
不同育肥阶段荷斯坦阉牛亚急性瘤胃酸中毒(SARA)发病情况及相关指标分析
2020-02-24 08:21:22王建发赵晓川
中国兽医学报 2020年12期
卜 也,王建发,孙 芳,连 帅,赵晓川,武 瑞*
(1.黑龙江八一农垦大学 动物科技学院,黑龙江 大庆 163319;2.黑龙江省农业科学院畜牧研究所,黑龙江 哈尔滨 150086)
我国牛肉进口量逐年增长,2019年上半年牛肉进口量56.04万吨,同比增51.2%,出口量69.7吨,同比减60.2%,国内市场呈供不应求现象。荷斯坦品种牛具有奶公犊生长速度快,成年牛体型大,活牛价格低并且种源丰富的育肥优势,但其出肉率较纯种肉牛低,为提高其生产性能,在实际生产中不可避免地使用高精料育肥。日粮营养成分搭配不当,饲养管理不佳等问题,会导致瘤胃功能降低,极易诱发亚急性瘤胃酸中毒(SARA),影响育肥牛的健康,降低其增重水平,直接增加了养殖成本。(前人的研究进展)育肥牛采食大量精饲料,通过瘤胃微生物发酵作用产生过量的挥发性脂肪酸(VFA)[1]和乳酸(LA)[2]等代谢产物继而诱发SARA,导致瘤胃菌群结构异常[3],使瘤胃液中VFA等代谢产物浓度持续升高。瘤胃液pH值长期处于较低水平是继发非创伤性瘤胃炎的直接因素,其原因为发生SARA后,刺激瘤胃上皮结构改变,长乳头受损缺失,以及细胞受损后溃疡增生形成局部过度角质化[4-6],且高浓度的LA会引起真菌性瘤胃炎[7]。反刍动物发生瘤胃炎后,瘤胃内容物中致病性细菌和致炎物质穿过受损的瘤胃黏膜屏障进入血液循环系统,经体循环到达肝脏。化脓性细菌在肝脏内大量繁殖,毒素的产生超过肝脏清除能力,最终导致肝脓肿,也称瘤胃性肝脓肿[8-9,4]。另外,有研究表明肝脓肿会影响机体的脂肪代谢[10],瘤胃发酵产物多数为能量代谢基底物,通过瘤胃吸收入血,经肝脏转化为甘油三酯(TG)和胆固醇(TC)等脂肪合成前体物在肝脏中蓄积[11-12],导致脂肪肝。不同育肥阶段的SARA发病情况、代谢特征有差异。荷斯坦阉牛在育肥过程中所诱发的SARA对动物机体代谢的影响尚不完全明确。本试验旨在研究荷斯坦阉牛育肥过程中SARA的发病情况及相关指标的变化,为SARA的预防保健及早期诊断提供有效依据。
1 材料与方法
1.1 试验动物本试验选用黑龙江省某肉牛育肥场荷斯坦阉牛为研究对象,在同批断奶荷斯坦阉牛中随机选择30头为试验样本。该场饲养方式为舍饲散栏饲养,自拌料于每天7:00和17:00投料,自由饮水,各阶段换料过渡期3 d。经过育肥前期(6~12月龄)、育肥中期(12~18月龄)和育肥后期(18~24月龄)3个阶段的试验期。该牛场各阶段育肥牛饲粮组成及营养水平见表1(营养水平为实测值)。
1.2 样品采集试验期内,育肥前期、中期、后期分别采样3次,采样节点平均分布于各育肥阶段。
清晨给料前测体质量并保定育肥牛,尾静脉采集非抗凝血10 mL,4℃保存运输,实验室使用离心机3 000 r/min 离心5 min取血清,-80℃冰箱储存用于生化检测;使用瘤胃管于进食后4 h采集瘤胃液。
1.3 SARA指标便捷式pH计直接测定瘤胃液pH值。目前,该病诊断的主要依据是瘤胃液pH值,当为瘤胃液pH值处于5.0~5.8,且数值每天在这个区间内超过3 h,持续数天,可定义为SARA[9,13]。

表1 基础饲粮组成及营养水平(干物质基础) %
1.4 肝功指标血清谷氨酰转肽酶(GGT,U/L,IFCC法)、血清丙氨酸氨基转移酶(ALT,U/L,速率法)、血清天门冬氨酸转移酶(AST,U/L,速率法)、血清碱性磷酸酶测定(ALP,U/L,酶联免疫法);总蛋白(TP,g/L,比色法)、白蛋白(ALB,g/L,比色法)、球蛋白(GLO,g/L,比色法)。以上指标采用全自动生化分析仪按照试剂盒操作说明进行。
1.5 能量代谢指标血清甘油三酯浓度测定(TG,mmol/L,GPO-PAP法)、血清胆固醇浓度测定(TC,mmol/L,氧化酶法)。以上指标采用全自动生化分析仪按照试剂盒操作说明进行。
1.6 数据分析所得数据利用Excel软件整理,使用SPSS 20统计软件的ANOVA程序进行分析,用于评估每组中平均参数值之间差异的显著性的概率。数据分析所得结果中P<0.05表示差异显著,P<0.01表示差异极显著,在表格中分别用小写字母(abc)和大写字母(ABC)表示。
2 结果
2.1 SARA发病情况由表2可见,育肥前期健康牛比例为30%,轻度SARA发病率为50%,中度SARA发病率20%;育肥中期健康牛比例为20%,轻度和中度SARA发病率均为40%;育肥后期荷斯坦阉牛轻度SARA和中度SARA发病率分别为20%和80%。
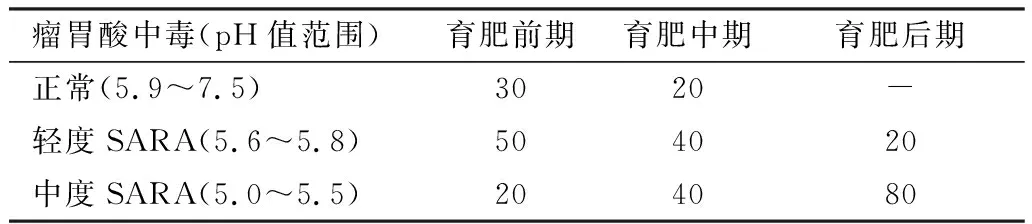
表2 3个育肥阶段瘤胃酸中毒发病情况 %
2.2 肝脏功能指标由表3可见,育肥后期荷斯坦阉牛肝功GGT活性极显著高于育肥前期和中期(P<0.01),但均在正常区间内11.00~66.00 U/L[14];3个育肥阶段荷斯坦阉牛肝功ALT活性组间差异极显著(P<0.01),且育肥中期和后期荷斯坦阉牛肝功ALT活性高于肉牛正常值40.53 U/L[15];3个育肥阶段荷斯坦阉牛肝功AST活性均高于正常值53.67 U/L[16],且育肥后期显著高于前期和中期(P<0.05);育肥后期荷斯坦阉牛肝功ALP活性极显著低于育肥前期(P<0.01),显著低于育肥后期(P<0.05),但均在正常区间内150.53~482.08 U/L[15]。

表3 不同育肥阶段荷斯坦阉牛肝功酶指标检测结果 U/L
由表4可见,3个阶段荷斯坦阉牛血清TP含量组间无显著性差异(P>0.05);育肥前期荷斯坦阉牛血清ALB含量显著高于中期和后期(P<0.05),且高于正常值40.20 U/L[17];育肥后期荷斯坦阉牛血清GLO含量极显著高于育肥前期和中期(P<0.01),且高于正常值41.66 U/L[18]。
2.3 能量代谢指标由表5可见,3个阶段荷斯坦阉牛血清TG含量均高于正常值0.23 mmol/L[19],其中育肥后期极显著高于中期(P<0.01),显著高于前期(P<0.05),中期显著高于前期(P<0.05);育肥后期荷斯坦阉牛血清TC含量高于正常值3.23 mmol/L[15],且极显著高于育肥前期和中期(P<0.01)。
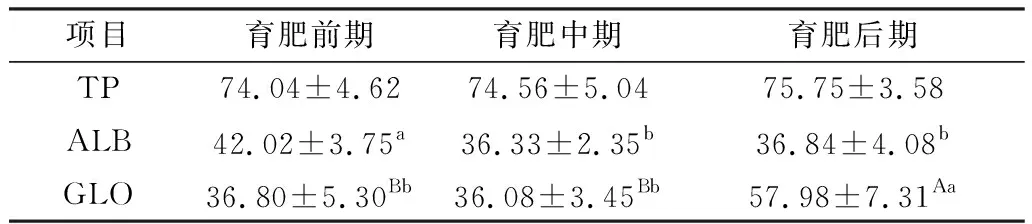
表4 不同育肥阶段荷斯坦阉牛血清蛋白含量检测结果 U/L

表5 不同育肥阶段荷斯坦阉牛血清中TG和TC含量 mmol/L
2.4 生长情况由表3可见,随着育肥期延长,3个阶段荷斯坦阉牛平均日增重(ADG)呈下降趋势,分别为1.68,0.75,0.63 kg/d。

表3 不同育肥阶段荷斯坦阉牛增重水平 kg
3 讨论
3.1 牛场SARA发病情况SARA是肉牛场常见的代谢病之一,PLAIZIER[9]和ENEMARK等[20]确定SARA的诊断标准主要依据瘤胃液pH值的范围。根据本试验所得数据可以看出,3个阶段荷斯坦阉牛均患有不同程度的SARA。育肥前期荷斯坦阉牛为断奶犊牛,通常使用高蛋白、高脂肪的精补料作为早期断奶的母乳替代品,采食开食料初期,瘤胃对颗粒料饲料消化能力较弱,容易造成积食,未消化完全的饲料在瘤胃内逐渐发酵产酸,使育肥前期犊牛瘤胃液pH值低于正常范围[21]。该牛场育肥前期荷斯坦阉牛多数处于轻度SARA状态,此时病情轻,随着犊牛瘤胃的发育,可逐渐转归。育肥中期的牛多数为架子牛,除少部分为本场自繁外,多数为购自周边农户的荷斯坦阉牛。进场育肥前此阶段的牛多数以草料饲养,膘情较瘦,经过不同距离的运输,进场后精饲料改饲过渡不完全或方法不当,因此育肥中期SARA发病情况较育肥前期严重。育肥后期荷斯坦阉牛采食量增大,高能饲料长期摄入,超过育肥后期荷斯坦阉牛吸收转化速率,使患病率达到100%。
荷斯坦阉牛长期采食高精料,经淀粉分解菌发酵作用使丙酸酶活性逐渐升高,随着育肥期增长,超过瘤胃酸碱代偿能力,使瘤胃液pH值持续降低,诱发SARA。瘤胃内酸碱度耐受能力不同的微生物因其外环境改变而呈暴发增殖或失活状态[22],导致SARA异常代谢产物经瘤胃壁毛细血管和损伤的瘤胃上皮组织扩散至肝脏及机体其他器官、组织,引发机体代谢紊乱,甚至更严重的疾病,危害牛群健康[23]。
3.2 SARA对荷斯坦阉牛肝功的影响肝脏是机体重要的物质合成和转化器官,也是机体营养物质储存和毒素代谢场所。GGT、ALT、AST和ALP是肝脏活动的主要酶系,也是临床上评价肝功的关键指标[24]。GGT是肝胆疾病的预警指标,主要存在于肝胆管上皮和肝细胞浆中,当机体肝胆管阻塞或肝内梗阻时,GGT随胆汁返流,导致血清GGT活性升高,因此血清GGT活性升高预示动物存在患急性或慢性肝炎、肝硬化及脂肪肝等肝胆疾病的风险[25]。多种肝脏疾病均可导致ALT和AST在血清中活性升高,因此这两种酶偏高是反映肝细胞损伤的主要指标[26]。ALP的确切生理作用尚不明确,一般认为其与骨骼的钙化和机体生长密切相关,当动物患阻塞性黄疸、肝脓肿、肝硬化坏死以及脂肪肝等疾病时,肝脏异常增生,肝细胞过度产生ALP,返流入血,导致血清ALP活性升高,但不能确认某种疾病[27]。本试验中,随着育肥期的增加,3个阶段荷斯坦阉牛血清GGT活性虽在正常范围内,但育肥后期荷斯坦阉牛血清GGT活性明显增高,提示存在患肝胆疾病的风险。3个阶段荷斯坦阉牛血清中ALT和AST活性整体增加,提示肝细胞损伤程度可能加大。3个阶段荷斯坦阉牛血清ALP活性逐渐下降,可能是因为育肥后期荷斯坦阉牛骨骼已基本钙化完全,育肥前期和中期荷斯坦阉牛骨骼生长和钙化速度高于育肥后期,所以育肥后期荷斯坦阉牛血清ALP活性低于前两个阶段。根据以上分析可以看出,育肥后期的荷斯坦阉牛患肝胆疾病的风险最高,肝细胞损伤程度相对较重。
TP是ALB和GLO的合称,主要由肝脏生成进入血液循环,具有维持血液渗透压、pH值、营养、调节代谢及免疫功能,是临床血清生化检查的关键指标,ALB反映肝脏合成能力,GLO具有免疫作用,是机体的主要免疫蛋白[28]。本试验3个阶段荷斯坦阉牛TP含量较稳定,ALB含量依次降低,GLO含量升高明显[29],ALB/GLO值减小[30],说明随着育肥期的延长,荷斯坦阉牛摄入的蛋白和能量虽增加,但营养物质的转化率降低,造成其在机体蓄积,使荷斯坦阉牛的肝脏负担逐渐加重,有患脂肪肝的风险[31],甚至肝硬化,同时育肥后期荷斯坦阉牛血清GLO含量偏高,说明机体可能存在炎症反应[32],严重时将影响荷斯坦阉牛的健康。
3.3 SARA对荷斯坦阉牛能量代谢的影响TG是机体主要的储能物质,主要来源分为外源性和内源性,外源性由食物中脂肪在肠道经消化酶作用下,在肠黏膜上皮细胞内合成,内源性主要是由肝脏合成其次是脂肪组织,根据情况分解供能[33]。TC是类脂的主要成分,存在形式包括高密度脂蛋白胆固醇、低密度脂蛋白胆固醇、极低密度脂蛋白胆固醇和乳糜微粒,参与机体的营养代谢调节和生物膜的组成[34]。本试验育肥后期荷斯坦阉牛血清TG、TC含量整体高于其他阶段,且高于正常水平,说明育肥后期可能患有高血脂症,甚至脂肪肝,屠宰可见肝脏、心脏和肾脏有大量脂肪沉积,增加脏器负担[35]。另外,育肥后期荷斯坦阉牛等量全血分离血清相对较少,呈现乳糜血现象[36],这与荷斯坦阉牛育肥后期摄入大量的高能量精料有关。
3.4 SARA对荷斯坦阉牛增重的影响本试验育肥前期、中期、后期荷斯坦阉牛所用饲粮综合净能(NEmf)分别为10.2,11.5,12.3 MJ/kg,平均日增重(ADG)分别为1.68,0.75,0.63 kg/d,根据《肉牛营养需要》所给出的不同育肥阶段荷斯坦阉牛所需NEmf和ADG标准可知,育肥前期和中期荷斯坦阉牛所用饲料实测NEmf均高于实际需要[37],料重比过高,增加饲养成本。
综上所述,通过试验数据可推测,荷斯坦阉牛育肥过程中饲粮能量水平过高会诱发SARA,同时导致肝功和能量代谢指标异常,且随育肥期延长而加重,影响荷斯坦阉牛育肥性能。该牛场饲粮配方不适于荷斯坦阉牛育肥,且换料过渡时间短可能是造成SARA高发的原因。
猜你喜欢
今日畜牧兽医(2022年10期)2022-12-23 06:23:18
中国饲料(2022年5期)2022-04-26 13:42:34
医学信息(2017年23期)2017-12-05 13:44:16
兽医导刊(2016年12期)2016-05-17 03:51:35
兽医导刊(2016年12期)2016-05-17 03:51:31
华南农业大学学报(2015年5期)2015-12-04 03:04:38
今日健康(2015年8期)2015-10-17 10:50:10
畜牧兽医学报(2015年3期)2015-07-05 08:22:16
中国当代医药(2015年21期)2015-03-01 02:05:01
云南畜牧兽医(2014年2期)2014-02-28 21:25:17
