
坡垒生境调查及濒危原因分析
2020-03-05 09:39方发之陈素灵桂慧颖
热带林业 2020年4期
方发之,陈素灵,桂慧颖
海南省林业科学研究院(海南省红树林研究院),海南海口 571100
坡垒(Hopea hainanensis)为龙脑香科常绿大乔木,是海南热带沟谷雨林代表种和特有种[1]。坡垒树形优美,木质坚硬,结构致密,纹理交错,不易变形,切面油润,光泽性强,材色美观,特别耐腐性强,不受虫蛀。可与世界名材坤甸媲美,是极珍贵的工业用材和上等家具用材[2],为海南特类材。从20 世纪60 年代以来,坡垒列为禁伐树种,在1999 年国家林业局和农业部发布的《中国国家重点保护野生植物名录(第一批)》中,被列为国家Ⅰ级保护植物。但历经近60 年,坡垒种群资源状况一直得不到有效的改善,被《中国植物红皮书》定为濒危种,被IUCN 评估定为“濒危灭绝”(EN)等级[3]。导致植物濒危的原因包括外部因素(气候、生境、过度利用和采收)和内部因素(包括遗传力、生殖力、生活力、适应力的衰竭等)两大类,该研究通过实地调查坡垒野外生境、种群特征及种苗生长发育情况等,分析总结导致坡垒濒危的主要因素,为保护和人工栽培坡垒提供科学依据。
1 研究地概况
据资料记载,坡垒的野生分布区主要集中在海南中部的霸王岭、尖峰岭、黎母山等林区,地理坐标18°14′N~19°25′N,109°31′E~110°09′E。属热带季风性海洋气候,平均气温22℃,年均降水量为1900mm。但年内雨量分布不均匀,80%雨量集中在5 月~10 月,形成明显的干湿季。年平均相对湿度为80%~85%,年平均蒸发量为1824.1mm,土壤主要为花岗岩砖红壤[4-5]。
2 研究方法
采用文献检索、问询与野外实地调查相结合的方法,对主要分布区坡垒野外种群资源进行调查。记录其所处的地理坐标、海拔、坡度、坡位、坡向等。并以坡垒为中心,布设25m×40m 的调查样地,对样地内的植物进行每木调查,记录植物种类、高度、胸径(基径)、冠幅、林冠、郁闭度等群落特征,并调查样方内坡垒单株结实量、种子传播距离及幼苗数量等。
采集成熟种子,测量种子长度、直径、千粒重等特征,开展野外种子萌发试验,在野外人工坡垒林下及林窗内分别设置1m×1m 的试验小样方,样方内整地,播撒经催芽后的种子50 粒,设置覆盖枯枝落叶2cm、细土2cm、细土1cm 及对照4 个不同覆盖措施,每种处理3 个重复,35d 后调查统计种子萌发率。此外还对坡垒人工造林林分进行生长情况调查。
3 结果与分析
3.1 自然分布与伴生树种
坡垒是龙脑香科植物,地处热带北缘的中国海南和越南义安有分布。在海南岛主要分布于霸王岭、尖峰岭、黎母山、鹦哥岭、五指山、吊罗山等热带雨林区,分布面积约8820hm2[6]。野外调查发现,坡垒天然分布区主要集中于海南中部山区,其中霸王岭(266株)、尖峰岭(31 株)、黎母山(22 株)、鹦哥岭(5 株)。在各个调查样地中,均发现有橄榄(Canarium album)、黄杞(Engelhardia hainanensis)、青梅(Vatica mangachapoi)、 托盘青冈 (Cyclobalanopsis patelliformis)、乌柿(Diospyros cathapensis)、长序厚壳桂(Cryptocarya metcalfiana)等的树种伴生。此外在尖峰岭热带雨林中还发现坡垒常与中华厚壳桂(Cryptocarya chinensis)、红锥(Castanopsis hystrix)、卵叶桂 (Cinnamomum rigidissimum)、 鸡毛松(Podocarpus imbricatus)、 吊皮锥(Castanopsis kawakamii)、大叶蒲葵(Livistona saribus)等混生。
3.2 生境特征
3.2.1 气候
气温、水、日照等气象因子是决定植物分布的重要因子。根据收集的气象资料统计,坡垒分布区为典型的热带海洋性季风气候,分布区年均气温为21.5℃,最热月均温22.8℃(5 月),极端高温37.5℃,在冬季1 月均温12.2℃,极端低温1.1℃;年均降雨量1650mm~1900mm,平均降雨日数100d。干湿季分明,5 月~10 月为雨季,11 月~次年4 月为旱季。季风对坡垒分布区的降水量分配影响很大,使月、季、时段降水量出现明显的差异。坡垒在1.1℃能正常生长,具有一定抗寒能力,但在气温骤降到-1℃时死亡[7]。
3.2.2 海拔
坡垒分布的海拔范围大致在300m~1100m 之间,其在600m~700m 海拔范围内分布最多,表明坡垒的分布范围较广、生态幅较宽[8]。在人工种植中,坡垒在海南的乐东、定安、澄迈、儋州等海拔低于100m 的台地均能正常生长。
3.2.3 地形因子
对霸王岭(266 株)坡垒的实地调查发现,长于山脊的有48 株,占18.0%;山谷的有96 株,占36.1%;溪涧两旁边的有122 株,占45.9%。其中山谷与溪涧共218 株,占82%,说明坡垒喜生于温暖静风,空气湿度较大的山谷与溪涧边。
3.3 种群特征
调查发现,野生坡垒成材树(胸径≥20cm)均单株零星地分布于天然林中,未发现片林形成的群落。324 株成材坡垒均处于所在群落的最顶层,说明坡垒在其所在的群落种间竟争较强。此外调查还发现,散落的坡垒种子具有较高萌发率,坡垒母树周边约50m~100m 范围内,株高3cm~5cm 的幼苗总数约有280 株~510 株,幼苗密度最高达36 株/m2。树高0.5m~1.5m 的幼树仅有2 到3 株,均生长于石头缝隙或石头下方。说明在热带雨林中,由于林木和杂草灌木茂密,种间竟争激烈,坡垒幼苗竞争和适应能力弱,易被淘汰,而石头周边林木和杂草灌木稀疏,能提供坡垒生长空间,并且坡垒在石缝有较强适应能力,这与上述有68.7%的成材树分布石头边或石缝基本相吻合。调查中则未发现高1.5m 以上的坡垒未成材植株,可推测坡垒野外自然更新能力较弱。
3.4 繁育特征
人工种植的坡垒7a 可开花结果,花期为5 月至10 月上旬,种子成熟期为次年3 月上旬到5 月上旬。坡垒种子为坚果类,带翅。种子产量存在大小年之分,大年时单株产果量达10kg~30kg。种子近似心形,长约1.1cm~1.6cm,直径0.7cm~1.2cm,带萼片千粒重1025g~1109g,除去萼片千粒重650g~849g。种子成熟后易脱落,随风飘落。飘落至阴湿环境中的种子在5d 后可发芽。至开阔干燥之处,种子则失去水分难以萌发。60%~70%种子飘落在距母株60m 内,表明坡垒种子虽带翅,但其种子质量相对重,传播距离有限。
不同郁闭度不同覆盖措施对坡垒种子发芽率的影响结果见表1。
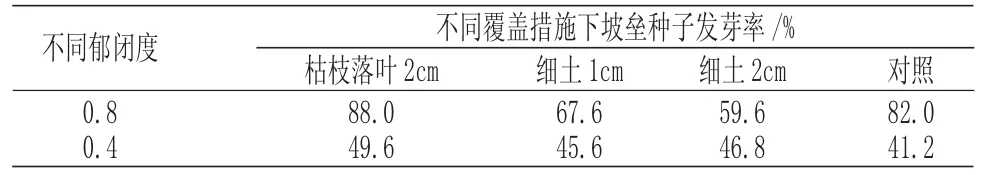
表1 不同郁闭度及覆盖措施下坡垒种子发芽率Tab.1 The seed germination rate of Hopea hainanensis under different canopy density and mulching measures
由表1 可知,所有处理中,坡垒林下被枯枝落叶覆盖2cm 的坡垒种子发芽率最高,郁闭度0.4 下不覆盖的坡垒种子发芽率最低。郁闭度对坡垒种子萌发有显著影响,郁闭度0.8 的各项措施发芽率均高于郁闭度0.4 下发芽率。这可能与萌发温度有关,郁闭度小的区域光照更强,蒸发量更大,坡垒种子经照晒脱水后失去生活力,导致种子未能萌发。此外对比不同郁闭度下的各项覆盖措施发芽率均可发现,枯枝落叶覆盖2cm 处理的发芽率均高于对照处理,而细土覆盖则表现为在坡垒林下发芽率低于对照,在林窗下发芽率高于对照。这可能与枯枝落叶层质量较轻、疏松多孔、空气流通良好有关,且适当的覆盖使得坡垒种子避免阳光直射而过分失水。
3.5 生长特征
从表2 可看出,坡垒幼苗在郁闭度0.4 的以上林下生长缓慢,并随着郁闭度的增加而降低,而保存率则相反,说明幼苗生长需要阳光,但光照强的地方种间竞争激烈,坡垒幼苗容易被淘汰。在东方市江边造林中采用不套种不抚育措施的保存率逐年降低,同样也说明这个问题。从东方市江边(套种农作物)的调查结果来看,人工种植的坡垒3a 后,其年生长旺盛,第4 年至第7 年,树高年均生长量为0.9m~1.3m。杨德军等[9]对42 年生坡垒生长观测也发现,坡垒树高、胸径生长速度较为均匀,其生长速度没有出现较大波动,最大高年生长量0.95m,最大胸径年生长量1.4cm。因此,坡垒能成片林成长,并有长时期旺盛的生长力。

表2 不同造林区域及条件下的坡垒生长情况Tab.2 The growth of Hopea hainanensis in different afforestation areas and conditions
3.6 人类活动的影响
(1)毁林开垦种植农作物,导致坡垒生境萎缩。解放初期海南原始热带雨林面积为120 万hm2,森林覆盖率35.5%。从20 世纪50 年代起,由于过量采伐、毁林垦植、毁林开荒、乱砍滥伐和森林火灾等人为破坏,至1978 年森林资源清查时天然林仅存40万hm2,森林覆盖率12.0%。1987 年下降到24.4 万hm2,森林覆盖率7.2%。到1999 年海南原始热带雨林面积仅13.3 万hm2,森林覆盖率4%。58%的天然林郁闭度从0.8 下降至0.4~0.5。大面积原始热带雨林被开垦和破坏,使坡垒的生境逐步退化丧失和碎片化,导致繁殖能力下降,种群数量不断减少。
(2)坡垒被偷砍盗伐,成材树数量锐减。坡垒木材材质坚硬,耐腐力极强,是极珍贵的上等家具用材。2012 年至2013 年间,黑市上交易价格达100 元/kg 至200 元/kg,在经济利益的驱使下不法分子偷砍滥伐,导致野生坡垒母株数量锐减。近些年来,执法部门对偷砍滥伐行为进行严厉打击,偷砍盗伐的行为受到厄制,坡垒资源得到有效保护。
4 濒危原因
造成物种濒危的外界因素主要为生境退化丧失、人类对生物资源的过度开采利用、外来种入侵、全球气候变化等,以及物种繁殖力低、种群小与种群密度低、地理分布狭窄、栖息地特化等与植物自身相关的易导致其濒危的灭绝特征[10-11]。坡垒在海南的分布范围较广、生态幅较宽,同时有较大结实量,种子发芽率较高,因此不存在自身繁殖能力上的问题。根据调查研究结果,导致坡垒濒危的原因归结为下列4 点:
(1)人类破坏是导致坡垒濒危的主要原因,从1952 年至1999 年间,由于不合理的人类活动,海南原始热带雨林面积从120 万hm2锐减至13.3 万hm2。近90%的热带雨林遭受破坏,坡垒群落作为热带雨林顶级群落代表之一,其生境的剧烈收缩,加之不法分子的偷砍滥伐,使坡垒母株数量锐减。
(2)坡垒生长特性也是导致濒危的原因之一,在热带雨林中各物种种间竞争较为激烈,而坡垒幼苗期生长极其缓慢,虽有大量幼苗分布,但坡垒幼苗竞争力弱易被淘汰,导致仅存的成年坡垒植株较少,种群野外自然更新能力弱。
(3)种群密度低,致使种群自然恢复与发展缓慢。野生坡垒成材树均单株零星地分布于天然林中,未发现片林形成的群落,在8820hm2的范围内,调查仅发现324 株成材树,种群密度极低,导致种群繁殖能力与抗逆性弱,种群自然恢复与发展缓慢。
(4)坡垒种子及其萌发特性制约种群扩张。坡垒种子为带翅坚果,但种子质量相对较重传播距离有限,其种子集中脱落分布在距母株100m 的范围内,限制其种群生长范围的扩张。同时种子含水率高,在阴湿环境下适宜发芽,开阔干燥环境下则易被晒干而失去萌芽力,坡垒种子在自然环境下对萌芽条件有较高要求,制约种群的发展。
5 保护及发展建议
(1)加强坡垒的保护与宣传,恢复其生境与种群。一是加强启动坡垒专项保护,以林业主管部门为主体,开展野外极小种群保护行动,优先关注和保护坡垒等濒危物种。建立自然保护区和保护小区是有效保护濒危物种的方法之一[12],在集中分布地建立重点保护小区,定期巡视以保护种群生境,维护种群稳定,降低坡垒因种群过小而灭绝的风险。二是打击偷砍盗伐行为,保护好坡垒成材母树。三是积极开展珍稀濒危植物的宣传教育工作,增强公众保护意识和各渠道宣传是保护珍稀濒危植物的有效手段之一[13],相关职能部门应充分利用各种媒体,采用多种形式,广泛开展宣传教育活动,普及保护珍稀濒危植物的重要性及方式方法,在民众中树立保护共识,提高参与保护行动积极性,使坡垒等珍稀濒危植物保护工作成为民众的自觉行为。
(2)加大人工造林力度,扩大种群资源。坡垒是特类用材树种,其材质好,用途广,经济价值极高,是优良的国家储备林树种,且人工造林的纯林林分生长快,是今后海南发展珍贵乡土树种的主要造林树种。建议采取以下措施扩大坡垒种群资源:一是大力发展以坡垒等珍贵乡土树种代替生态、经济效益低下的多代桉树林分的更新改造模式。二是在重要生态区位的桉树等人工公益林内,应用坡垒等珍贵乡土树种进行补植补造。三是将坡垒作为四旁绿化树种进行推广种植。坡垒是常绿乔树种,其冠形优美,应用于园林绿化,既可提升绿化成效,又可增加优质木材储备,扩大坡垒种群数量,保存坡垒种质资源。
猜你喜欢
中国农村水利水电(2021年11期)2021-11-29
风景园林(2021年2期)2021-04-24
绿色科技(2021年1期)2021-02-22
落叶果树(2021年6期)2021-02-12
热带作物学报(2020年6期)2020-08-04
福建林业(2020年5期)2020-03-18
农业与技术(2019年19期)2019-11-11
热带农业科学(2019年2期)2019-04-28
农民致富之友(2018年9期)2018-06-27
体育科技(2016年2期)2016-02-28
