
伪足、有氧糖酵解与肿瘤细胞迁移的研究进展
2020-02-12 05:18:06滕春莹单毓娟
癌变·畸变·突变 2020年1期
黄 蕾,滕春莹,雷 鹏,单毓娟
(哈尔滨工业大学,黑龙江 哈尔滨 150001)
肿瘤细胞运动是由若干化学诱导物刺激胞内信号通路,使细胞骨架肌动蛋白重组形成膜突起继而引发的多步骤、多阶段的复杂过程[1]。肿瘤细胞运动的原动力来源于细胞骨架蛋白,它是形成细胞伪足的核心成分。作为细胞运动的执行者,伪足可形象地被称为细胞运动的“脚”,其结构特征是肿瘤细胞迁移能力强弱的关键所在[2]。
1 伪足与肿瘤细胞迁移
1.1 伪足分类、形成及功能
参与肿瘤细胞运动的伪足主要有4种:片状伪足、丝状伪足、侵袭性伪足和足体。如图1所示,片状伪足是细胞表面较宽且短的板状膜突起,又叫做细胞膜波动,富含动力型的肌动蛋白,其内部的肌动蛋白呈网络状排列;丝状伪足是细胞表面延伸的细长呈针状的膜突起,其内部充满平行紧密排列的丝状纤维[3]。此外,还有些膜突起可以降解和重构细胞外基质,这类侵袭性细胞在正对基底膜一侧形成的富含肌动蛋白的膜突起叫侵袭性伪足,它仅在细胞侵袭过程中出现,且无固定形态。目前认为丝状伪足是由片状伪足发展而来,而足体被认为是侵袭性伪足的前体。
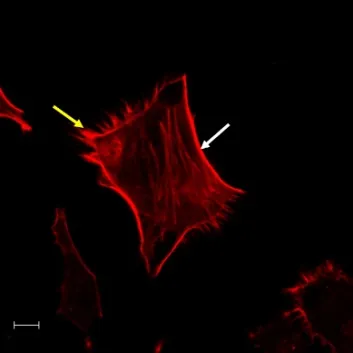
图1 细胞伪足形态
片状伪足的膜突起是由肌动蛋白的局部聚合引起的,首先是肌动蛋白单体(G-actin)聚集形成肌动蛋白纤维(F-actin),同时在前缘产生肌动蛋白丝的游离带刺末端,继而促进肿瘤细胞的迁移。其中产生游离带刺末端的方式主要有以下3种:肌动蛋白相关蛋白2/3复合物(actin-related protein complex,Arp2/3 complex)调控肌动蛋白重新聚集;丝切蛋白切断已存在的肌动蛋白纤维;肌动蛋白纤维的脱帽作用[2,4]。片状伪足与丝状伪足的核心成分是肌动蛋白纤维,而侵袭伪足的成分较为复杂,由一系列蛋白共同构成,如肌动蛋白、肌动蛋白调控蛋白、黏附分子、膜重塑和信号蛋白及基质降解酶等,其中核心蛋白为皮层肌动蛋白(Cortactin)和神经Wiskott-Aldrich综合征蛋白(neural Wiskott-Aldrich syndrome protein,N-WASP),这些蛋白在多种恶性肿瘤中表达均上调。足体在外观和分子构成与侵袭性伪足相似。典型的足体由单核型细胞形成,此外在平滑肌细胞和内皮细胞也发现了类似于足体的结构。
作为细胞膜的特殊形式,这几种伪足有着不同的功能:丝状伪足主要负责细胞黏附和摄取营养,而片状伪足对于细胞的长距离迁移起着重要的作用[5];侵袭性伪足只有在细胞侵袭的过程中出现,可帮助肿瘤细胞侵入并通过基质进入血管。足体与侵袭性伪足相同,也能够降解细胞外基质[6-7]。这些伪足之间的关系目前报道较少,已有文献认为丝状伪足与片状伪足之间存在着互相转换的关系,这种转换直接调节细胞的运动,在癌细胞快速转移过程中能够发现大量的丝状伪足向片状伪足的转换[2,8]。细胞伪足总结见表1。

表1 细胞伪足概况
1.2 伪足形成的调节与肿瘤细胞迁移
对伪足形成的研究,主要是围绕伪足核心成分及主要调控因子展开的。皮层肌动蛋白是一种肌动蛋白结合蛋白,通过磷酸化而被激活,活化后可招募Arp2/3复合物,继而促进肌动蛋白细胞骨架的聚合和重组[9]。其中Arp2/3复合物是一种七亚基蛋白复合物,可促进肌动蛋白纤维聚合过程中的成核和分支化,并能被WASP家族蛋白激活,在调控肌动蛋白细胞骨架中起重要作用。近期研究发现,Cortactin在胶质瘤组织中的表达明显高于非肿瘤组织,且与肿瘤的恶性程度呈正相关;通过RNAi沉默Cortactin后,胶质瘤细胞的片状伪足大小和存留时间均有所减少,表明Cortactin可通过调控片状伪足的形成促进胶质瘤细胞的迁移[10]。此外,Cortactin还作为侵袭伪足的核心成分之一,参与调控肿瘤细胞的侵袭过程[1]。Lin等[11]发现,在乳腺癌细胞中足细胞标志蛋白1可促进Cortactin磷酸化,并通过激活Rac1/Cdc42/Cortactin信号通路促进侵袭伪足形成及肿瘤细胞的迁移。在哺乳动物细胞中,WASP家族蛋白由5个成员组成(WASP、N-WASP、WAVE 1、WAVE 2和WAVE 3),其中N-WASP是构成侵袭伪足的另一核心成分,活化的N-WASP可以起到帮助侵袭伪足降解细胞外基质的作用。在大鼠乳腺癌细胞MTLn3中,敲除N-WASP或其上下游效应器(Cdc42,WIP或Arp2/3复合物),均可显著抑制侵袭伪足的形成和基质降解活性[12]。Huang等[13]发现,通过转染siRNA阻断WAVE2的表达,可显著抑制纤维肉瘤HT-1080细胞中片状伪足的形成及细胞迁移运动。
Rho GTP酶是Ras超家族的成员,目前已发现20多种。其中,Cdc42、Rac1和Rho A是研究最多的。采用RNAi技术抑制前列腺癌细胞Rho A表达后,细胞形态变为细长,并延伸出多个丝状伪足,且肿瘤侵袭性增强;相反,当抑制Rho C表达时,细胞则呈椭圆状,片状伪足增多,这表明不同Rho GTP酶可调控不同的伪足形成[14]。Yuki[15]等发现,p39-Cdk5可通过降低Rac1活性来抑制小鼠成神经细胞瘤的片状伪足形成。此外,Cdc42和Rac1在卵巢肿瘤组织中均高表达;使用酮咯酸R抑制Cdc42和Rac1活性后,卵巢癌细胞中Cdc42介导的丝状伪足减少,细胞迁移能力降低[16]。Rho GTP酶在细胞骨架重组调控以及细胞运动等方面起重要作用。根据现有文献资料,将片状伪足、丝状伪足和侵袭伪足的主要调控通路总结如下。如图2所示,片状伪足的形成主要由Rho A-ROCK或Cortactin-Arp2/3介导的actin聚合调控;丝状伪足形成主要由Cdc42-WASPArp2/3通路调控;而侵袭性伪足的形成主要是由其核心成分Cortactin和N-WASP促进Arp2/3复合物而调控的。综上所述,干扰细胞伪足的形成能够从根本上阻断细胞运动的驱动力,从源头上抑制肿瘤细胞的侵袭和转移。

图2 伪足形成的调控信号通路
2 有氧糖酵解与肿瘤细胞迁移
肿瘤细胞迁移不仅与伪足的核心成分有关,还需要足够能量的供应。在生理条件下,伪足核心成分G-actin与ATP结合形成ATP-actin,牢固的聚集在原有F-actin的前端,随后ATP水解变成结合能力较弱的ADP-actin而脱落,这个动态循环的过程使细胞获得可以向前运动的驱动力(图3)[17]。因此,肿瘤细胞若想快速迁移,大量能量的获取是必不可少的。正常情况下,每消耗1个葡萄糖分子,低氧环境中无氧糖酵解可产生2个ATP分子;而常氧环境中经过线粒体电子传递链彻底氧化则可产生36个ATP分子。肿瘤细胞的快速增殖和迁移造成细胞内的低氧环境,同时还需要高速率的产能。这种需求与实际情况相矛盾,使得肿瘤细胞获得重新编排自身能量代谢的能力,即能量代谢重排。此时,肿瘤细胞通常会打破生理状态下的能量代谢模式,呈现如下异常特征:糖酵解升高、氨基酸和脂质代谢增强、线粒体生物合成及戊糖磷酸途径增强等。能量代谢重排被视为肿瘤的一个新特征[18]。其中,“Warburg效应”又称为有氧糖酵解(即使在氧气充足的情况下,肿瘤细胞仍优先利用糖酵解供能),是具有较强迁移能力的恶性肿瘤细胞普遍具有的一个代谢重排特征。
糖酵解异常升高是大多数肿瘤细胞能量代谢一个备受关注的特征。图4概括了细胞内葡萄糖代谢的基本过程及参与调控的主要调节因子,葡萄糖代谢为乳酸的产能效率远低于氧化分解为二氧化碳和水。因此,部分肿瘤细胞通过上调葡萄糖转运蛋白(GLUT-1、GLUT-2、GLUT-3和GLUT-4)来摄取更多的葡萄糖,以维持ATP的供应[19]。葡萄糖摄取增加是区别肿瘤细胞和正常细胞的主要特征。据报道,Ras、Myc和HIF-1α等癌基因是肿瘤细胞糖酵解的主要诱导因子[20]。其中c-Myc和低氧诱导因子(HIF-1α)这两种主要的糖酵解激活转录因子可协同促进肿瘤糖酵解酶的表达,如己糖激酶(HK2)、磷酸果糖激酶1(PFK-1)、乳酸脱氢酶A(LDHA)等[20-21]。虽然HIF-1α主要在低氧环境中发挥作用,但c-Myc可在氧气充足环境中促进糖酵解相关基因的表达。这种协同作用使得肿瘤能够持续促进糖酵解代谢,从而支持其快速增殖和迁移运动[22]。相反,p53则可通过直接抑制GLUT-1和GLUT-4的转录来下调葡萄糖的摄取[23]。因此,p53、c-Myc和HIF-1α之间的平衡对肿瘤细胞糖酵解的状态有决定性的影响。

图4 细胞葡萄糖代谢过程及调控
糖酵解酶在肿瘤迁移过程中起着重要作用。丙酮酸激酶(PK),催化糖酵解的最终限速步骤。在分化的胃癌细胞中,敲除PKM2可以降低E-钙黏蛋白(E-cadherin)的表达,从而激活EGFR下游信号通路(如PLC-γ1和ERK1/2),抑制肿瘤细胞的迁移和侵袭[24]。此外,在结直肠癌中PKM2表达异常升高,这与肿瘤淋巴转移相关,敲除PKM2可抑制结肠癌细胞的增殖和迁移[25]。乳酸脱氢酶(LDH),催化丙酮酸盐向乳酸转化的关键代谢酶;转化生长因子-β2(TGF-β2)是高级别胶质瘤侵袭的重要调节因子。LDHA和乳酸可以调节胶质母细胞瘤细胞中TGF-β2的表达,增加基质金属蛋白酶(MMP2)的表达,继而增强胶质瘤细胞的迁移[26]。除上述糖酵解酶外,其他糖酵解酶也在肿瘤细胞迁移过程中发挥潜在作用。据报道,己糖激酶2(HK2)和6-磷酸果糖-2-激酶(PFKFB)均是HIF-1的转录靶点。
糖酵解增强的另一个重要后果就是乳酸的大量堆积。过多的乳酸通过单羧酸转运蛋白(MCTs)转出细胞,导致细胞外微环境酸化,pH值降低,基质金属蛋白酶(MMPs),尿激酶型纤溶酶原激活剂和组织蛋白酶B、D和L等蛋白酶激活,进而诱导细胞外基质降解并促进肿瘤细胞转移。此外,乳酸不仅是一种代谢中间产物,而且是一种信号分子,能激活HIF[26]。除了诱导糖酵解相关酶的表达,HIF还可调节控制多种细胞功能的基因,如血管生成、能量代谢、侵袭和转移、凋亡或存活等。
糖酵解不仅为癌细胞提供能量,而且为生物合成提供必要的前体。糖酵解代谢产物,如葡萄糖-6-磷酸、磷酸二羟丙酮等,可以被转移到其他代谢途径。例如,磷酸戊糖途径可利用葡萄糖-6-磷酸合成核苷酸和NADPH,其中NADPH是一种重要的还原剂,对氧化还原稳态十分重要。磷酸二羟丙酮可用于脂质合成,对组装新的细胞器并促进肿瘤生长具有重要的意义。此外,糖酵解代谢产物也是氨基酸生产和大分子合成的重要原料[26]。除了维持代谢功能外,糖酵解酶在促进癌症生存、转移、侵袭、染色质重塑、基因表达调控和其他必要的细胞过程中均发挥着积极的促进作用[25]。因此,靶向糖酵解酶的表达可能是癌症治疗的有效策略。目前,通过糖酵解靶向肿瘤细胞伪足形成的研究甚少。据报道,前列腺癌和内皮细胞依赖糖酵解来调节片状伪足的形成和细胞运动[27-28]。在糖酵解被抑制的情况下,线粒体产生的ATP不能被运用到细胞骨架重塑的位点上,这表明葡萄糖代谢在细胞骨架重塑和细胞运动中起着基础作用[27]。
3 总结
复发及转移是导致癌症患者不良预后的主要原因。细胞伪足是肿瘤迁移运动能力强弱的关键所在,伪足的形成过程需要大量ATP供应。可见,充足的能量获取是肿瘤细胞运动必不可少的驱动力。目前大量研究证实,多数肿瘤细胞都存在能量代谢重排现象,以满足肿瘤细胞快速增殖及运动迁移的需求。因此,通过靶向能量代谢重排来研究伪足形成对肿瘤细胞转移的影响已经成为肿瘤防治的一个新策略。
猜你喜欢
中南医学科学杂志(2022年6期)2022-12-04 12:01:20
生物化工(2022年4期)2022-09-20 09:18:08
中国畜牧杂志(2020年1期)2020-01-16 04:09:54
中国循证心血管医学杂志(2020年11期)2020-01-08 11:38:42
中华老年口腔医学杂志(2016年4期)2017-01-15 14:25:05
中国医学装备(2016年6期)2016-12-01 06:44:29
中外医疗(2015年5期)2016-01-04 03:57:57
恋爱婚姻家庭·养生版(2015年10期)2015-05-14 21:46:23
化工管理(2015年6期)2015-03-23 06:03:38
中国医科大学学报(2015年10期)2015-03-01 02:09:59
