
水分胁迫对日光温室葡萄叶片生理变化及果实发育的影响*
2020-02-13 10:26牛最荣王旺田
中国生态农业学报(中英文) 2020年2期
牛最荣, 赵 霞, 张 芮**, 王旺田, 温 文
水分胁迫对日光温室葡萄叶片生理变化及果实发育的影响*
牛最荣1, 赵 霞1, 张 芮1**, 王旺田2, 温 文1
(1. 甘肃农业大学水利水电工程学院 兰州 730070; 2. 甘肃农业大学生命科学技术学院 兰州 730070)
为探究鲜食日光温室葡萄高效节水生产的水分管理方式, 选取日光温室6 a生葡萄‘红地球’为试验材料, 以充分灌溉为对照[CK, 土壤含水率为75%~100%的田间持水率(f)], 分别在萌芽期、新梢生长期、开花坐果期、果实膨大期、着色成熟期设置土壤含水率为55%f(其他4个生育期为75%~100%f)的5个水分胁迫处理, 调查叶片超氧化物歧化酶(SOD)活性及膜质过氧化物丙二醛(MDA)和脯氨酸(Pro)含量、果实纵横径、果实硬度、单粒重量和产量, 研究日光温室葡萄叶片生理和果实产量对水分胁迫的响应机制。结果表明, 在果实膨大期、着色成熟期施加水分胁迫均会显著降低葡萄叶片SOD活性和Pro含量, 导致MDA含量大量积累, 对叶片产生不利影响。在萌芽期、新梢生长期、开花坐果期进行水分胁迫对葡萄叶片SOD活性、Pro含量影响不大, 但对叶片MDA含量的影响具有滞后效应。果实膨大期水分胁迫葡萄产量仅为14 830 kg·hm–2, 较CK和萌芽期、新梢生长期水分胁迫显著减产20%以上。因此, 水分胁迫显著降低果实膨大期SOD活性及渗透调节物质含量, 增加膜质过氧化物含量, 并导致产量降低, 该时期不宜进行亏水处理; 生长前期的水分胁迫对葡萄叶片的生理和果实的影响不显著。
葡萄; 水分胁迫; 生育期; 叶片生理; 果实发育; 日光温室
葡萄(L.)是中国北方地区重要的林果树种, 葡萄产量和品质对农民收入有重要影响[1]。甘肃省河西走廊, 气候干燥、光照充足、昼夜温差大, 是发展优质高产葡萄生产的理想区域之一[2-3]; 目前, 葡萄已成为甘肃武威、张掖、酒泉等地农村发展和农民增收致富的支柱产业。近年来, 利用日光温室对鲜食葡萄进行春季避光、冬季增温的栽培方式实现了葡萄的冬季新鲜上市供应, 提高了葡萄价格和农民的种植收入。葡萄是一种对水分需求较大的经济作物, 而目前我国干旱缺水形势是影响农业生产和农民增收的重要的瓶颈问题[4-5]。实现葡萄节水高效生产是解决甘肃河西走廊发展葡萄种植产业与水资源先天不足矛盾的关键所在。
目前, 对日光温室葡萄的研究主要集中在灌溉制度、耗水规律、水分胁迫对果实生长、产量和品质的影响。如殷常青等[6]根据日光温室葡萄在不同生育期的耗水规律, 筛选出温室葡萄最优滴灌制度; 张芮等[7-8]的研究表明, 萌芽期中度水分胁迫可提高温室葡萄产量和水分利用效率, 而抽蔓后期至果实膨大初期的中度水分胁迫对果实膨大速率存在明显的复水补偿效应。浆果膨大—聚糖着色期亏水处理可降低温室葡萄可滴定酸, 提高花青素及可溶性固形物[9]。以上研究为发展日光温室葡萄节水优质高效提供了依据。然而不同水分胁迫下, 日光温室葡萄生理指标如何?水分胁迫与葡萄叶片抗逆性互馈机理如何?目前关于这方面的内容鲜有报道。水分胁迫对植物的生长发育、生理和产量造成极大的影响。在正常水分条件下, 植物细胞内的有害氧自由基的产生和清除处于一种动态平衡[10]。当植物受到水分胁迫时, 这种平衡遭到破坏, 产生大量对细胞有毒害作用的氧自由基, 导致膜质过氧化物丙二醛(MDA)含量增加[11]。同时, 植物体内的渗透调节物质(Pro)大量积累, 其含量在一定程度上可以反映出植物体内水分状况。而核心酶超氧化物歧化酶(SOD)能有效清除氧自由基, 保护细胞免受活性氧自由基的毒害。为此, 本研究在葡萄不同生育期设置水分胁迫, 并测定葡萄各生育期叶片超氧化物歧化酶(SOD)、膜质过氧化物丙二醛(MDA)、渗透调节物质脯氨酸(Pro)和葡萄果实生长等指标, 旨在探讨上述指标对各生育期土壤水分胁迫的生理响应机理, 以期为鲜食日光温室葡萄高效节水生产提供科学依据。
1 材料与方法
1.1 研究区概况
试验于2014—2016年在甘肃省河西走廊东端张掖市灌溉试验基地(100°26′E, 38°56′N)的日光温室进行。该地海拔1 482.7 m, 多年平均降水量125 mm,多年平均蒸发量2 047.9 mm。供试土壤主要为中壤土, pH 7.8, 密度1.47 g∙cm–3, 田间持水率22.8%(体积分数), 土壤有机质13.7 g∙kg–1, 碱解氮32.04 mg∙kg–1, 速效磷27.8 mg∙kg–1, 速效钾1 137.4 mg∙kg–1。
1.2 研究对象
试验对象为6 a生葡萄‘红地球’(red globe), 采用单壁篱架栽培。葡萄栽培设施采用日光温室, 单棚建筑面积为8 m×80 m。随机布设试验小区, 小区长宽为8 m×4 m, 每小区两行葡萄。灌溉方式采用滴灌, 1管1行控制模式。贴片式滴灌带, 滴头流量为3 L∙h–1, 间距50 cm。
1.3 研究方法
根据已有研究[5]将葡萄生育期划分为萌芽期(5月6―29日, 14 d)、新梢生长期(5月30日―6月18日, 20 d)、开花坐果期(6月19―29日, 11 d)、果实膨大期(6月30日―9月9日, 72 d)和着色成熟期(9月10日―12月15日, 97 d)。为探讨葡萄对不同生育期水分亏缺的生理响应, 以全生育期充分灌溉为对照(CK), 把5个生育期中的1个生育期土壤含水率设置为55%田间持水率(中度水分胁迫)[12], 其他4个生育期土壤含水率设置为75%的田间持水率作为1个处理(T), 共5个胁迫处理, 每个处理3个重复, 共计18个小区。各处理试验设计见表1, 当CK和T1-T5处理土壤含水率达到表1中土壤含水率时进行灌溉, CK和T1-T5土壤含水率分别达田间持水率的100%和80%时停止灌溉。土壤含水率利用烘干法测定, 土层测定深度为100 cm, 每20 cm一层, 每7 d测定一次, 萌芽前、灌水前后、土壤含水率接近灌水下限(表1)时加测。每个小区3个测点, 其中2个测点分别位于该小区一行葡萄两端的2株葡萄的中间位置, 第3个测点位于另一行葡萄的中间位置。

表1 不同水分胁迫处理葡萄不同生育期土壤水分下限(占田间持水率的百分比)
1.4 测定指标与方法
于2016年试验进行3 a后进行以下各指标的测定。
叶片生理指标: 当各处理在各生育期末期土壤含水率达到试验设计对应的下限值时, 采摘各处理小区定株测量葡萄(5株)新稍第六、七节位的叶片, 每株2~3片, 用纱布擦干, 锡箔纸包裹后立即放置于液氮罐内, 待冷冻后放置于超低温冰箱内保存。叶片丙二醛(MDA)含量及脯氨酸(Pro)含量利用邹琦[13]的方法测定, 叶片超氧化物歧化酶(SOD)活性采用高俊凤[14]和赵世杰等[15]的方法测定。
果粒纵横径: 葡萄进入成熟期后, 对各处理小区定株测量葡萄(5株)的所有果穗, 在果穗上、中、下部位顺序各选6粒果粒, 利用游标卡尺定期测定葡萄果粒纵横径, 单位mm; 葡萄采摘时, 采用GY-1果实硬度计测定果粒硬度, 单位kg∙cm–2。
产量: 葡萄成熟采摘时, 对各处理小区所有葡萄果穗现场收获, 测定产量, 根据小区实际面积推算标准产量, 单位kg∙hm–2。
1.5 数据处理
利用Excel 2013软件对试验数据进行整理, 使用SPASS 19.0软件对数据进行单因素方差分析。
2 结果与分析
2.1 水分胁迫对日光温室葡萄叶片SOD活性的影响
不同生育期水分胁迫对葡萄叶片超氧化物歧化酶(SOD)活性的影响如表2所示。SOD活性的变化趋势均随葡萄生长呈“由高变低、再升高又降低”变化规律。各处理叶片SOD活性基本在萌芽期最高, 且萌芽期各水分胁迫处理的SOD活性均小于同期CK, 但只有T4、T5处理SOD活性显著低于CK(<0.05)。进入新梢生长期, 所有处理葡萄叶片SOD活性均迅速降低, CK降幅度最大; 各水分胁迫处理的SOD活性均高于CK, 且T4、T5处理与CK处理差异显著(<0.05)。开花坐果期葡萄叶片SOD活性较前一生育期有所提高, 但T3处理不增反降, 且显著低于其他处理(<0.05), T1、T2和T5处理也显著低于CK处理。果实膨大期所有水分胁迫处理SOD活性均显著低于CK(<0.05)。着色成熟期, CK处理的SOD活性最高; T5处理SOD活性最低, 显著低于T2、T3、T4和CK处理, 较CK显著下降84.96%(<0.05)。
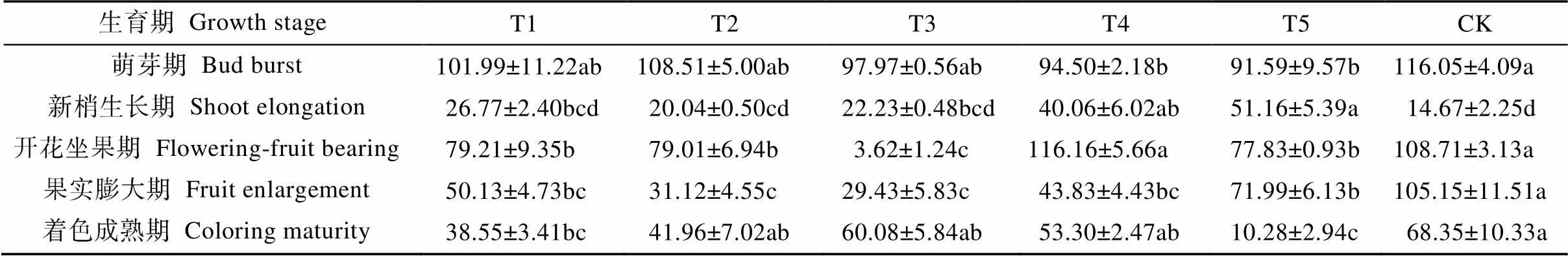
表2 不同生育期水分胁迫处理对日光温室葡萄不同生育期叶片SOD活性的影响
T1、T2、T3、T4和T5处理的水分胁迫时期分别为萌芽期、新梢生长期、开花坐果期、果实膨大期和着色成熟期。CK处理在葡萄的生育期无水分胁迫。同行不同小写字母表示处理间差异显著(<0.05)。The water stress periods of T1, T2, T3, T4 and T5 treatments are bud burst, shoot elongation, flowering-fruit bearing, fruit enlargement and coloring maturity stages, respectively. CK is the control treatment, without water stress. Different lowercases in the same line indicate significant differences among treatments at 0.05 level.
2.2 水分胁迫对葡萄叶片MDA含量的影响
表3为不同处理对葡萄叶片丙二醛(MDA)含量的影响。表3显示, 整体而言, 葡萄叶片MDA含量在新梢生长期—果实膨大期较高, 该时段是葡萄生长发育的旺盛期, 也是气温较高的时段。萌芽期、着色成熟期MDA含量明显低于另3个生长时期, 且萌芽期各处理间差异不显著。新梢生长期T1处理的MDA含量显著高于CK(<0.05), 其他处理之间无显著差异。开花坐果期, T2处理MDA含量最高, 与T1、T3、T4和CK处理呈显著性差异(<0.05)。果实膨大期T4处理MDA含量最高, 与T1、T3、T5处理差异显著(<0.05)。着色成熟期T5处理葡萄叶片MDA含量最高,与T1、T2、T3、T4处理差异显著(<0.05)。

表3 不同生育期水分胁迫处理对日光温室葡萄不同生育期叶片MDA含量的影响
T1、T2、T3、T4和T5处理的水分胁迫时期分别为萌芽期、新梢生长期、开花坐果期、果实膨大期和着色成熟期。CK处理在葡萄的生育期无水分胁迫。同行不同小写字母表示处理间差异显著(<0.05)。The water stress periods of T1, T2, T3, T4 and T5 treatments are bud burst, shoot elongation, flowering-fruit bearing, fruit enlargement and coloring maturity stages, respectively. CK is the control treatment, without water stress. Different lowercases in the same line indicate significant differences among treatments at 0.05 level.
2.3 水分胁迫对葡萄叶片Pro含量的影响
表4为不同处理对葡萄叶片脯氨酸(Pro)含量的影响。表4显示, Pro含量在葡萄整个生育期变化较为平稳。在萌芽期、开花坐果期2个短生育期(分别为14 d和11 d), 水分胁迫对Pro含量的影响并不明显。新梢生长期、果实膨大期和着色成熟期不同处理间呈现显著差异。果实膨大期, T2和T4处理Pro含量最低, 较CK分别显著减少28.97%和29.84% (<0.05), 其他处理间无显著差异。果实着色成熟期T5处理叶片Pro含量最低, 与T2处理差异显著。

表4 不同生育期水分胁迫处理对日光温室葡萄不同生育期叶片脯氨酸含量的影响
T1、T2、T3、T4和T5处理的水分胁迫时期分别为萌芽期、新梢生长期、开花坐果期、果实膨大期和着色成熟期。CK处理在葡萄的生育期无水分胁迫。同行不同小写字母表示处理间差异显著(<0.05)。The water stress periods of T1, T2, T3, T4 and T5 treatments are bud burst, shoot elongation, flowering-fruit bearing, fruit enlargement and coloring maturity stages, respectively. CK is the control treatment, without water stress. Different lowercases in the same line indicate significant differences among treatments at 0.05 level.
2.4 水分胁迫对葡萄果实的影响
表5为水分胁迫对葡萄果实性状的影响。从表5可知, 萌芽期和果实膨大期水分胁迫(T1和T4处理)果粒横径、纵径和单粒重量均显著低于CK (<0.05), T4处理又均低于T1处理。与CK相比, 水分胁迫处理对果形指数和果实硬度(除T5处理显著低于其他处理外)无显著影响。表明水分胁迫对果粒生长均有不同程度抑制作用, 其中T4处理抑制作用最显著。T4处理葡萄产量比CK显著减少19.98%, 显著低于(<0.05)萌芽期和新梢生长期水分胁迫(T1和T2处理)。

表5 不同生育期水分胁迫处理对日光温室葡萄果实物理特性和产量的影响
T1、T2、T3、T4和T5处理分别为萌芽期、新梢生长期、开花坐果期、果实膨大期和着色成熟期水分胁迫; CK为对照处理, 无水分胁迫。同列不同小写字母表示处理间差异显著(<0.05)。The water stress periods of T1, T2, T3, T4 and T5 treatments are bud burst, shoot elongation, flowering-fruit bearing, fruit enlargement and coloring maturity stages, respectively. CK is the control treatment, without water stress. Different lowercases in the same column indicate significant differences among treatments at 0.05 level.
3 讨论与结论
当植物受到水分胁迫(亏缺灌溉)时, 产生大量对细胞有毒害作用的氧自由基, 这种失衡的过量氧自由基造成植物蛋白质损伤、脂质过氧化、 DNA突变和酶失活等[16]。SOD作为植物体中的核心酶, 与土壤水分状况有关[17]。本研究表明, 在葡萄开花坐果期(营养生长和生殖生长养分竞争的最激烈时期)进行短时间(11 d)水分亏缺处理(T3), SOD活性显著降低; 在果实膨大期(T4)、着色成熟期(T5)等2个葡萄生殖生长高峰期长时段亏水处理后SOD活性较CK也显著下降; 葡萄萌芽期(T1)和新梢生长期(T2)亏水处理下这两个生育期SOD活性与CK无显著差异, 可能与这两个生育期属于营养生长阶段、尚未进入生殖生长, 且水分胁迫时间较短, 气温相对较低有关。说明SOD酶活性变化对水分亏缺的响应与水分亏缺时长和生育时期等有关, 在葡萄生殖生长阶段(开花坐果期、果实膨大期、着色成熟期等后3个生育期)水分亏缺会显著抑制叶片SOD酶活性, 对叶片产生不利影响。另外, 萌芽期T4、T5处理SOD活性显著低于CK, 可能由于试验温室的葡萄已连续进行3年的水分胁迫试验(水分胁迫试验小区不变), 葡萄为多年生植物, 上一年度生育后期(果实膨大期—着色成熟期)的水分胁迫对第2年萌芽期生长产生了持续影响。
叶片丙二醛(MDA)是膜脂过氧化作用的产物之一。其产生原因是叶片受到逆境条件或衰老的过程中出现过多的活性氧, 造成了细胞膜脂过氧化。其含量越高表示膜质过氧化作用的程度越强[18]。何岸镕等[19]研究表明葡萄不同生育期中度亏水, MDA含量开始显现差异性是在开花坐果期。本文研究结果与此基本一致, 即萌芽期各处理之间MDA含量无显著差异。萌芽期水分胁迫处理(T1)的葡萄叶片MDA含量在新梢生长期迅速升高, 而新梢生长期水分胁迫处理(T2)的MDA含量在开花坐果期达到最大值, 其显著高于CK, 说明上述生育期水分胁迫对叶片MDA含量的累积具有滞后性。果实膨大期胁迫处理(T4)和着色成熟期胁迫处理(T5)的葡萄叶片MDA含量均在胁迫时期达到最高, 这可能与这两个生育期时段长(分别为72 d和97 d), 水分胁迫持续时间较长, 使叶片MDA在胁迫时段持续积累有关。
脯氨酸(Pro)是植物渗透调节物质中有机溶质之一, 其功能在于逆境胁迫下, 植物积累大量Pro, 以保证原生质与外界环境渗透平衡, 进而保证细胞生理代谢活动的正常进行。植物从胁迫条件恢复时, 还可作为氮、碳及还原力的一种快速补偿来源[20-21]。薛吉全等[20]发现玉米(L.)水分胁迫期间游离Pro累积量与水分胁迫程度和生育期有关。本文研究发现, 不同生育期水分胁迫处理, 葡萄叶片累积Pro的情况各不相同。在萌芽期、开花坐果期2个短生育期水分胁迫对葡萄叶片Pro含量影响不大, 胁迫处理与CK不存在显著性差异。果实膨大期水分胁迫处理(T4)的叶片Pro含量在该生育期降至最低, 与CK呈显著性差异; 果实着色成熟期水分胁迫处理(T5)叶片Pro含量也在果实着色成熟期最低, 表明在葡萄的长生育期水分胁迫会降低葡萄叶片Pro含量。
Mccarthy[22]研究得出, 水分胁迫会抑制葡萄果粒生长, 减轻果粒重量。本文研究结果与其一致, 不同生育期水分亏缺葡萄果粒纵横径均较对照减小。赵霞等[3]研究表明不同生育期水分亏缺葡萄果粒果形指数均与CK无显著性差异, 果实膨大期中度亏水处理产量较对照显著降低40.2%。张芮等[23]研究表明, 萌芽期中度亏水处理产量最高, 显著高于果实膨大期和着色成熟期中度亏水处理(<0.05)。本文研究也表明, 葡萄萌芽期(T1)和新梢生长期(T2)水分胁迫处理产量高于对照(CK); 果实膨大期水分胁迫处理(T4)则导致产量显著低于CK、T1、T2处理。葡萄着色成熟期水分胁迫处理(T5)果实硬度在采收期最小, 主要原因是该生育期水分胁迫促进葡萄果实中糖分的积累, 有利于葡萄早熟, 导致硬度显著低于其他处理。由此可以看出, 葡萄生育前期耐旱能力较强, 适当控制土壤水分有利于葡萄高效稳产; 果实膨大期是葡萄对水分胁迫最敏感的时期, 干旱将导致减产[24-25]; 着色成熟期水分亏缺对促进果实糖分积累促进早熟有利。
因此, 综合单个生育期水分调亏处理保护酶(SOD)活性、膜质过氧化物(MDA)和渗透调节物质(Pro)含量及果实生长、产量等指标分析, 可以得出果实膨大期水分胁迫对葡萄叶片生理指标、果实生长发育和产量产生极其不利影响, 在生产实践中应予以避免; 萌芽期、新梢生长初期、着色成熟期可以进行适当的水分胁迫以达到节水高效生产目的。
[1] 张付春, 潘明启, 麦麦提阿卜拉·麦麦提图尔荪, 等. 浮尘天气对墨玉河流域葡萄叶片光合及水势的影响[J]. 中国生态农业学报, 2018, 26(7): 990–998 ZHANG F C, PAN M Q, MEMETABLA·M, et al. Effect of floating dust weather on leaf photosynthesis and water potential of grapes in Karakash River Basin[J]. Chinese Journal of Eco-Agriculture, 2018, 26(7): 990–998
[2] 翟衡, 杜远鹏, 孙庆华, 等. 论我国葡萄产业的发展[J]. 果树学报, 2007, 24(6): 820–825 ZHAI H, DU Y P, SUN Q H, et al. On the development situation of Chinese grape industry[J]. Journal of Fruit Science, 2007, 24(6): 820–825
[3] 赵霞, 张芮, 成自勇, 等. 不同生育期调亏灌溉对荒漠绿洲区葡萄生长、产量和品质的影响[J]. 灌溉排水学报, 2017, 36(10): 20–23 ZHAO X, ZHANG R, CHENG Z Y, et al. Effect of regulated deficit irrigation at different growth stages on growth and fruit quality of wine grape in desert oasis[J]. Journal of Irrigation and Drainage, 2017, 36(10): 20–23
[4] 刘昌明. 中国农业水问题: 若干研究重点与讨论[J]. 中国生态农业学报, 2014, 22(8): 875–879 LIU C M. Agricultural water issues in China — Discussions on research highlights[J]. Chinese Journal of Eco-Agriculture, 2014, 22(8): 875–879
[5] 董宝娣, 刘会灵, 王亚凯, 等. 作物高效用水生理生态调控机制研究[J]. 中国生态农业学报, 2018, 26(10): 1465–1475 DONG B D, LIU H L, WANG Y K, et al. Physio-ecological regulating mechanisms for highly efficient water use of crops[J]. Chinese Journal of Eco-Agriculture, 2018, 26(10): 1465–1475
[6] 殷常青, 费良军, 李渊, 等. 日光温室延后栽培葡萄滴灌灌溉制度试验研究[J]. 节水灌溉, 2016, (10): 39–43 YIN C Q, FEI L J, LI Y, et al. Research on drip irrigation schedule of delayed planting grape in sunlight greenhouse in Tianzhu sloping land[J]. Water Saving Irrigation, 2016, (10): 39–43
[7] 张芮, 王旺田, 吴玉霞, 等. 水分胁迫度及时期对设施延迟栽培葡萄耗水和产量的影响[J]. 农业工程学报, 2017, 33(1): 155–161 ZHANG R, WANG W T, WU Y X, et al. Effect of moisture stress level and stage on evapotranspiration and yield of grape under protected and delayed cultivation[J]. Transactions of the CSAE, 2017, 33(1): 155–161
[8] 张芮, 成自勇, 王旺田, 等. 水分胁迫对延后栽培葡萄果实生长的影响[J]. 华南农业大学学报, 2015, 36(6): 47–54 ZHANG R, CHENG Z Y, WANG W T, et al. Effect of water stress on grape fruit growth under delayed cultivation[J]. Journal of South China Agricultural University, 2015, 36(6): 47–54
[9] 徐斌, 张芮, 成自勇, 等. 不同生育期调亏灌溉对设施延后栽培葡萄生长发育及品质的影响[J]. 灌溉排水学报, 2015, 34(6): 86–89 XU B, ZHANG R, CHENG Z Y, et al. Effects of deficit irrigation on growth and quality of greenhouse grape under delayed cultivation in different stages[J]. Journal of Irrigation and Drainage, 2015, 34(6): 86–89
[10] 齐建波, 荣新民, 陈虎, 等. 不同水分胁迫水平对葡萄叶水势及生理指标的影响[J]. 石河子大学学报: 自然科学版, 2011, 29(4): 437–441 QI J B, RONG X M, CHEN H, et al. Effect of different water stress quantity on leaf water potential and the physiological indexes of grape[J]. Journal of Shihezi University: Natural Science, 2011, 29(4): 437–441
[11] 张正斌. 作物抗旱节水的生理遗传育种基础[M]. 北京: 科学出版社, 2003 ZHANG Z B. Basis of Physiological Genetic Breeding of Crop Water Saving[M]. Beijing: Science Press, 2003
[12] 郑睿, 康绍忠, 胡笑涛, 等. 水氮处理对荒漠绿洲区酿酒葡萄光合特性与产量的影响[J]. 农业工程学报, 2013, 29(4): 133–141 ZHENG R, KANG S Z, HU X T, et al. Effects of water and nitrogen conditions on the diurnal variation of photosynthesis characteristic and yield of grapevine in arid oasis region[J]. Transactions of the CSAE, 2013, 29(4): 133–141
[13] 邹琦. 植物生理学实验指导[M]. 北京: 中国农业出版社, 2000 ZOU Q. Experimental Instruction of Plant Physiology[M]. Beijing: China Agriculture Press, 2000
[14] 高俊凤. 植物生理学实验指导[M]. 北京: 高等教育出版社, 2006 GAO J F. Experimental Instruction of Plant Physiology[M]. Beijing: Higher Education Press, 2006
[15] 赵世杰, 史国安, 董新纯. 植物生理学实验指导[M]. 北京: 中国农业科学技术出版社, 2002 ZHAO S J, SHI G A, DONG X C. Experimental Instruction of Plant Physiology[M]. Beijing: China Agricultural Science and Technology Press, 2002
[16] 董亮, 何永志, 王远亮, 等. 超氧化物歧化酶(SOD)的应用研究进展[J]. 中国农业科技导报, 2013, 15(5): 53–58 DONG L, HE Y Z, WANG Y L, et al. Research progress on application of superoxide dismutase (SOD)[J]. Journal of Agricultural Science and Technology, 2013, 15(5): 53–58
[17] GIRDTHAI T, JOGLOY S, VORASOOT N, et al. Associations between physiological traits for drought tolerance and aflatoxin contamination in peanut genotypes under terminal drought[J]. Plant Breeding, 2010, 129(6): 693–699
[18] 杜彩艳, 段宗颜, 潘艳华, 等. 干旱胁迫对玉米苗期植株生长和保护酶活性的影响[J]. 干旱地区农业研究, 2015, 33(3): 124–129 DU C Y, DUAN Z Y, PAN Y H, et al. Effect of drought stress on growth and activities of antioxidant enzymes of maize seedling[J]. Agricultural Research in the Arid Areas, 2015, 33(3): 124–129
[19] 何岸镕, 安进强, 张芮, 等. 不同生育期水分调亏对设施延后栽培葡萄叶片保护系统及产量品质的影响[J]. 水土保持学报, 2016, 30(3): 196–201 HE A R, AN J Q, ZHANG R, et al. Effects of water deficit on leaf protecting system and quality yield of delayed grape cultivation during different growth stage[J]. Journal of Soil and Water Conservation, 2016, 30(3): 196–201
[20] 薛吉全, 任建宏, 马国胜, 等. 玉米不同生育期水分胁迫条件下脯氨酸变化与抗旱性的关系[J]. 西安联合大学学报, 2000, 3(2): 21–25 XUE J Q, REN J H, MA G S, et al. Relationship between proline’s change and drought resistance under the condition of water stress in different developmental stages in corn[J]. Journal of Xi’an United University, 2000, 3(2): 21–25
[21] 蔡昆争, 吴学祝, 骆世明. 不同生育期水分胁迫对水稻根叶渗透调节物质变化的影响[J]. 植物生态学报, 2008, 32(2): 491–500 CAI K Z, WU X Z, LUO S M. Effects of water stress on osmolytes at different growth stages in rice leaves and roots[J]. Journal of Plant Ecology, 2008, 32(2): 491–500
[22] MCCARTHY M G. The effect of transient water deficit on berry development of cv. Shiraz (L.)[J]. Australian Journal of Grape & Wine Research, 2010, 3(3): 2–8
[23] 张芮, 成自勇, 王旺田, 等. 不同生育期水分胁迫对延后栽培葡萄产量与品质的影响[J]. 农业工程学报, 2014, 30(24): 105–113 ZHANG R, CHENG Z Y, WANG W T, et al. Effect of water stress in different growth stages on grape yield and fruit quality under delayed cultivation facility[J]. Transactions of the CSAE, 2014, 30(24): 105–113
[24] 刘静霞, 张芮, 成自勇, 等. 不同生育期水分亏缺对酿酒葡萄产量及品质的效应研究[J]. 干旱地区农业研究, 2016, 34(3): 78–83 LIU J X, ZHANG R, CHENG Z Y, et al. Effect research of water deficit in different growth stage on wine grape yield and quality[J]. Agricultural Research in the Arid Areas, 2016, 34(3): 78–83
[25] 纪学伟, 成自勇, 赵霞, 等. 调亏灌溉对荒漠绿洲区滴灌酿酒葡萄产量及品质的影响[J]. 干旱区资源与环境, 2015, 29(4): 184–188 JI X W, CHENG Z Y, ZHAO X, et al. Effect of regulated deficit drip irrigation on yield and quality of wine grape in desert oasis[J]. Journal of Arid Land Resources and Environment, 2015, 29(4): 184–188
Effects of water stress at different growth stages on leaf physiological changes and fruit development of grape in greenhouse*
NIU Zuirong1, ZHAO Xia1, ZHANG Rui1**, WANG Wangtian2, WEN Wen1
(1. College of Water Conservancy and Hydropower Engineering, Gansu Agricultural University, Lanzhou 730070, China; 2. College of Life Science and Technology, Gansu Agricultural University, Lanzhou 730070, China)
Gansu Hexi Corridor is one of the main grape producing areas in China. In recent years, the greenhouse cultivation improved the price of table grape and farmers income. However, as a high water demand economic crop, grape has been affected by drought in Hexi Corridor. The key to solve the contradiction between grape planting and shortage of water resources is to promote the water-saving and efficient production mode of grape cultivation. In order to explore the efficient water management mode and water-saving production of table grape in greenhouse, the experiment was conducted in a 6-year-old vineyard with water stress (the soil moisture was controlled at 55% to 75% of field water holding capacity) at budburst stage (its duration was 14 d), shoot elongation stage (its duration was 20 d), flowering-fruit bearing stage (its duration was 11 d), fruit enlargement stage (its duration was 72 d), and coloring maturity stage (its duration was 97 d), respectively; while the other stages was at full irrigation (the soil moisture content was up to 75%-100% of field water holding capacity). And the full-irrigation during the whole growth period of grape was set as the contral treatment (CK). The activity of superoxide dismutase (SOD), contents of malondialdehyde (MDA) and proline (Pro), as well as fruit diameter, hardness of fruit, single fruit weight and yield were observed, which were used for study the response mechanism of grape leaf physiology and fruit yield to water stress. The results showed that the SOD activity and Pro content of grape leaves were significantly reduced by water stress at fruit enlargement stage and coloring maturity stage. The MDA content of grape leaves was also significantly increased by water stress during the above two long growing stages, which had a negative effect on grape leaves. The water stresses at budburst stage, shoot elongation and flowering-fruit bearing stages had no significant effects on SOD activity and Pro content of grape leaves, significantly impacted MDA content of grape leaves. The yield under water stress at fruit enlargement stage was 14 830 kg·hm-2, which decreased by more than 20% compared with CK, and treatments of water stress at budburst stage and shoot elongation stage. Therefore, the water stress during fruit expansion stage in greenhouse significantly decreased SOD activity and osmoregulation substances contents, increased MDA content, and led to yield decrease of grape. In Gansu Hexi Corridor, it was not suitable to apply water stress at fruit expansion stage for greenhouse grape. The water stress in the early growth period had no significant effect on the physiology of grape leaves and fruit.
Grape; Water stress; Growth stage; Leaf physiology; Fruit development; Greenhouse
S275.9

10.13930/j.cnki.cjea.190734
* 国家自然科学基金项目(51769001, 51569002, 31560552)资助
张芮, 主要研究方向为节水灌溉与水资源利用。E-mail: zhangrui@gsau.edu.cn
牛最荣, 主要研究方向为水文与水资源学。E-mail: niuzr@gsau.edu.cn
2019-11-01
2019-12-18
* This study was funded by the National Natural Science Foundation of China (51769001, 51569002, 31560552).
, E-mail: zhangrui@gsau.edu.cn
Nov. 1, 2019;
Dec. 18, 2019
牛最荣, 赵霞, 张芮, 王旺田, 温文. 水分胁迫对日光温室葡萄叶片生理变化及果实发育的影响[J]. 中国生态农业学报(中英文), 2020, 28(2): 265-271
NIU Z R, ZHAO X, ZHANG R, WANG W T, WEN W. Effects of water stress at different growth stages on leaf physiological changes and fruit development of grape in greenhouse[J]. Chinese Journal of Eco-Agriculture, 2020, 28(2): 265-271
猜你喜欢
农业知识(2022年9期)2022-10-13
农业工程学报(2022年7期)2022-07-09
煤气与热力(2021年10期)2021-12-02
今日农业(2021年9期)2021-11-26
落叶果树(2021年6期)2021-02-12
今日农业(2020年16期)2020-12-14
房地产导刊(2020年9期)2020-10-28
电影(2018年10期)2018-10-26
中国计算机报(2017年47期)2018-01-12
电影文学(2016年23期)2017-02-13
