种小麦成熟期不同器官干物质积累和分配的影响
2.4 花后弱光对小麦干物质转运及其对籽粒产量贡献率的影响
由表4可知,灌浆期遮光后小麦叶片、颖壳+穗轴、茎鞘等器官花前积累的干物质在花后向籽粒的转运量较CK均显著上升,平均增幅 28.1%,其中穗轴与颖壳平均增加78.3%,茎鞘平均增加87.8%;另外遮光处理的花后光合物质生产量显著下降,且其下降幅度远大于叶片、茎鞘、颖壳等器官干物质转运量的增加幅度,表明花后弱光虽然促进了茎鞘、叶片、颖壳等器官暂存库碳水化物的再分配,但不能弥补其对花后光合生产所造成的损失。

表4 遮光对小麦干物质转运及籽粒产量贡献率的影响
品种间比较,弱光敏感品种营养器官花前干物质的转运量上升幅度远大于弱光钝感品种,遮光后扶麦1228和生选6号的总转运量平均增加70.0%,而襄麦55和扬麦158平均增加42.7%。对于花后积累的干物质量,遮光后扶麦1228和生选6号平均下降85.0%,而襄麦55和扬麦158平均下降62.4%,表明遮光对弱光敏感品种花后光合物质生产能力影响较大。
灌浆期遮光后小麦花前积累的碳水化合物和花后的积累碳水化合物对籽粒产量的贡献率发生不同变化,前者上升,后者下降,且弱光敏感型品种扶麦1228和生选6号两者变化幅度显著大于弱光钝感品种。这说明弱光条件下,小麦籽粒的充实对花前积累的碳水化合物的再分配依赖加大,而花后碳水化合物对籽粒产量的贡献相对 弱化。
2.5 花后弱光对不同小麦品种成熟期各器官氮素积累量的影响
灌浆期遮光显著影响了成熟期叶片、茎鞘、颖壳、籽粒等器官的氮素分配量(即成熟期残留量)(表5)。遮光后各品种营养器官中的氮素分配量即残留量较CK均上升,籽粒中氮素分配量均下降,说明灌浆期遮光抑制了营养器官中的氮素向籽粒的再分配,导致更多的氮素滞留在营养器官中。其中,茎鞘中残留量最大,5个品种平均增加了88.0%,穗轴与颖壳次之,叶片增加幅度最小。

表5 遮光对不同品种小麦成熟期不同器官氮素积累和分配的影响
品种间比较,弱光敏感型品种扶麦1228和生选6号的籽粒氮素积累量的下降幅度大于弱光钝感品种襄麦55和扬麦158,营养器官氮素总残留量的增加幅度亦较大。总之,弱光敏感型品种遮光后成熟期总氮素积累量略增,但营养器官残留氮素量大幅增加,籽粒氮素积累量则显著降低;弱光钝感型品种遮光后籽粒氮素积累量下降幅度和营养器官氮素残留量增加幅度均相对较小。
2.6 花后弱光对小麦氮素转运及其对籽粒氮素贡献率的影响
对营养器官花前积累氮素转运的分析(表6)表明,茎鞘氮素再分配受灌浆期遮光影响最大,遮光处理的茎秆氮素转运量较CK平均下降 44.2%,其次为颖壳+穗轴,叶片氮素转运量受遮光影响最小。品种间比较,扶麦1228和生选6号营养器官花前积累氮素的转运量和转运效率受弱光的影响显著大于其他品种。总之,弱光状态下,氮素总积累量不变甚至略有上升,但却抑制了营养器官氮素的再分配。

表6 遮光对小麦花前贮藏氮素转运和花后氮素积累的影响
由于遮光抑制了叶片、茎鞘、颖壳等器官花前积累氮素的转运,花前营养器官积累的氮素对籽粒氮素的贡献率显著下降,各品种平均下降 7.4%,而花后吸收积累的氮素对籽粒氮素贡献率则平均上升7.4%,表明遮光条件下籽粒中的氮素对于花后氮素积累的依赖加大,而花前积累氮素的再分配对籽粒的贡献减少。
3 讨 论
3.1 花后遮光对干物质积累与转运的影响
花后遮光会降低小麦的干物质生产[11-12]。本试验中,遮光处理的小麦花后光合产物供应不足,各器官的干物质积累速率显著低于CK,导致灌浆中期和成熟期干物质积累量均显著小于CK,而且成熟期干物质积累量的降幅明显大于灌浆中期。对比两种类型品种,在CK条件下弱光钝感型品种干物质积累量在花后均衡增加,敏感型品种则表现为灌浆前期增加慢,后期增加快,灌浆前期和后期干物质积累进程的差异可能是不同品种对弱光响应不同的生理基础之一。
一般情况下,经济产量与生物产量呈正相关[22-23]。弱光钝感型品种成熟期干物质积累量下降幅度显著小于敏感型品种,与籽粒产量变化趋势一致。进一步分析表明,成熟期生物产量下降主要归因于各器官干物质积累量的减少,遮光处理下干物质在籽粒中的分配比例显著低于CK,而在茎、叶中的分配比例大于CK,弱光敏感型品种受影响大于弱光钝感型品种。
小麦籽粒灌浆速率取决于当前光合产物的供应能力及营养器官储存碳水化合物的再分配能力[24]。当光合产物供应在非生物胁迫下受限时,后者被认为是小麦维持稳定的籽粒灌浆速率的一种主要补偿机制[25]。遮光后小麦籽粒灌浆速率显著下降,快速增长期更明显,使得最终粒重显著降低。这主要归因于花后光合产物供应不足,虽然茎、叶中开花前贮藏的同化物再分配比例增大,使营养器官开花前贮存同化物的转运量以及对籽粒的贡献率增大,但是并不能弥补花后光合产物减少造成的损失,从而导致遮光处理生物产量以及籽粒产量下降。这说明弱光胁迫状态下,碳水化合物的合成受阻,籽粒灌浆源物质不足,小麦植株启动自我反馈机制,营养器官(叶、茎)中暂存的碳水化合物被调动,以备籽粒充实所需,但这种反馈调节有其阈值,终不能逆转源的不足,从而导致籽粒产量大幅度下降。就5个品种而言,弱光钝感型品种遮光处理花后光合物质生产能力较弱光敏感型相对增大,营养器官开花前贮藏的同化物再分配比例亦增大,两者是其生物量下降幅度较小的生理基础;另一方面,弱光钝感型品种灌浆速率快,灌浆进程较弱光敏感型品种短,相对早熟,可以作为品种筛选的一项优化指标。
3.2 花后遮光对氮素积累与转运的影响
小麦籽粒氮素20%左右来自花后直接吸收同化的氮素,而80%左右来自营养器官在花前贮存的氮素[26-27],所以籽粒中的大部分氮来源于开花前营养器官中贮藏氮素的再分配[28-29]。本试验中,遮光显著降低了各营养器官花前贮存氮素的转运量,弱化了其再分配能力,使残留在营养器官中的氮素显著大于CK。与牟会荣等[14]的研究不同的是本试验中营养器官花前贮存氮素的转运量与转运效率均显著小于CK,且遮光处理茎秆+叶鞘的转运效率下降最多,叶片的降幅相对较小,可能叶片转运效率依然存在补偿反馈机制来弥补转运量的下降,但是补偿效应不足以弥补其亏损,这可能是由于遮光时长与强度较大导致的。
遮光降低了花前贮存氮素转运量对籽粒氮素的贡献率,表明遮光后小麦籽粒氮素的积累更多地依赖于花后吸收的氮素,相对弱化了开花前营养器官中贮藏氮素的再分配。弱光敏感型品种花后氮素积累量提高幅度较小,其花后吸收的氮素对籽粒氮素的贡献率小于钝感型品种,这说明在受到胁迫后,弱光钝感型品种对于胁迫应激反应的调控能力大于敏感型品种。另一方面,遮光处理籽粒的氮素积累量显著下降,这主要归因于营养器官花前贮存氮素再分配能力下降,虽然遮光后花后氮素积累量提高,但是远不能弥补花前贮存氮素的转运量的下降造成的损失。就品种而言,籽粒氮素积累量下降幅度趋势与籽粒产量趋势一致,均表现为敏感型品种下降幅度大于钝感型品种。
此外,遮光处理花后氮素积累量升高,表明遮光相对促进了花后小麦植株氮素吸收,可能与遮光后小麦植株(根系+地上部)衰老进程延缓、相关酶活性相对升高有关[30],但是遮光后根系和地上部衰老进程与氮素吸收之间的联系以及作用机制还需进一步研究。另一方面,小麦植株吸收的氮素大部分来自土壤氮[31-32]。有研究表明,遮光降低了土壤氮素向籽粒的转移,但是提高了肥料氮素向籽粒的转移[12],选用氮肥利用效率高的品种和增加施氮量也许可以作为弱光条件下籽粒氮素减少的应对措施。
3.3 花后遮光对小麦籽粒产量的影响
弱光对小麦产量的影响结果不一致,可能因遮光强度和试验材料不同而存在差异[19,33]。在本试验中,开花期至成熟期遮去45%自然光强情况下,小麦籽粒产量显著下降,但是下降幅度因品种而异,弱光敏感型品种扶麦1228和生选6号的降幅显著大于弱光钝感型品种襄麦55和扬麦158。
从产量构成看,千粒重降低是遮光造成产量下降的主要原因。值得注意的是,千粒重下降幅度亦因品种而异,各品种间千粒重与籽粒产量降幅的变化趋势一致,虽然千粒重下降幅度低于籽粒产量降低幅度,但是均表现为生选6号>扶麦1228>郑麦9023>襄麦55>扬麦158。穗数和穗粒数不受遮光处理及遮光与品种互作的影响。刘希伟等[18]研究表明,遮光处理的小穗败育数增大,导致穗粒数减少,这与本试验结果不一致,可能与品种以及遮光强度不同有关。就5个品种而言,遮光后襄麦55和扬麦158籽粒产量降低幅度显著小于目前大面积推广的郑麦9023,表明此二品种耐荫性高于郑麦9023,可以作为耐荫品种进行试点推广。虽然扶麦1228的籽粒产量下降幅度较郑麦9023大,但是其自然生长状态下产量表现较好,具有一定的发展潜力。
4 结 论
灌浆期遮光影响了小麦植株中碳、氮的分配及再分配,遮光后光合产物合成受阻,对籽粒灌浆的供应不足,使其更多地依赖于营养器官中暂存的碳水化合物的再分配;而籽粒氮素积累则更多的依赖于花后吸收的氮素,营养器官花前贮存氮素的转运量显著下降。遮光后小麦籽粒产量显著下降,但是襄麦55和扬麦158籽粒产量降低幅度显著小于郑麦9023,表明品种耐荫性高于郑麦9023,可以作为耐荫品种进行试点推广。
猜你喜欢
广西糖业(2024年1期)2024-04-25 17:59:28
作物学报(2022年1期)2022-11-05 08:06:34
河北科技师范学院学报(2022年2期)2022-08-26 08:55:32
农业工程学报(2022年7期)2022-07-09 06:16:32
今日农业(2019年11期)2019-08-15 00:56:32
现代园艺(2017年23期)2018-01-18 06:57:39
浙江大学学报(农业与生命科学版)(2017年1期)2017-04-17 07:05:01
江苏农业科学(2017年1期)2017-02-27 10:14:38
动物营养学报(2015年10期)2015-12-01 06:45:19
苏州科技大学学报(工程技术版)(2015年3期)2015-02-28 16:21:12
