真光层海水过饱和甲烷的来源及机制探讨
2020-02-28 07:45张亭亭梁前勇赵静肖曦董一飞郭斌斌钟超吴学敏杨林
海洋地质与第四纪地质 2020年1期
张亭亭,梁前勇,赵静,肖曦,董一飞,郭斌斌,钟超,吴学敏,杨林
1. 中国地质调查局广州海洋地质调查局,自然资源部海底矿产资源重点实验室,广州 510075
2. 中国地质调查局天然气水合物工程技术中心,广州 510075
甲烷是大气重要的温室气体,其温室效应是CO2的28倍,可直接或间接地影响全球气候,对全球变暖贡献约占20%[1]。海洋是一个极其庞大的甲烷库,据估计海洋每年产生的甲烷量约占全球甲烷产生总量的30%[2-3],对大气甲烷的贡献可达2%~4%[4]。目前普遍认为甲烷主要由厌氧微生物产生,因此,对海洋甲烷循环的研究主要集中于厌氧沉积物中[3]。此外,真光层海水中也普遍存在高浓度甲烷现象,如黑海[5],波罗的海[6]、北极水域[7]等(表 1),并且部分海域的大洋区、陆架斜坡区和海湾区的真光层甲烷浓度都依次处于不同程度的过饱和状态[8]。海洋天然气水合物区真光层也存在甲烷异常现象,例如对南海北部陆坡天然气水合物区海水甲烷分布连续5年的调查发现,部分季节真光层海水甲烷浓度可高达20~30 nm·L-1[9]。由于真光层海水甲烷非常接近海气界面,甲烷氧化和扩散过程并不明显,可直接扩散到大气中[10-11],因此,真光层海水过饱和甲烷可能是引起大气甲烷含量变化的直接因素,是导致全球气候变暖的重要推手。
真光层是浮游植物进行有效光合作用的水层,其深度主要取决于海域的纬度、季节和海水的混浊度,在混浊的近岸海域,深度仅有数米;在透明度较大的热带海区,深度可达200 m以上。真光层相对于大气中的氧气是饱和或者稍微过饱和,并不利于厌氧微生物生成甲烷,这种尚未解释的海水甲烷过饱和现象被称为“海洋甲烷相悖论”[12-13]。很多研究者对真光层海水过饱和甲烷的来源机制进行了大量的研究,相比于厌氧沉积物,真光层海水甲烷的来源更为复杂,可能受水动力、化学生物等多种因素共同影响,并可能衍生出新的产甲烷代谢类型与机制。本文综合前人研究成果对真光层海水过饱和甲烷产生的来源进行梳理,并对远洋海域原位微生物可能参与的甲烷代谢机制进行探讨,希望为天然气水合物试采与开发时的基础环境评价提供借鉴,为了解海水甲烷对大气甲烷通量和全球变化的贡献提供理论依据。
1 真光层海水过饱和甲烷的来源
1.1 海底沉积物甲烷泄漏并运移到海水真光层
海底含有丰富的甲烷库,其以溶解态、气态或固态(天然气水合物)形式存在。此外,海底每年还产生85~300 Tg甲烷,大于90%的甲烷在排放到水圈之前都被微生物介导的甲烷厌氧氧化过程所消耗[3,14],所以一般海底甲烷泄漏量较小。然而,海底天然气水合物受到扰动或冷泉等特殊生态系统仍会释放出大量的甲烷,例如,调查发现天然气水合物钻探后,有大量气泡从废井冒出,并形成甲烷气体羽流[15]。在冷泉系统中,由于甲烷大量高速运移,其氧化效率较低,大量甲烷以溶解态或气态形式从沉积物中溢出,其中,溶解甲烷流出量范围可达1~1 200 mmol·m-2·d-1,在部分高流速冷泉环境中,每年每千米的甲烷流出量可达数千吨碳;气体甲烷还可能在冷泉上方形成气体羽流,部分羽流每年可以释放 10~100 t碳[16]。
从沉积物溢出水圈的甲烷是否能够到达真光层受区域性海水深度影响极大[17-18]。在较浅的水域中,甲烷氧化屏障较小,大部分甲烷从沉积物运输到水体表层[19-21],例如,在北极洋西伯利亚东部水深50 m的大陆架区域,底层甲烷向上运移直接导致大面积表层海水的甲烷过饱和[22];然而,通过模拟研究发现,在水深2 000 m的海底,即使有1 000个泥火山同时喷发甲烷(甲烷量为179 Tg·d-1),表层海水甲烷含量变化仍非常小[23]。由此推测,在较深的水域中,大部分甲烷被氧化或被密度跃层阻挡难以到达真光层。因此,真光层海水过饱和甲烷可能来源于海底沉积物(包括冷泉)的向上运移(图1),但此过程受区域海水深度、微生物氧化和密度跃层的影响较大[23-25]。
1.2 富甲烷河流输入导致海水真光层甲烷富集
在近岸海域,甲烷主要产生于河流富含有机质的厌氧沉积物中,随着河流冲淡水向海洋的入侵,由于贫甲烷海水的稀释、海气界面甲烷扩散作用、盐度/硫酸盐的升高导致甲烷的产生升高/氧化降低等因素影响[26],真光层甲烷浓度从沿岸到外海呈现逐渐降低的舌状分布[27-28],例如从珠江口到南海北部,珠江口高浓度甲烷的影响可延伸到70 km之外[26]。由此推测,海水真光层过饱和甲烷可能来源于河流的输入,但此来源可能仅仅局限于离海岸较近的区域(图1)。
1.3 真光层海水原位生物作用导致甲烷的产生
对海洋和深湖宽阔的水域研究发现,真光层水体过饱和的甲烷不能单独由海底或邻近河流供给[33-35],随着与海岸的距离越来越远,甲烷浓度可能由外海的原位生物主导(图1)[26]。
1.3.1 产甲烷古菌介导甲烷产生
产甲烷古菌是甲烷代谢微生物的主要类群,也是目前研究最清楚的产甲烷微生物之一。它们是一组严格的厌氧微生物,几乎都存在于所有类型的厌氧环境中[36-39]。那么产甲烷古菌如何在真光层海水中介导甲烷的产生?碳稳定同位素分析表明,它们可能存在于真光层海水的缺氧微环境中[40-41],如悬浮物[42]或下沉碎屑颗粒内部[43-44]、鱼等浮游动物的消化道或粪粒中[45-49]。最近,Schmale等[30]研究发现,波罗的海含氧层的甲烷富集与中型浮游生物内
产甲烷古菌有关。然而,在培养过程中获得的甲烷产出率太低,仍无法解释有氧层甲烷形成的原因。

表1 真光层海水甲烷过饱和现象的实例Table 1 Examples of methane supersaturation in euphotic zone of the ocean

图1 真光层海水过饱和甲烷可能的来源Fig.1 The possible sources of oversaturated methane in euphotic zone of the ocean
另一方面,产甲烷古菌也可存在于多种有氧环境中,且具有代谢活性[49-53](表2)。例如,通过16S rRNA基因序列的多元分析发现产甲烷古菌类群Methanocellales, Methanomicrobiales和Methanosarcinales可存活在有氧环境中[54]。综合多个研究发现,产甲烷古菌的3种代谢类型(氢营养型、甲基营养型和乙酸营养型[55])都可能参与有氧水体中甲烷的产生[49,52,56-57](表2),产甲烷古菌主要附着在藻类上与其共生,藻类为产甲烷古菌提供低氧环境和产甲烷底物[52]。例如,藻类可产生甲基型产物(如甲胺和二甲基硫化物),蓝藻还可以通过夜间固氮活动产生氢气[58],这都是产甲烷古菌参与甲烷生成时的重要底物。因此,真光层海水中的产甲烷古菌能够进行产甲烷代谢,它们可能存在于微厌氧环境中,也可能需要与藻类共同作用。
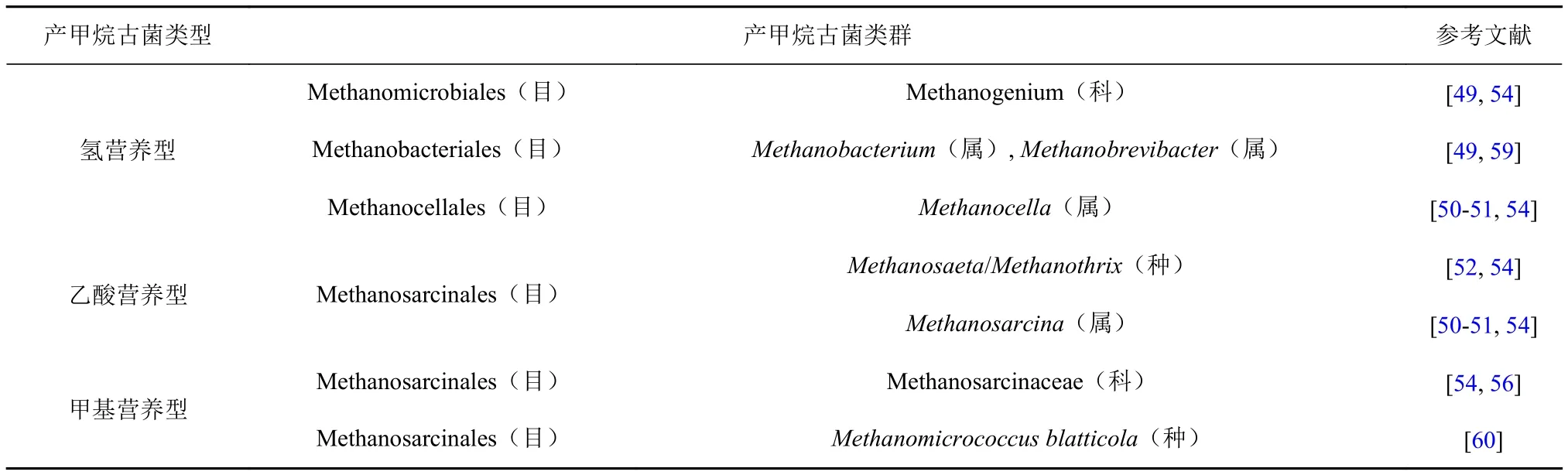
表2 存活于有氧环境下产甲烷古菌类群Table 2 The surviving methanogenic archaeal communities in oxic environments
1.3.2 细菌介导甲烷的产生
随着研究发现,产甲烷古菌并不是唯一的产甲烷生物,例如陆地植被、真菌和哺乳动物也可能在没有产甲烷古菌且有氧条件下产生甲烷[61-66],这打破了甲烷产生仅能由古菌介导的传统观念。在有氧水体中,细菌利用其高度的变异性,极有可能突破环境的限制产生甲烷,并衍生出新的产甲烷方式。例如,细菌可能以二甲基巯基丙酸内盐(DMSP)等甲基化合物为前体参与真光层海水甲烷产生。DMSP是海洋浅表层丰富的甲基化底物,由海洋浮游植物产生,是异养细菌的主要碳源[67]。在部分甲烷过饱和海域,DMSP与甲烷含量存在明显的耦合关系,由此推测,由藻类提供的DMSP可能再次被细菌利用并产生甲烷[31,68]。通过培养实验对比研究发现,加入底物DMSP的培养瓶中甲烷的含量明显增高,并且在此过程中细菌的丰度也大幅增加,且几乎检测不到古菌存在[69],这验证了细菌以DMSP为底物介导甲烷产生的可能性。此外,DMSP的降解可形成DMS(二甲基硫)[70-71],DMS的厌氧代谢可产生甲烷[72-73],实验验证发现表层海水DMS也能增加甲烷产量[32]。对热带寡营养西太平洋研究发现,DMSO(二甲基亚砜)与甲烷也存在正相关关系,DMSO也可能是甲烷产生的底物[68]。
磷酸酯(MPn)等有机磷酸化合物也可作为另一类甲烷产生前体。海洋浮游细菌以MPn作为磷源,而甲烷作为MPn分解的副产物[74]。且此代谢过程及其普遍,存在于许多微生物中[75],例如,培养实验发现,海洋α-变形菌门亚类群SAR11和假单胞菌属等都能够利用MPn产生甲烷[76-77]。海洋奇古菌Nitrosopumilus maritimus具有MPn生物合成的途径[78],且此古菌在海洋中的含量较为丰度[79-80],这为MPn的物质来源提供了保障。然而,自然条件下,并未在表层海水的颗粒物中检测到MPn的存在[81-82],这表明奇古菌在产生的MPn并不丰富[79,83],或者MPn不能大量积累。并且,测量发现MPn产甲烷速率过低,无法解释所观察到的甲烷过饱和现象[84]。随后,Repeta等[85]研究表明除了MPn,降解磷酸酯(Phn)也能导致甲烷的形成。虽然已知许多生物体可产生Phn[86],但是并非Phn降解都能产生甲烷[87],Phn的降解对真光层海水甲烷的贡献量仍然值得进一步探索。
1.3.3 藻类参与甲烷的产生
早有研究者提及甲烷原位产生可能与海洋藻类有关[88-89],随后研究也表明,甲烷浓度与初级生产力、叶绿素a有显著相关性[31,41,90]。藻类可能与细菌、产甲烷古菌协同作用参与真光层海水甲烷产生,由藻类产生DMSP、H2等底物,而细菌、产甲烷古菌利用这些底物介导甲烷产生[7,31,52,69]。此外,藻类也可能自身直接参与甲烷代谢,例如,蓝藻能够利用MPn获取磷,并释放副产物甲烷[87];在无产甲烷古菌的环境中,藻类能以碳酸氢盐和硫代甲基化合物作为碳前体直接产生甲烷[90]。因此,藻类可能与细菌、产甲烷古菌协同作用间接产甲烷,也可能直接产甲烷。
1.4 真光层海水非生物作用导致甲烷产生
在无生物作用下,光或氧化剂(如铁矿物质)可降解蛋白质、丙酮或抗坏血酸,产生并释放甲烷[91-93];有机硫化物也可在高氧条件下被铁(II/III)转化为甲烷[92]。因此推测,非生物作用可能也会引起真光层海水甲烷的产生。通过培养表层海水进一步证明发现,排除微生物影响后,DMS等有机化合物可以通过光化学途径产生甲烷[94],这表明在真光层海水中可能存在非生物引起的甲烷产生过程。然而,自然海水环境中的DMS含量远远低于实验中添加的DMS浓度,在没有甲基自由基前体的情况下,甲烷的光生成作用几乎检测不到,在现今的海洋中甲烷的光化学形成微乎其微[93]。因此,真光层海水中,非生物作用可能导致甲烷的产生,但其影响极小。
1.5 真光层海水过饱和甲烷现象形成的影响因素
真光层海水过饱和甲烷现象产生受甲烷形成因素和消耗因素共同作用。首先,真光层过饱和甲烷形成受区域性变化影响,在离海岸较近、深度较浅的海域,真光层海水甲烷可能来源于沉积物、冷泉或邻近河流,它们通过垂直或水平运移到达真光层;而对于远洋海域,甲烷可能受原位生物影响。此外,季节性变化是导致甲烷过饱和另一个重要因素,很多研究表明夏季更易出现有氧水体甲烷过饱和现象[30-31,95]。营养盐变化对甲烷过饱和也有重要影响,营养盐贫乏可能刺激甲烷产生,例如,西弗拉姆海峡由于具有寡营养的极地水域输入,藻类大量勃发,从而导致甲烷产生并富集[7];Karl等[74]认为磷酸盐缺乏的水域,细菌可利用MPn产生甲烷;而Damm等[69]认为寡硝酸富磷酸的水域有利于提高细菌竞争磷酸盐的能力,并促进甲烷产生。然而,Florez-Leiva等[32]研究富营养的沿海上升流发现,表层海水即使富含硝酸和磷酸,也可产生甲烷。因此,真光层过饱和甲烷现象的形成受区域、季节、营养盐等多种因素控制。
甲烷氧化是平衡海水甲烷过饱和状态的重要途径,也是一种调节甲烷从海水到大气通量的机制[96]。研究表明,当海水甲烷浓度达到一定阈值时,甲烷开始消耗[31],甲烷氧化速率与甲烷和氧气浓度正相关[97]。然而,甲烷的总氧化率仍较低,仅占甲烷总初级产量的1.5%[96],Tang等[57]与Murase和Sugimoto[98]认为可能是光照作用抑制了甲烷氧化;同时,Grossart等[52]通过分子标记方法发现甲烷氧化菌仅存在于温跃层以下的水体中,真光层甲烷氧化菌的缺失可能进一步降低了甲烷氧化效率。此外,甲烷扩散也是消耗真光层甲烷重要途径,但是其效率也非常低[29]。因此推测,甲烷氧化和扩散对真光层海水甲烷影响较小,只需要向真光层提供少量的甲烷,就可以维持真光层甲烷过饱和状态。
2 真光层海水原位微生物介导甲烷产生的代谢途径
综上,真光层海水过饱和甲烷可能来源于海底沉积物或河流,而对远洋海域,甲烷可能主要由原位微生物介导产生。那么,微生物如何在有氧真光层海水中产生甲烷?一种可能性是:主要由产甲烷古菌介导,它们依旧利用传统的产甲烷途径,存在于动物肠道、颗粒物等微厌氧环境中,或者自身产生抵抗氧气影响的能力[30,51-52];另一种可能性是,主要由细菌介导,它们避开常规途径所述的氧敏感酶的生物化学途径,转而使用新的产甲烷途径(图2)[7,10,95]。
在传统产甲烷途径中,碳载前体分子充当电子受体,并转化为最终产物甲烷,此过程主要在产甲烷古菌细胞内完成。产甲烷古菌通过产甲烷代谢获取能量,其代谢类型可分为3种:CO2还原途径、甲基裂解途径和乙酸发酵途径[99]。在CO2还原途径中,CO2首先被还原为甲酰基(-CHO),并转移到C1载体甲烷呋喃(MFR)上形成CHO-MFR。甲酰基又转移到第二个C1载体四氢甲基喋呤(H4MPT),并依次被还原为次甲基(≡CH)、亚甲基(=CH)和甲基(-CH3),甲基再次被转移到第三个C1载体巯基辅酶M(HS-CoM),产甲烷的最后一步是产生最末端产物甲烷。在甲基裂解途径中,甲基底物可包含甲醇(CH3OH)、甲氨(CH3NH2)、甲基硫((CH3)2S)等,甲基底物中的甲基由甲基转移复合体激活并转运到HS-CoM,形成CH3-S-CoM,最后转化为甲烷。在乙酸发酵途径中,乙酸可通过两种途径形成乙酰辅酶A(CH3COSCoA),之后乙酸上的羧基被氧化成CO2,乙酸中的甲基依次被转移到C1载体四氢甲基喋呤(H4MPT)和巯基辅酶M(HS-CoM),最后还原形成CH4[100-101]。所有的产甲烷古菌产甲烷代谢途径的最后一步都需要甲基辅酶M还原酶(Mcr)参与,此酶由 3 个亚基(α2β2γ2)和辅酶 F430组成[102]。此外,Mcr也是甲烷厌氧氧化的关键功能酶[103],编码Mcr α亚基的功能基因(mcrA)是研究产甲烷古菌多样性的重要标志物[104]。由于Mcr对有氧环境中的O2衍生物或活性氧(ROS)及其敏感[105],因此,由此酶催化还原反应必须在严格厌氧条件下进行。然而,研究表明部分产甲烷古菌已演化出特定的功能来抵制氧气的影响。例如,沙漠土壤中的Methanosarcina和Methanocella即使在含氧条件下也可以转录mcrA基因,同时转录氧解毒过氧化氢酶基因来抵抗氧气影响[51]。因此,真光层海水的产甲烷古菌使用传统产甲烷途径代谢的同时,也可能转录抗氧基因,从而为产甲烷古菌在真光层合成甲烷提供前提条件。
细菌能够参与甲烷产生,但仍缺少传统产甲烷途径中的关键功能酶(即辅酶M甲基转移酶和甲基辅酶M还原酶)[95],因此推测,细菌可能利用新的替代途径产生甲烷。目前,研究较为普遍是C-S键裂解途径和C-P键裂解途径,这两个过程分别需要对C-S键和C-P键进行裂解[7,74]。在C-S键裂解途径中,DMSP首先去甲基形成甲基巯基丙酸盐,并转移到C1载体巯基辅酶A(HS-CoA)上,依次转化为甲基巯基丙酸辅酶A和甲硫基丙烯酰辅酶A,之后转化为甲硫醇,并进入甲基裂解途径最终产生甲烷[10]。然而,最后一步的甲基还原过程仍需要对氧敏感的Mcr酶参与,对此,Damm等[7]通过理论模型研究发现,细菌可通过代谢活动维持细胞内的厌氧环境,但此结果仍缺乏验证的实验证据。在C-P键裂解途径中,微生物首先分解MPn,并依次转化为α-D-核糖-1-MPn-5-三磷酸和 α-D-核糖-1-MPn-5-磷酸,最终转化为甲烷[10]。C-P键裂解酶对氧气并不敏感,且此酶的Phn的操纵子广泛存在于整个细菌结构域中[75],因此,此代谢过程及其普遍,可由多种细菌介导。此外,由于传统产甲烷途径中的大部分基因都可在细菌中发现,仅仅缺少部分关键酶,Tang等[10]和 Bižić-Ionescu等[95]推测存在另一种新的产甲烷途径,此途径可能包括一个去甲基化途径,一个已知或新的C1-载体(如四氢叶酸或四氢甲烷),一个甲基转移酶,一个甲基辅酶M类似物(如巯基丙酸)和一个甲基还原酶,这个过程最有可能直接作用于C1载体,但此产甲烷途径是否存在仍需要大量实验证据验证。

图2 真光层海水甲烷产生可能存在的代谢途径(据文献[95,10,106]修改)虚线蓝框表示传统产甲烷途径,包括CO2还原途径、甲基裂解途径和乙酸发酵途径;虚线红框表示替代的新产甲烷途径,包括C-S键裂解途径,C-P键裂解途径,以及以R-CH3为底物的假设途径(红色虚线箭头)。每种代谢途径的底物用红色字体标出。Fig.2 Possible methanogenesis metabolic pathways in ocean’s euphotic zone (modified from references [95,10,106])The dash blue box indicates the classical methanogenic pathways mediated by methanogenic archaea, including the CO2 reduction pathway, the methylation pathway and the acetate fermentation pathway; the dash red box indicates bacterial-mediated alternative new pathways, including cleavage pathway of the C-S bond, cleavage pathway of the C-P bond and hypothetical pathway with R-CH3 as a substrate (dash red arrows).The substrates for each metabolic pathway are indicated in red.
3 结论与展望
真光层海水中普遍存在甲烷过饱和现象,由于接近大气,真光层海水甲烷可能是引起大气甲烷含量变化的直接因素,开展真光层甲烷的来源研究对评估甲烷及气候的影响具有重要意义。综合研究结果表明,真光层海水过饱和甲烷可能存在以下来源:①在离海岸较近、深度较浅的海域,真光层海水过饱和甲烷可能来源于沉积物、冷泉或邻近河流;②对于远洋海域,甲烷可能产生于原位生物。产甲烷古菌和细菌可能是真光层甲烷产生的执行者,它们利用藻类提供的H2、甲基化合物等为底物参与甲烷代谢;藻类可能是真光层海水甲烷产生的参与者,除了能为产甲烷古菌、细菌提供底物,自身也可能直接产生甲烷;③非生物作用也会产生甲烷,但其贡献可能极低。此外,真光层海水过饱和甲烷的形成受区域、季节及营养盐变化的影响,同时受甲烷氧化和扩散作用调节。远洋海域真光层过饱和甲烷可能由原位微生物主导,由于常规产甲烷过程需要严格的厌氧条件,因而真光层海水甲烷产生的代谢机制有其特殊性,目前推测甲烷产生①可能由产甲烷古菌介导,它们利用常规的产甲烷途径,存在于海水微厌氧环境,或自身形成抵抗氧气影响的能力;②也可能由细菌介导,它们避开传统的对氧敏感途径,转而使用替代的C-S键裂解途径或C-P键裂解途径产生甲烷。
即使真光层原位微生物能够介导甲烷产生,但这些微生物的代谢过程以及对真光层海水甲烷的贡献程度有待确定:①在产甲烷古菌介导的途径中,由于海水古菌的丰度非常低,产甲烷古菌是否能够维持真光层甲烷过饱和状态?②在以降解DMSP为代表的C-S键裂解途径中,细菌如何确保甲基辅酶M还原酶不受氧气影响?③在以降解MPn为代表的C-P键裂解途径中,海洋中是否有大量MPn底物来源?虽然细菌也可以Phn为底物,但未必Phn的降解都会导致甲烷产生,其对真光层甲烷的贡献量是多少?此外,这些产甲烷路径的详细代谢步骤仍需进一步研究,稳定同位素示踪和微生物纯培养等技术是研究代谢通路的重要手段;明确过饱和甲烷形成的条件与规律,全方位探究真光层海水甲烷参与的生物地球化学循环,是评估真光层海水甲烷对全球气候变化影响的重要条件。
猜你喜欢
分子催化(2022年1期)2022-11-02
化学工业与工程(2022年1期)2022-03-29
化工环保(2021年5期)2021-10-19
化学与生物工程(2021年8期)2021-08-26
军民两用技术与产品(2021年10期)2021-03-16
环境卫生工程(2020年6期)2020-12-30
江苏理工学院学报(2020年2期)2020-10-23
水上消防(2020年1期)2020-07-24
中成药(2019年12期)2020-01-04
疯狂英语·新读写(2018年3期)2018-11-29