
大马士革Ⅲ玫瑰蕾期净光合速率日变化特征及其与生理生态因子的关系
2020-03-03 14:37刘雷黄怡赵丹母欢索朗次珍陈红梦李志鹏杨玉霞
江苏农业科学 2020年22期
刘雷 黄怡 赵丹 母欢 索朗次珍 陈红梦 李志鹏 杨玉霞
摘要:以绵阳地区同等立地栽培条件下大马士革Ⅲ玫瑰为研究材料,采用LI-6400XT便携式光合仪测定其蕾期净光合速率(Pn)、蒸腾速率(Tr)日变化过程,同步测定空气CO2浓度(Ca)、气温(Ta)、胞间CO2浓度(Ci)、叶片温度(Tl)、光合有效辐射(Pra)、相对湿度(Rh)、气孔导度(Gs)和叶面水汽压亏缺(Vpdl)等相关生理和生态因子,采用DPS7.05对测得数据进行相关分析、逐步多元回归分析及通径分析,探讨大马士革Ⅲ玫瑰净光合速率与上述生理生态因子之间的关系。结果表明,大马士革Ⅲ玫瑰蕾期净光合速率呈非典型双峰型曲线,没有“光合午休”情况;偏相关分析得出,净光合速率与胞间CO2浓度、空气CO2浓度、叶片温度、相对湿度、气孔导度、空气温度以及叶面水汽压亏缺呈极显著相关关系;逐步多元回归分析得出,Pn与各生理和生态因子的回归方程为Y=113.41+2.19Tr+81.37Gs-0.04Ci-1.58Tl+0.12Rh-0.21Ca+5.55Vpdl+0.51WUE(R2=0.9999);影响大马士革Ⅲ玫瑰蕾期净光合速率的主要因素为Tl、Ta、Rh、Ca、Ta、Ci和Gs。该项研究为大马士革Ⅲ玫瑰的规范化丰产栽培管理以及高光效品种(系)选育提供了更多研究基础。
关键词:大马士革Ⅲ玫瑰;净光合速率;生理生态因子;相关分析;逐步多元回归
中图分类号:S685.120.1文献标志码:A
文章編号:1002-1302(2020)22-0139-05
作者简介:刘雷(1980—),男,四川彭州人,硕士,副研究员,主要从事微生物学、植物资源评价与利用、农业产业化与标准化等研究工作。E-mail:33020897@qq.com。
通信作者:杨玉霞,博士,研究员,主要从事中药材遗传育种、资源评价与利用等研究。E-mail:yangyuxia-7@163.com。
大马士革玫瑰(RosadamasceneMill)为蔷薇科蔷薇属纯甜型油用玫瑰品种。因其鲜花提炼的玫瑰精油纯正、香气浓郁、质量稳定,符合ISO9842国际质量标准,一直为国际市场主流玫瑰精油品种之一[1-2]。其精油价格高于黄金数倍,被誉为“液体黄金”,是国际奢侈香水、化妆美容、保健医疗、烟草和食品添加剂等行业的主要原材料。20世纪80年代,大马士革玫瑰开始引入中国,现已在陕西省渭南和商周[3]、浙江省安吉市[4]、湖北省西部地区[5]、北京市房山[6]、四川省攀西地区[7]、四川省绵阳[8]等地成功引种栽培并推广。大马士革Ⅲ玫瑰(Rosadamascenetyigintipetala)是从大马士革玫瑰中选育出来的更为优良的油用玫瑰品种(系),它既传承了大马士革玫瑰的优点,同时在生长适应性、抵御抗病虫害力、鲜花产量和精油得率上均有所提升,广泛用于栽培生产[7]。
国内外学者对大马士革玫瑰的研究主要集中在生产栽培[4-5,7]、精油化学成分与提取工艺[8-9]、精油的次生代谢调控[10-11]等方面。光合作用是大马士革玫瑰生长发育的重要生理指标,也是鲜花产量形成的基础,至今仍未见对大马士革Ⅲ玫瑰光合特性及其与生理生态因子相关性的研究报道。因此,研究大马士革Ⅲ玫瑰光合特性及与生理生态因子的相互关系,可探究其光合潜能,对大田栽培技术进行指导以提高鲜花产量,也是开展高光效育种工作的主要理论基础[12-13]。本试验对大马士革Ⅲ玫瑰蕾期叶片净光合速率、蒸腾速率和水分利用率全天候变化进行研究,分析生理生态因素等对其光合特性的影响,以期丰富大马士革Ⅲ玫瑰的系统研究,同时也为其优质高产栽培、有效管理及高光效新品种(系)选育提供理论依据。
1材料与方法
1.1试验地及材料
试验于2016年4月20日进行。试验地点位于四川绵阳游仙区柏林镇金马村宏林玫瑰开发有限公司大马士革Ⅲ玫瑰示范基地内,年均气温16.4℃,平均降水量675.3mm,年均日照时数2527.9h,年日照百分率58.3%,属于亚热带湿润型季风气候区。供试品种于2010年定植于绵阳市宏林玫瑰开发有限公司玫瑰种植资源圃内,经绵阳师范学院生态安全与保护四川省重点实验室陈艳教授鉴定为蔷薇科蔷薇属植物大马士革Ⅲ玫瑰(Rosadamascenetyigintipetala),试验地为沙质壤土,立地栽培及水肥管理条件相同,植物正常生长。
1.2生理与生态指标的测定
2016年4月下旬,晴天偶见有云,采用美国产LI-6400XT便携式光合仪在田间活体测定大马士革Ⅲ玫瑰蕾期叶片净光合速率和蒸腾速率日变化值,并同步测定和计算有关生理和生态因子日变化值,包括空气CO2浓度、叶面水汽压亏缺、气温、胞间CO2浓度、叶片温度、气孔导度、光合有效辐射和相对湿度等参数的变化。水分利用效率(WUE)=净光合速率/蒸腾速率。选择株龄、立地条件、栽培管理和长势基本一致的植株3株,自上而下随机选择最外层主枝5~7张完整健康叶片进行测定,每株测5张叶,累计测定15次,08:00—18:00每整点测量1次,每个时间段均重复测定3次。最后用去掉最大值和最小值后的13次测定的平均值作图和统计分析。
1.3统计分析
利用测定和计算数据,采用Excel对大马士革Ⅲ玫瑰净光合速率及相关生理生态因子日变化作图,采用DPS7.05对光合蒸腾值及相关生理生态因子进行相关分析、逐步多元回归分析和通径分析。
2结果与分析
2.1光合作用日变化
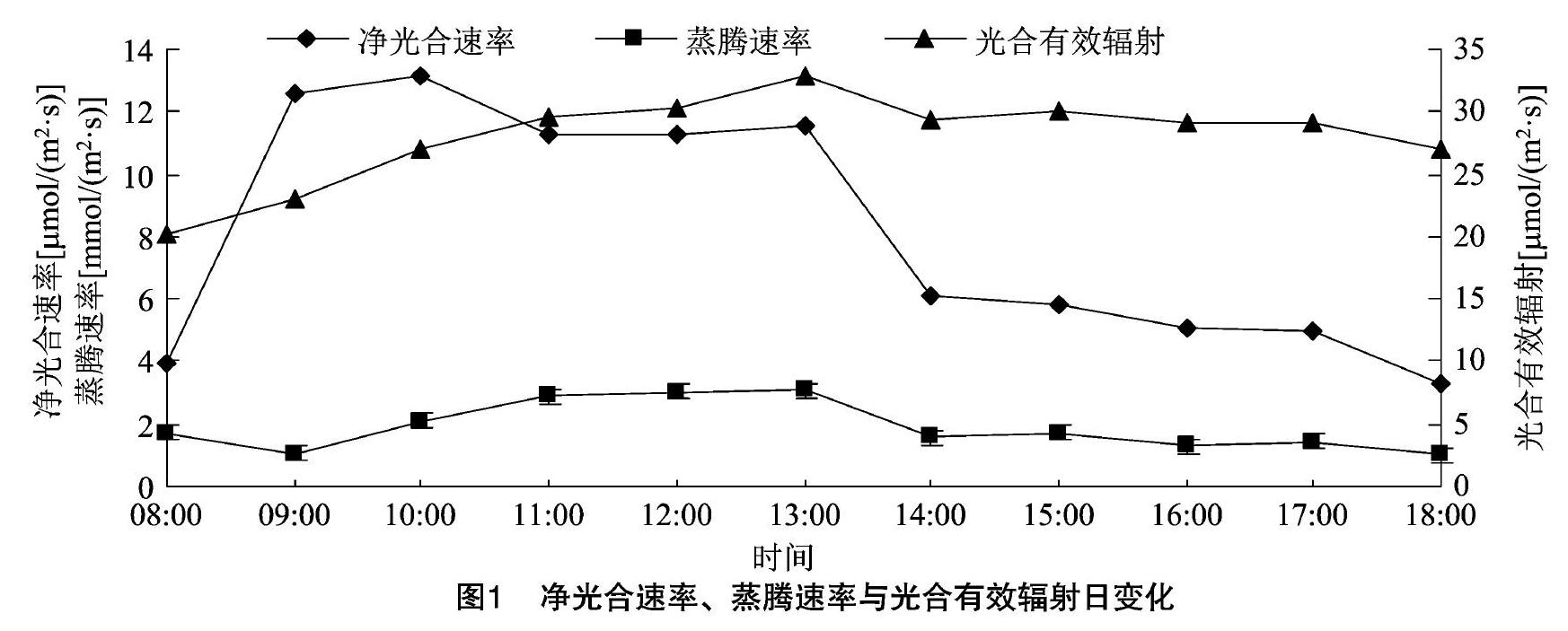
2.1.1净光合速率、蒸腾速率及光合有效辐射日变化
自然条件下,大马士革Ⅲ玫瑰蕾期光合蒸腾特性及生理生态因子的日变化见图1至图4。从图1可以看出,其净光合速率日变化呈非典型双峰型曲线,从08:00起Pn值持续上升,10:00达到最高值,此后又逐步开始下降,12:00达到一个低值,12:00—13:00间略有回升,此后就一直处于下降状态。蒸腾速率Tr值08:00—09:00略有下降,09:00—13:00之间持续上升,13:00达到峰值,与Pn值一样,此后就一直处于下降状态。Pra值日变化为单峰型曲线,08:00—13:00持续上升,13:00以后持续下降,峰值出现在13:00。
[FK(W11][TPLL111.tif;S+3mm][
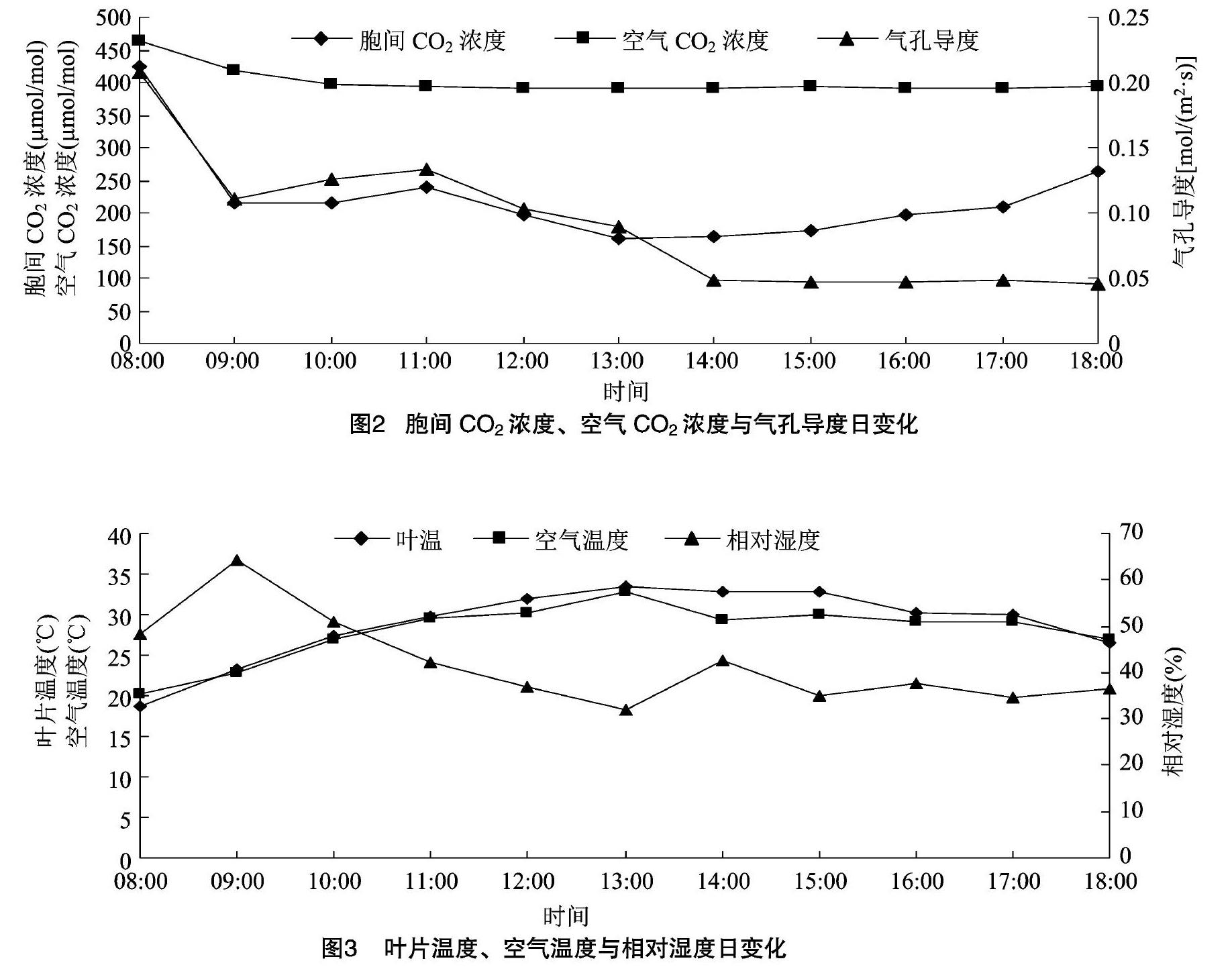
2.1.2胞间CO2浓度、空气CO2浓度与气孔导度日变化
从图2可以看出,胞间CO2浓度峰值出现在08:00,08:00—09:00迅速下降,09:00—11:00又有所回升,11:00—13:00又开始下降,13:00—18:00逐步开始回升。空气CO2浓度最高值也出现在08:00,08:00—10:00之间有一个相对明显的下降趋势,10:00—18:00变化趋势趋于平稳。气孔导度在08:00—13:00之间,变化趋势与胞间CO2浓度基本一致,14:00—18:00之间,变化趋势趋于平稳。[HJ2mm]
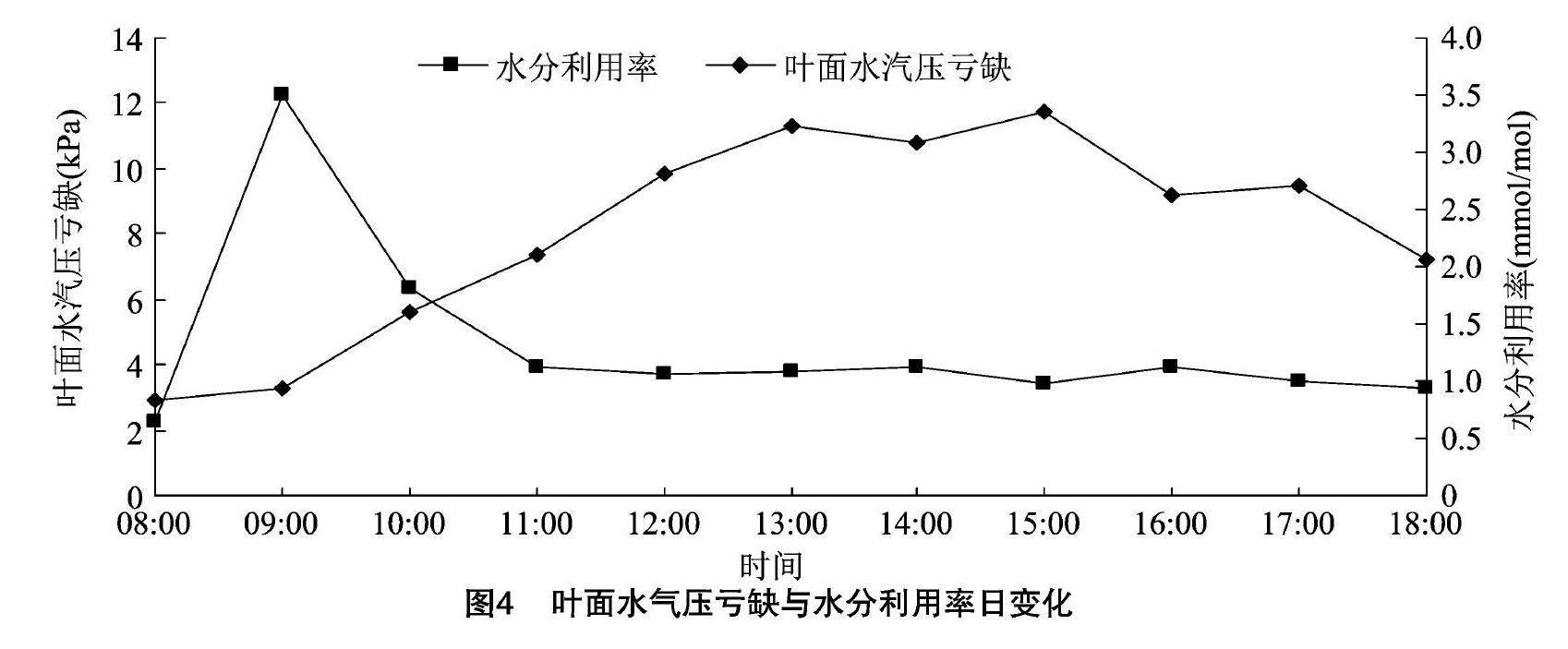
2.1.3叶片温度、空气温度与相对湿度日变化
从图3可以看出,叶片温度和空气温度的变化趋势基本一致,08:00—13:002个温度值均持续上升,13:00达到峰值,13:00—18:00逐步下降,且相同时间点上叶片温度和空气温度数值相差不大。相对湿度在09:00、14:00各有1个明显的峰值,最大值出现在9:00,属双峰型变化曲线。
2.1.4叶面水汽压亏缺与水分利用率日变化
从图4可以看出,水分利用率值08:00—09:00迅速上升,09:00出现一个明显峰值,09:00—11:00迅速下降,11:00之后變化趋势趋于平稳,属于单峰型变化曲线。叶面水汽压亏缺08:00—13:00持续上升,13:00—14:00略有下降,14:00—15:00略有上升,15:00之后持续下降,属于非典型双峰型曲线,2个峰值分别出现在13:00和15:00。
2.2大马士革Ⅲ玫瑰光合及蒸腾特性与生理生态因子间的相关分析
大马士革Ⅲ玫瑰净光合速率、蒸腾速率、水分利用率与环境因子的相关系数见表1。从表1可以看出,净光合速率与8个环境因子之间简单相关均未达到显著水平,但与气孔导度、叶片温度、叶面水汽压亏缺、空气温度、相对湿度以及胞间CO2浓度的偏相关均达极显著水平。蒸腾速率与8个环境因子之间简单相关也均未达到显著水平,但与气孔导度、空气温度、光合有效辐射以及叶面水汽压亏缺的偏相关均达极显著水平。水分利用率与相对湿度的简单相关达极显著水平,与胞间CO2浓度、叶片温度、相对湿度、空气CO2浓度以及叶面水汽压亏缺的偏相关均达极显著水平。
2.3逐步回归分析
以Tr、Gs、Ci、Tl、Ta、Pra、Rh、Ca、Vpdl和WUE为自变量,Pn为因变量,运用逐步多元回归分析法计算拟合度最佳的回归方程,Y=113.41+2.19Tr+81.37Gs-0.04Ci-1.58Tl+0.12Rh-0.21Ca+5.55Vpdl+0.51WUE(R2=0.9999,F=55437.23,P=0.00),表明大马士革Ⅲ玫瑰净光合速率Pn与各生理和生态因子间存在极显著的线性关系,在影响Pn的各个因子中Gs贡献最大,其次为Vpdl和Tr,Ci贡献最小。当影响Pn的其他因素均维持在平均水平时,Tr、Gs、Rh、Vpdl和WUE每增加1个单位,Pn值将分别增加2.19、81.37、0.12、5.55和0.51,Ci、Tl、Ca每增加1个单位,Pn值将分别减少0.04、1.58和0.21。决定系数R2=0.9999,表明这10个生理和生态因子可以解释Pn值变化的99.99%以上,还有0.01%的变化由其他因素决定。所得回归方程能够很好地预判Pn值线性变化。
2.4通径分析
为找出大马士革Ⅲ玫瑰蕾期光合特性的主要影响因子,分析生理生态因子的间接与直接作用效应,对所得数据作通径分析(表2)。结果表明,影响大马士革Ⅲ玫瑰净光合速率的主要生理和环境生态因子为Gs、Ci、Tl、Ta、Rh和Ca,各生理和生态因子对Pn作用大小依次为Ta>Rh>Ca>Ta>Ci>Gs。Tl对Pn的直接作用最大,而其通过Ca和Ta对Pn间接负作用较大;Rh对Pn的直接作用居第2位,而其通过Tl的间接负作用最大;Ca对Pn的直接作用居第3位,而其通过Tl的间接负作用也是最大;Ta对Pn的直接作用居第4位,而其通过Tl的间接正作用最大;Ci和Gs对Pn的直接作用最小。本试验的决定系数为0.99946,剩余通径系数为0.02318,表明影响Pn的主要生理生态因素已经包括在内。
3讨论与结论
高等植物的光合作用是一个系统的生理过程,生态环境因子对其有直接影响,而且植物的相关生理因子受到生态环境因子间接影响后也会进一步影响最终光合结果[14]。先前研究得出净光合速率与生理生态因子的关系不尽相同[15-17]。本研究偏相关分析表明,气孔导度、空气温度、胞间CO2浓度、叶片温度、相对湿度、叶面水汽压亏缺与净光合速率有极显著相关性。逐步多元回归分析表明,影响净光合速率的主要影响因子为气孔导度、叶面水汽压亏缺、蒸腾速率、相对湿度和叶片温度等。通径分析表明,影响净光合速率的主要影响因子为空气温度、相对湿度、空气CO2浓度、叶片温度、胞间CO2浓度和气孔导度。可见上述3种分析方法得出的生理生态因子与大马士革Ⅲ玫瑰光合特性的关系不完全相同,究其原因,偏相关分析没有考量其余变量对考察变量的影响,考察的是2个变量之间真实关联程度。逐步多元回归分析是从1个自变量开始,考察自变量对因变量作用的显著程度,从大到小依次逐个引入回归方程,引入新的自变量后的会对之前不显著的自变量进行剔除,最终引入的全部自变量对Y值影响均显著,用所得回归方程对因变量进行预测的统计学方法。而通径分析则是回归和相关分析的纵深与拓展,不仅获得自变量对因变量的影响程度,还可以获得性状间的直接和间接作用关系。
植物光合作用是一个系统而交杂的生命过程,净光合速率日变化特征不仅与自身叶绿素含量、叶片厚度和成熟度密切相关,也是同各种相关生理生态因子相互作用的最后结果[18],研究光合日变化特征可用于分析植物生长环境中的限制因素[19]。根据植物光合最初同化产物的不同,可分为C3和C4植物两大类,其Pn日变化一般呈现“双峰”型或“单峰”型的规律性[20-23]。本研究得出大马士革Ⅲ玫瑰蕾期Pn日變化为非典型双峰型,09:00—13:00都保持较高的净光合速率,峰值出现在10:00左右,蒸腾速率变幅趋于平缓,在12:00—13:00处于峰值,水分利用率在09:00出现1个突出峰值,其余时间一直较低。研究结果表明,水稻[13]、大豆[24]、小麦[25]、柑橘[26]等C3植物为“双峰型”净光合速率日变化,而玉米[27]、甘蔗[28]、高粱[29]等C4植物则为“单峰型”。大马士革Ⅲ玫瑰是否为C3植物,还有待于解剖和生理结构方面的进一步研究。C3植物光能利用率较C4植物低,因此改造大马士革Ⅲ玫瑰植株株型,或结合现代生物技术通过增加C4光合代谢途径的表达,提高其光能利用率将是后期高光效新品种(系)选育时必须兼顾考虑的问题。
参考文献:
[1]张睿,魏安智,杨途熙,等.商丘产大马士革玫瑰精油研究[J].西北植物学报,2005,25(7):1477-1479.
[2]杨新征,杨德,张跃华.玫瑰的价值及其开发前景[J].特种经济动植物,2004,7(2):24-26.
[3]张睿,魏安智,杨途熙,等.秦渭玫瑰精油研究[J].西北植物学报,2003,23(11):1991-1993.
[4]王越,唐永红,吴丹青,等.浙江安吉地区大马士革玫瑰引种栽培[J].上海交通大学学报(农业科学版),2009,27(3):226-230.
[5]陈燚格,陈芳清,谢宗强,等.鄂西地区大马士革Ⅲ玫瑰的引种栽培[J].中国农学通报,2012,28(16):225-232.
[6]郭晓春.北京房山大马士革玫瑰枝叶化学成分及化感作用[D].北京:北京林业大学,2013:11-12.
[7]徐云,苏春江.攀西地区大马士革Ⅲ玫瑰引种栽培及产业化前景[J].山地学报,2006,24(5):636-640.
[8]刘雷,何江,唐宏波,等.引种大马士革Ⅲ玫瑰精油化学成分研究[J].安徽农业科学,2014,42(11):3372-3373.
[9]周围,王文军,刘倩倩.基于GC-MS与代谢组学探讨产地对大马士革玫瑰油化学组成的影响[J].香料香精化妆品,2019(4):1-6.
[10]张强,张晓伟,孟月娥,等.大马士革玫瑰RdGASA4-like基因启动子的分离及其在甘蓝中的瞬时表达分析[J].中国农学通报,2011,27(8):123-127.
[11]Jean-LouisM,RocciaA,Jean-ClaudeC,etal.Biosynthesisofmonoterpenescentcompoundsinroses[J].Science,2015,349(6243):81-83.
[12]张耀文,赵小光,光周博,等.油菜光合生理研究及高光效育种研究进展[J].中国农学通报,2017,33(3):44-51.
[13]牛丽芳,路铁刚,林浩.水稻高光效育种研究进展[J].生物技术进展,2014,4(3):153-157.
[14]张新慧,张恩和.当归叶片光合参数日变化及其与环境因子的关系[J].西北植物学报,2008,28(11):2314-2319.
[15]张美善,徐克章.西洋参叶片光合日变化与内生节奏的关系[J].吉林农业大学学报,2003,25(6):595-597.
[16]翁晓燕,蒋德安,陆庆,等.影响水稻叶片光合日变化因素的分析[J].中国水稻科学,1998,12(4):105-108.
[17]侍瑞,苏培玺,周紫鹃,等.高寒植物群落优势种光合日变化及其与环境因子的关系[J].中国沙漠,2019,39(4):46-53.
[18]张治安,杨福,陈展宇,等.菰叶片净光合速率日变化及其与环境因子的相互关系[J].中国农业科学,2006,39(3):502-509.
[19]马骏,唐海萍.北京世纪坛绿地国槐光合特征与生理生态因子关系[J].北京师范大学学报(自然科学版),2010,46(5):619-623.
[20]刘遵春,包东娥.“金光杏梅”叶片净光合速率与生理生态因子的关系[J].西北植物学报,2008,28(3):564-568.
[21]程林.枸骨冬青秋季光合生理特性研究[J].江苏农业科学,2015,43(12):221-223.
[22]李海梅,何兴元,宋力.3种灌木树种光合特性及影响因子研究[J].沈阳农业大学学报,2007,38(4):605-608.
[23]苏军德,勾晓华,曹宗英,等.祁连圆柏光合作用日变化特征及其与生理生态因子的关系[J].西北植物学报,2011,31(5):1011-1017.
[24]陈睿.季节性干旱对反枝苋和大豆光合特性及其竞争机制的影响[D].哈尔滨:东北农业大学,2019:16-17.
[25]孙旭生,林琪,刘义国,等.不同施氮量对超高产小麦灌浆期光合日变化的影响[J].华北农学报,2008,23(1):158-162.
[26]胡利明.柑橘光合特性研究及光合途径的初步探讨[D].武汉:华中农业大学,2007.
[27]张峰,周广胜.玉米农田冠层光合参数的多光谱遥感反演[J].植物生态学报,2014,38(7):710-719.
[28]孟丽萍,叶燕萍,王泽平,等.不同甘蔗新品系光合性状的比较研究[J].广西农业科学,2009,40(10):1292-1295.
[29]解婷婷,苏培玺.干旱区不同土地类型下甜高粱叶片光合特性和水分利用效率[J].中国农业科学,2011,44(2):271-279.
猜你喜欢
今日农业(2022年16期)2022-11-09
环球时报(2022-05-26)2022-05-26
中国化妆品(2018年6期)2018-07-09
中国集体经济(2016年26期)2016-11-19
环球时报(2012-01-31)2012-01-31
环球时报(2012-01-07)2012-01-07
