
缺氧诱导通路基因多态性与肺癌及临床特征的关联性研究
2020-02-14 10:30于伟玲谢宗宙黄琰菁
安徽医科大学学报 2020年1期
于伟玲,肖 莎,谢宗宙,黄琰菁,陈 元
肺癌是世界范围内死亡率最高的恶性肿瘤之一。在全球范围内,2012年诊断出约180万新发肺癌病例,占世界癌症总发病率的12.9%,同年其死亡159万人,占癌症死亡总数的19.4%[1]。目前的流行病研究结果显示中国过去10年肺癌发病率和死亡率呈持续上升趋势,平均每年以1.6%的比例上升[2]。
肺癌细胞的形成和转移与缺氧微环境适应和生长密切相关[3-4],现有研究发现缺氧诱导因子相关通路分子异常与肺癌进展有重要联系[5],其遗传多态性与肺癌发生密切相关[6-7]。近期日本人群研究发现缺氧诱导因子rs13419896 (Hypoxia-inducible factor, HIF2A)与血管内皮生长因子A rs833061-AA (vascular endothelial growth factor A, VEGFA)基因的多态性与肺癌发生风险相关[8-9]。由于亚太地区人在生活环境和人群遗传背景具有较高的相似性,而上述结果在中国地区的目前无相关证实,该研究拟探讨上述肺癌易感位点与我国肺癌发生的相关性。
1 材料与方法
1.1 病例资料样本来源海南汉族人群,共纳入肺癌患者438例,为2011年3月~2017年7月间,海口市人民医院和海南省人民医院病理检查确诊的肺癌患者,456例对照样本来源于同时期海南地区健康体检中心常规体检人群,无呼吸系统疾病及肺癌家族史。
1.2 流行病学调查制定统一流行病学调查量表,内容包括:一般人口学特征、吸烟、饮酒以及既往糖尿病史,排除呼吸系统家族史等。吸烟者定义为不吸烟者、既往吸烟者和目前正在吸烟者,饮酒者同样定义为不饮酒者、既往饮酒者和正在饮酒者,并收集临床病理分型和临床肺癌分期。临床数据整理和分析,按照第8版肺癌TNM分期标准。
1.3 样本采集在获取知情同意,并签署知情同意书后,分别采集肺癌患者和对照组外周静脉血3 ml,EDTA抗凝,-20 ℃冰箱保存。
1.4 DNA提取采用外周血DNA提取试剂盒(Axygen Biosciences公司)提取人全基因组DNA,提取步骤按操作说明书进行,提取的DNA保存于-60 ℃冰箱。
1.5 基因分型rs13419896和rs833061基因分型应用Taqman genotyping assays(applied biosystems, ABI),HIF2A, rs13419896, C__31180907_10和VEGFA, rs833061, C__1647381_10基因分型的探针,反应的条件如下2倍Taqman genotyping master mix(ABI) 2.5 μl, 20 SNP genotyping assay mix(ABI) 0.25 μl。模板DNA 2.25 μl。 PCR反应条件95 ℃、10 min, 95 ℃变性15 s, 60 ℃延长90 s,共40个循环, 完成后读取荧光信号完成基因分型。
1.6 统计学处理采用STATA 10.0SE软件对数据进行统计学分析。配对t检验比较病例组和对照组间性别、年龄、饮酒及吸烟情况,Hardy-Weinberg平衡检验检验对照组样本是否来源于同一个群体;χ2检验比较两组间高危因素情况及不同基因型在两组间的频率分布, 应用Logistic多因素回归分析计算OR和95%CI以估计不同基因型与肺癌风险之间的相关性,以P<0.05为差异有统计学意义。
2 结果
2.1 临床特点肺癌病例组平均年龄为29~87(55.8±15.1)岁;对照组平均年龄为30~72(48.1±8.07)岁,年龄上肺癌病例组年龄较健康对照组大(P<0.001),而肺癌病例组与对照组之间性别无明显差异(P=0.96)。通过吸烟和饮酒行分类分析,不同肺癌病例组吸烟个体和持续饮酒个体与对照组相比,差异有统计学意义,具体见表1。

表1 病例组和对照组临床特点[n(%)]
2.2 Hardy-Weinberg遗传平衡检验rs13419896和rs833061成功分型为98.2%和99.0%。通过Hardy-Weinberg检验分析对照组是否来源于同一个整体,rs13419896(χ2=1.88,P=0.17)、rs833061(χ2=0.38,P=0.532),差异无统计学意义,证明样本的收集来源一个总体。
2.3 HIF2A基因rs13419896和VEGFA rs833061多态性位点与肺癌关联分析分析结果显示携带rs13419896 A等位基因对比G等位基因具有降低肺癌发病风险[OR=0.72, 95%CI(0.58~0.88),P=0.001 1],通过对基因型的加性模型、隐性模型和显性模型遗传模型进行分析,发现在加性模型和显性模型中差异有统计学意义,见表2。进一步分析rs13419896基因型与肺癌中病理类型(χ2= 3.27,P=0.77)和临床分期(χ2=8.88,P=0.18)无相关性。而rs833061位点的等位基因C及T在肺癌组和对照组的分布情况分析,无统计学差异;对rs833061基因型进行三种不同遗传模型分析,亦无统计学差异,见表2。
2.4 HIF2A基因rs13419896基因型与肺癌发病相关通过对比rs13419896基因型、年龄、吸烟、饮酒、2型糖尿病等在条件回归模型中的影响,结果显示rs13419896基因型、吸烟、饮酒和年龄与肺癌相关,见表3。通过多因素回归分析,调整吸烟、饮酒和年龄的影响,rs13419896基因型仍可降低肺癌发生率,而回归模型中2型糖尿病和性别与肺癌的发病率无明显关联,见表3,回归分析中wald检验值为156.13。
2.5 肺癌高危因素肺癌分层分析进一步对肺癌高危因素进行分层分析,将以前有吸烟史和现在仍吸烟的定义为吸烟者,同样方法定义饮酒者。分析发现对于不吸烟者,rs13419896 AA基因型具有的保护作用,而在吸烟人群中保护作用不明显;rs13419896 AA基因型对饮酒者和不饮酒者均有降低肺癌的发生率;对于年龄大于50岁的患者,rs13419896 AA基因型对降低肺癌的发生率较为明显,见表4。
3 讨论
本研究通过对缺氧通路的多态性位点进行验证,发现在中国海南地区HIF2A基因rs13419896多态性位点与肺癌的发生具有相关性,rs13419896位点AA基因型降低肺癌发生,而同为该位点的等位基因GG型增加肺癌的发病风险,但未发现该阳性位点与临床分期和病理类型的关系。
通过对环境因素的分析表明,除吸烟外,饮酒和年龄也是中国海南地区肺癌发生的重要因素。同时对肺癌高危因素进行分层分析表明,对于吸烟患者其rs13419896-AA基因型保护作用不明显,而对于不吸烟患者保护作用较为明显,提示吸烟可能降低了rs13419896-AA基因型保护作用,rs13419896-AA保护作用与吸烟与否有密切相关。无论是饮酒患者或者不饮酒肺癌患者rs13419896-AA基因型性均有保护作用,可能的机制是饮酒不影响rs13419896-AA对肺癌发生保护机制,或者饮酒通过其他的通路致肺癌发生,本研究的结果显示饮酒本身仍是增加肺癌发生率的高危因素。
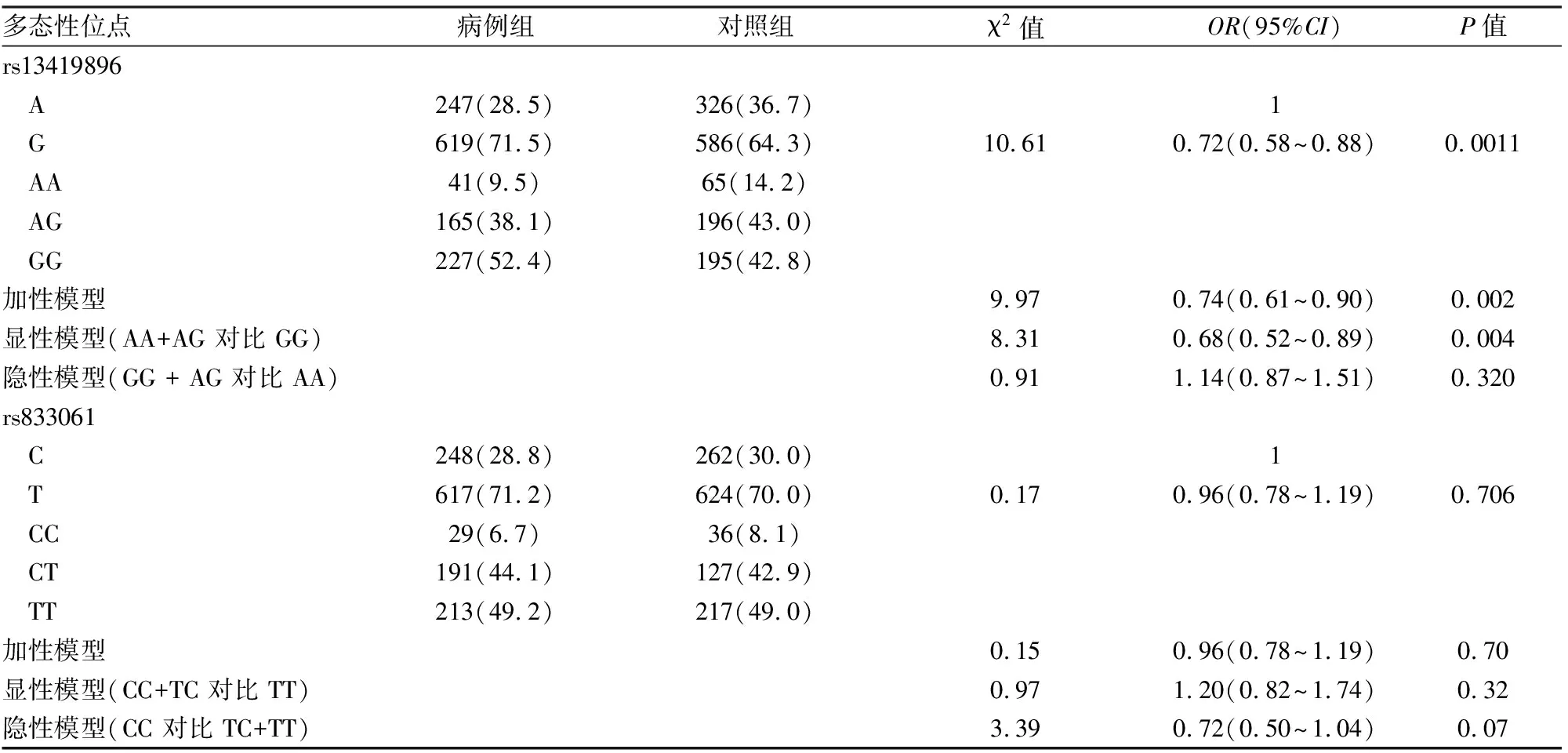
表2 rs13419896 和rs833061与肺癌的关联性[n(%)]
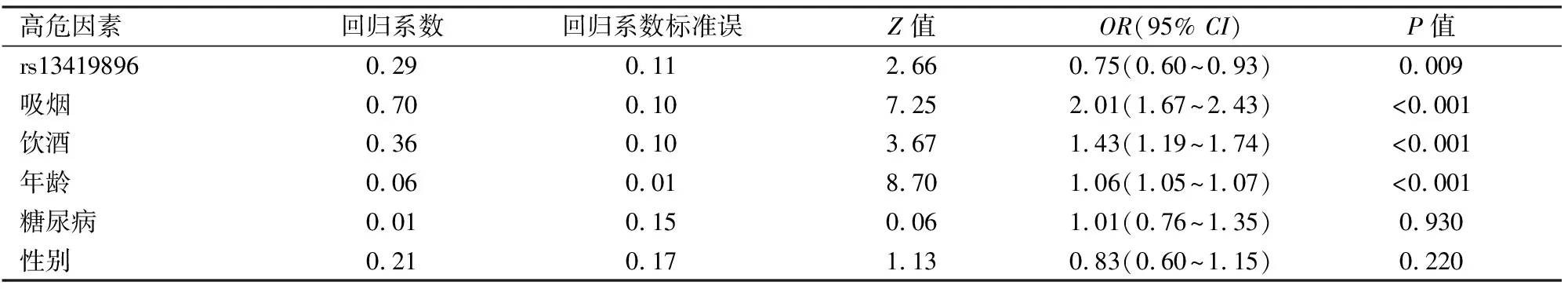
表3 多因素Logistic回归分析分析肺癌的高危因素

表4 rs13419896在吸烟、饮酒和年龄在肺癌组和健康对照组分层分析
本研究结果与Yamamoto et al[8]对日本九州地区肺癌患者的研究结果相同,rs13419896-AA可以显著降低肺癌的发生率,而未发现上述研究中VEGFA基因rs833061与肺癌的相关性,可能与该多态性位在中国人群(C/T:0.279/0.721)和日本人群(C/T:0.343/0.656)rs833061等位基因频率差异有关。不同于Kim et al[10]在广岛地区的研究结果,该研究对76例肺癌患者基因型分析,提示rs13419896-AA基因型增加肺癌发生率,并且对基因型的生物学功能进行了深入的验证,发现其有一定的相关性。该易感性结果的差异可能来源于研究中肺癌病例样本收集较少,或者样本来源地区的基因频率的差异,因而对在该地区进行独立大样本的验证,显得尤为必要。同时该研究对多态性位点的生物学功能进行分析发现rs13419896 A基因型相对于G基因型在A549、PC-9和 HSC-2细胞内可以增加转录活性,rs13419896-AA基因型与肺癌的临床进展和预后有关。虽然认为肺癌HIF2A组织高表达与肿瘤的进展和预后不良相关[10-11]。但有也研究指出HIF家族基因通过调节不同靶基因的转录活性,发挥促瘤与抗肿瘤的双重作用[12-13]。因而该上述结果仍然需要进一步独立的功能分析来证实,同时联系其临床特征来进一步完善对HIF2A在肺癌发生的认识。
少部分遗传易感性研究出现结果差异,这些差异主要来源于人群种族和种族内人群分层[14],或者独立研究本身的限制,这种情况使得跨区域的重复验证性研究具有重要的应用意义。虽然本研究分析了海南地区438例肺癌病人样本和456例健康对照人样本中rs13419896与肺癌的关系,但研究结果来源单一中心,仍需要多中心的进一步验证,明确rs13419896与肺癌的关系。验证后的结果将为精准医疗和个体化预防,提供一定的理论基础和临床参考。同时rs13419896位于HIF2A的转录调控区,为后续进一步的功能验证提供便利,也为探索HIF2A基因SNP位点在缺氧条件下,研究调节下游靶基因的机制提供了可能。
猜你喜欢
传染病信息(2022年4期)2022-11-23
西部医学(2022年9期)2022-09-26
世界科学技术-中医药现代化(2022年3期)2022-08-22
昆明医科大学学报(2022年3期)2022-04-19
昆明医科大学学报(2022年2期)2022-03-29
昆明医科大学学报(2021年2期)2021-03-29
生物学教学(2018年8期)2018-09-03
中华诗词(2017年5期)2017-11-08
作文世界(小学版)(2017年6期)2017-08-25
Coco薇(2016年8期)2016-10-09
