
马氏珠母贝黄壳色家系的生长、壳色分离及总抗氧化能力比较
2020-02-07 17:56刘文广潘肖兰刘惠茹何毛贤
海洋科学 2020年1期
许 濛 , 黄 文 , 刘文广 , 潘肖兰 , 刘惠茹 , 何毛贤
(1. 中国科学院 南海海洋研究所, 热带海洋生物资源与生态重点实验室, 广东省应用海洋生物学重点实验室, 广东 广州 510301; 2. 中国科学院大学, 北京 100049; 3. 中国科学院 南海生态环境工程创新研究院, 广东 广州 510301)
近年来, 人工养殖的马氏珠母贝(Pinctada fucata martensii)存在生长过缓、死亡率高、珍珠品质退化等问题[1], 定向人工选育有助于贝的遗传性状改良[2]和种质资源保护。我国目前以生长为主要选育目标的马氏珠母贝新品种已培育出4个(“海优1号”[3]、“海选1号”[4]、“南科1号”[5]和“南珍1号”[6])。马氏珠母贝具有红色[7-9]、白色[10-11]、黄色[12-14]、褐色[10-11,14]和黑色[9,14-15]等壳色, 除生长性状外, 对于壳色的选育也有报道, 如Wada 和Komaru通过选育实验发现马氏珠母贝的白壳色相对常见褐壳色是隐性纯合子[16], 白壳自交系相比褐色-白色杂交系体重增长和存活率较差[10], 但白壳贝作为供体小片产生黄色珍珠的概率显著低于褐壳贝[11]。类似地, 马氏珠母贝黄壳色性状会影响珍珠质量[17], 而且与生长性状存在关联[13,18-20], 我们在前期工作中也从马氏珠母贝野生群体中发现了极少量的黄壳色个体, 并构建了黄壳家系, 但目前对黄壳性状的壳色分离还未有报道。
有报道指出贝类的浅色类型存在生活力劣势[16,21], 黄壳色马氏珠母贝是否也是如此还不明确。抗氧化能力(total antioxidant capacity, TAC)是评估生物体免疫水平和环境污染程度的重要指标, 铁离子还原法(ferric ion reducing antioxidant power, FRAP)是测定TAC的常用方法, 在中草药植物[22]、人体[23]、鹿角珊瑚[24]和海洋贝类[25]中均有报道, Zheng等[26]通过FRAP法评价华贵栉孔扇贝(Chlamys nobilis)的TAC, 认为橙色扇贝相比褐色扇贝的卵巢中TCC和TAC含量更高。马氏珠母贝抗氧化性、抗逆性与壳色的关系目前已有少量报道, 白成等[15]认为马氏珠母贝的黑壳贝在温度、盐度胁迫下抗逆能力优于黄壳、红壳、白壳贝, 孟子豪等[27]认为高温胁迫下合浦珠母贝(Pinctada fucata)的总类胡萝卜素含量(TCC)与总抗氧化能力正相关, 但高TCC家系的抗氧化酶如SOD、CAT活性以及丙二醛含量均显著低于低TCC家系。
壳色作为易于观测的表型性状, 对壳色和生长性状进行协同选择和定向选育, 有助于培育易于鉴别的优良品种。本文拟研究马氏珠母贝黄壳色家系壳色遗传分离、生长及TAC含量, 以期为马氏珠母贝壳色品种的选育提供基础资料。
1 材料与方法
1.1 第一代黄壳家系的构建
第一代黄壳家系(F1)属于父本半同胞家系, 父本和母本均来源于深圳野生群体, 黄壳色个体外面观为黄色底色, 常见褐壳色个体外面观为褐色底色(图1)。1-3号家系是同一只黄壳色父本, 4-6号家系是另一只黄壳色父本, 母本是同一野生群体的黄壳色/褐壳色个体(交配组合见图2)。同时构建的还有35个褐壳家系, 亲本均为褐壳。

图1 马氏珠母贝黄壳家系中黄壳色(上方两排)和褐壳色个体(下方两排) Fig. 1 Yellow individuals (upper two rows) and brown individuals (lower two rows) in yellow shell families of Pinctada fucata martensii
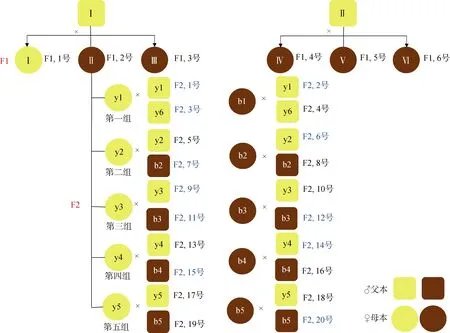
图2 第二代家系构建示意图 Fig. 2 Progeny tree of the established F2 families
1.2 第二代黄壳家系构建
第二代家系(F2)是以F1代2号家系中的黄壳、 褐壳色个体为亲本进行构建。第一组黄壳色父本(♂)2个, 黄壳、褐壳色母本各1个; 其余4个双列杂交组, 每组包含黄壳雌雄亲本各1个, 褐壳雌雄亲本各1个, 共构建20个家系。培育成功的家系有11个, 分别是黄×黄家系, 编号1、3、9; 黄×褐家系, 编号7、11、15; 褐×黄家系, 编号2、6、14; 褐×褐家系, 编号12、20(见图2)。
1.3 养殖及数据测量
所有家系的授精、孵化、育苗和养成在中国科学院大亚湾海洋生物综合实验站(深圳)进行, 期间的光照、水温、海区和养殖管理等相同, 不同家系严格隔离、分开笼养。分别在5月龄、7月龄、11月龄、14月龄、20月龄时, 测量F1黄壳家系和其他褐壳色家系(每个家系n=50)的4个生长性状, 用游标卡尺测量壳长、壳高、壳宽(精度为±0.01 mm), 电子天平测量总质量(精度为±0.01 g), 比较家系间生长差异; 另外, 在7月龄测定时, 对F1黄壳家系(n=50, 其中5号家系n=47)中黄壳、褐壳个体进行区分, 比较家系内不同壳色个体的4个生长性状。在F1、F2的5月龄时, 对家系所有个体的壳色进行统计。
1.4 FRAP法测定黄壳、褐壳贝不同组织的总抗氧化能力(TAC)
根据铁离子氧化还原能力(ferric reducing antioxidant power, FRAP) 评价马氏珠母贝不同组织的抗氧化能力[28], 操作按照试剂盒(碧云天S0116, 中国)说明书进行。首先配制FRAP工作液, 并在37℃孵育; 其次取F3代18月龄黄壳、褐壳色成贝各7只, 取鳃、足、心、闭壳肌、外套膜、消化腺、性腺这7个组织样, 加入500 μL预冷的PBS溶液并进行匀浆处理, 以充分破碎组织并释放其中的抗氧化物, 4℃下12 000g 离心10 min, 收集上清液, 5倍稀释后, 用96孔板在酶标仪(Tecan infinite 200Pro, 瑞士)上进行测定, 每个样品设3个生物学重复; 再次, 所有上清液稀释5倍后, 用BCA法(生工C503051, 中国)测定各组织中总蛋白含量, 以牛血清白蛋白(BSA, 0.025~1 g/L)作为标准品绘制标准曲线, 在562 nm下测定标准品和待测样品的吸光度值Cp。最后, 另取一个96孔板, 加入上清和FRAP工作液37℃孵育5 min, 以梯度稀释(0.15~2 mmol/L)的FeSO4·7H2O作为标准品绘制标准曲线, 在593 nm下测定吸光度值Ct。TAC(单位蛋白浓度下铁离子氧化还原能力)按照以下公式进行计算[26,28]:
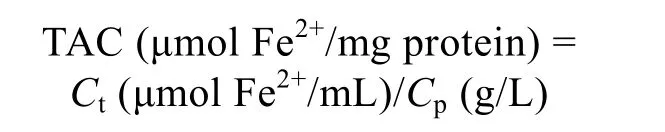
1.5 数据分析
家系的生长指标通过SPSS 16.0软件进行方差分析和t-检验, 壳色分离比采用卡方检验, 显著性水平包括差异显著(P<0.05)和差异极显著(P<0.01)。
2 实验结果
2.1 F1黄壳家系与褐壳色家系的生长性状比较
F1黄壳家系和对照组褐壳家系的4个生长性状的均值见表1。第一代黄壳家系(F1)5、7月龄贝仅壳宽值显著小于褐壳家系 (P<0.05), 壳高、壳长、总质量差异不显著, 11月龄贝仅壳长和壳宽值显著小于褐壳家系, 14、20月龄贝壳高、壳长、壳宽、总质量这4个生长性状数值均极显著小于褐壳家系(P<0.01), 见表1。
2.2 F1黄壳家系中黄壳、褐壳个体的生长性状比较
7月龄F1黄壳家系内黄壳个体和褐壳个体的生长指标见表2。经统计检验, F1代6个黄壳家系内黄壳个体与褐壳个体在壳高、壳长、壳宽和总质量上差异都不显著(P>0.05)。
2.3 F1黄壳家系壳色性状分离情况统计
在这6个F1家系中, 亲本之一为黄壳色, 则后代中会出现黄壳色贝, 黄壳贝的比例为42.11%~76.80%, 平均57.63%; 两个亲本为黄壳色, 后代中出现了黄、褐壳色, 即黄壳色性状发生了分离(表3)。第一代所有褐壳家系未产生黄壳色子代(数据未展示)。1、2、6号家系子代壳色分离实际比例不等于预测比例, 卡方适合性检验统计值P<0.01, 认为偏离孟德尔遗传分离定律, 而3、4、5号家系子代壳色实际比例接近预测比例, 认为符合分离定律。
2.4 F2家系壳色性状分离情况统计
在F2家系中, 亲本均为褐壳色的两个家系(编号12、20)没有产生黄壳色的子代, 推测褐壳色为纯合子; 亲本均为黄壳色的家系(编号1、3、9), 其子代呈现了黄、褐两种壳色, 即发生了壳色分离; 亲本为黄壳色和褐壳色的家系(编号2、6、14、7、11、15), 其子代也呈现了黄、褐两种壳色。除3、6、11号F2家系偏离孟德尔遗传分离定律外(P<0.01), 其他5个家系的壳色遗传符合孟德尔分离定律(表4)。 黄壳贝的比例为48.54%~98.88%, 平均70.22%。
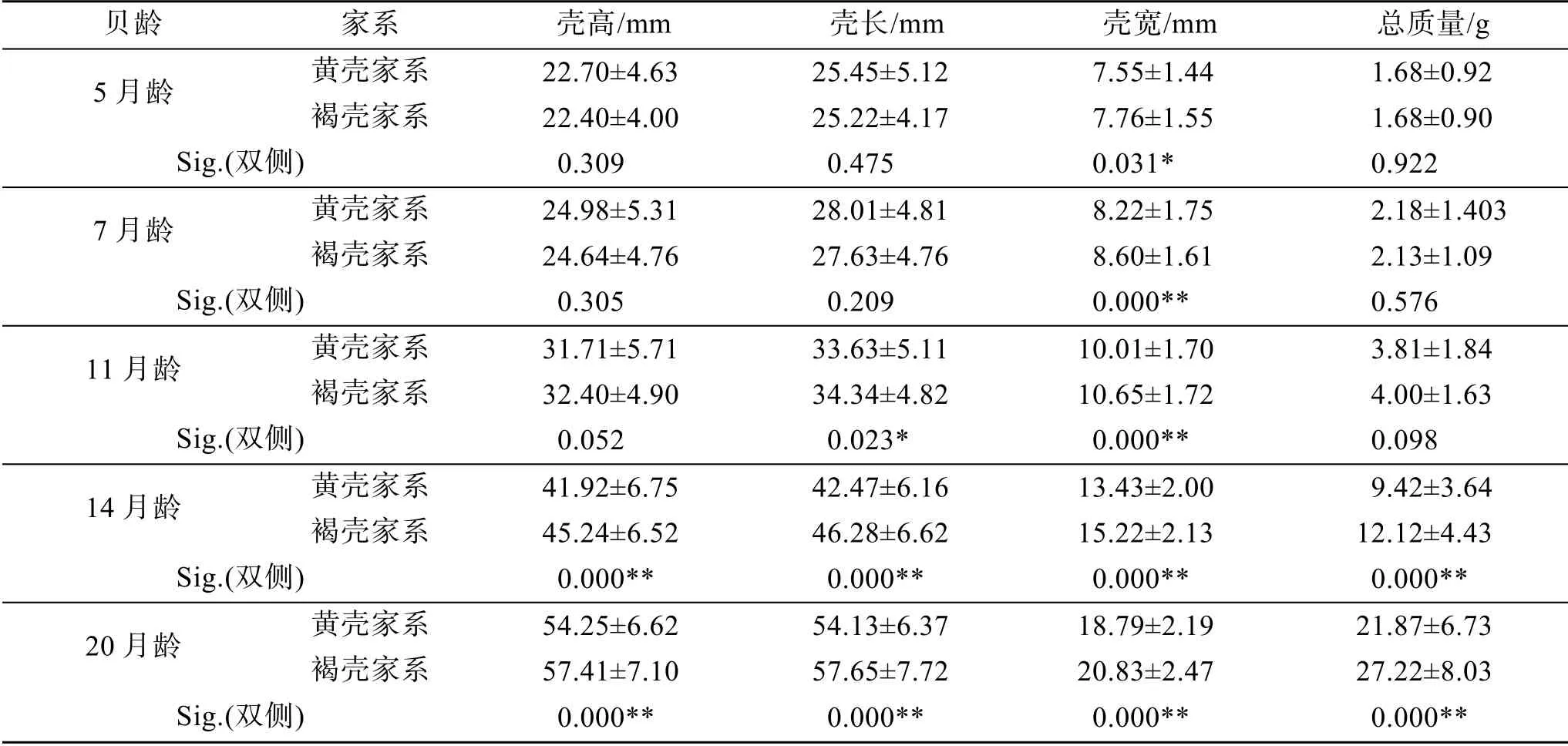
表1 第一代黄壳家系与褐壳家系4个生长性状的统计学分析 Tab. 1 Statistical analysis of the four growth traits between the F1 yellow shell families and brown shell families
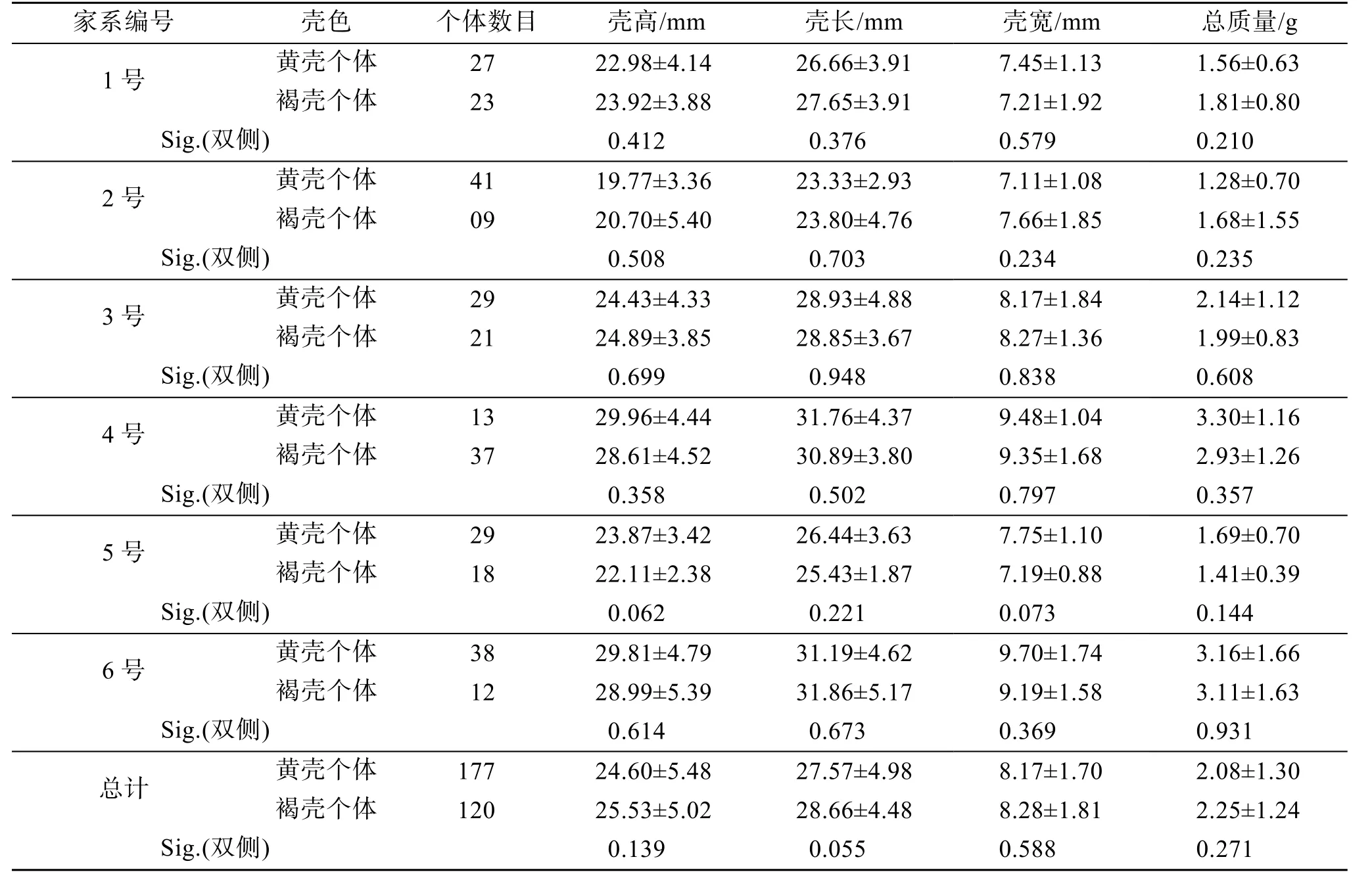
表2 F1黄壳家系中黄壳、褐壳色个体的生长统计学分析 Tab. 2 Statistical analysis of the four growth traits between yellow and brown individuals in the F1 yellow shell families

表3 F1黄壳家系壳色性状分离情况统计 Tab. 3 Statistical analysis of shell color segregation in the F1 yellow shell families
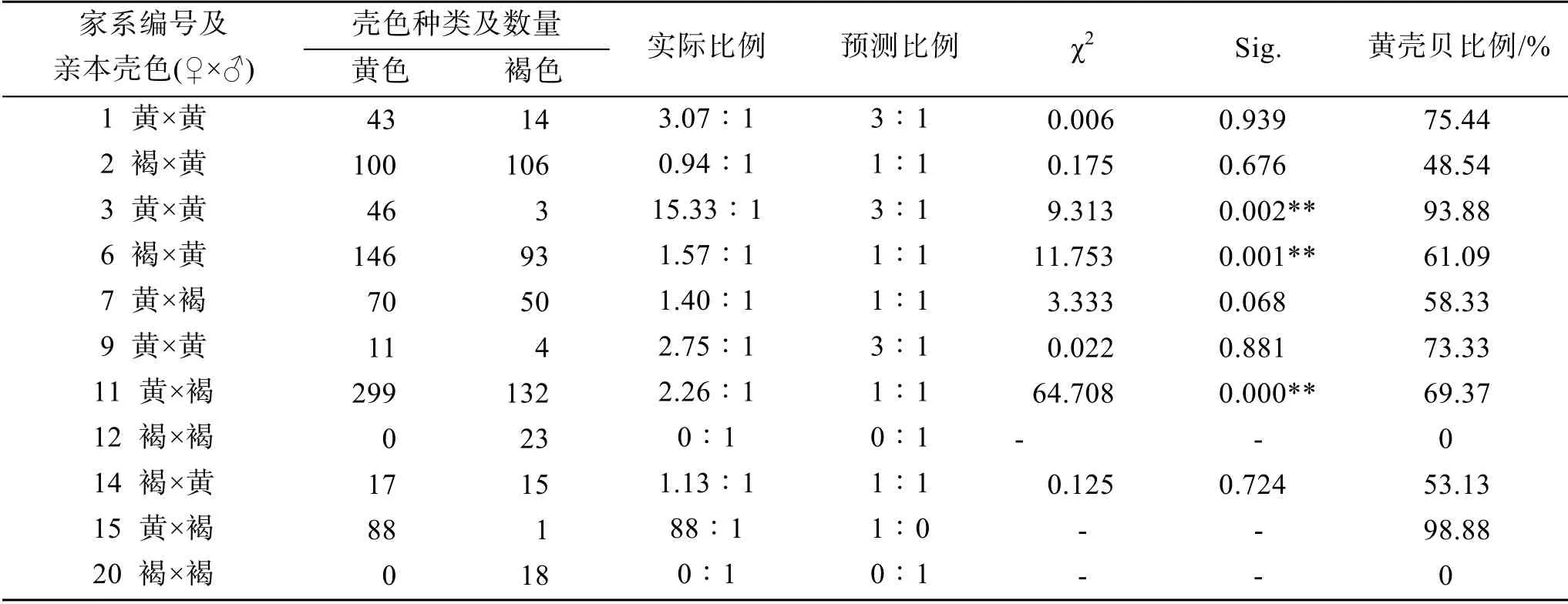
表4 F2家系壳色性状分离情况统计 Tab. 4 Statistical analysis of shell color segregation in the F2 families
2.5 黄壳家系内黄壳、褐壳个体不同组织的抗氧化能力比较
两标准品的标准曲线拟合出的线性回归方程见图3, R2值均不小于0.99。测定不同组织中单位蛋白含量的铁离子氧化还原能力后发现, 黄色壳马氏珠母贝在鳃和闭壳肌组织中TAC值较高。在足、心、性腺组织中黄壳贝TAC显著低于褐壳贝(P<0.05), 在鳃、闭壳肌、消化腺组织中黄壳贝TAC高于褐壳贝, 在外套膜组织中黄壳贝低于褐壳贝, 但差异都不显著(见图4)。
3 分析与讨论
3.1 马氏珠母贝黄壳色和褐壳色家系及个体的生长比较分析
马氏珠母贝壳色与生长的关联分析已有报道, 王庆恒等[18]、邓岳文等[13]、朱晓闻等[19]发现黄壳选育系生长性状均显著小于褐壳贝。本文中F1代6个黄壳家系, 在5、7月龄贝仅壳宽值显著小于褐壳家系(P<0.05), 11月龄贝仅壳长和壳宽值显著偏小, 14、20月龄贝的壳高、壳长、壳宽、总质量这4个生长性状均极显著偏小(P<0.01), 这一结果与邓岳文等[13,18]黄壳选育系F1情况类似。稚贝发育至成贝过程中, 黄壳家系和褐壳家系的差异也愈加显著, 可能是黄壳色对贝的生长影响存在累积效应。另外, 本文F1代 6个父本半同胞家系内, 黄壳、褐壳个体生长差异并不显著, 这一结果表明在亲本相同、且养殖环境、密度等条件一致的前提下, 黄壳个体具有与褐壳个体近似的生长性能。
马氏珠母贝中发现白色贝类[16]具有生活力劣势, 本文中F1代黄壳家系相比褐壳家系存在生长劣势, 但亲本相同时黄壳贝相比褐壳贝未见生长劣势, 可能是家系选育刚刚开始, 纯化不够。这一结果反映黄壳色亲本的引入, 会对全部不同壳色子代的生长性状产生影响, 再次证明黄壳色与生长性状紧密关联。
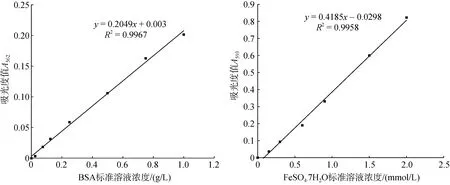
图3 测定组织总蛋白含量绘制BSA标准曲线 (左), 测定铁离子总抗氧化活性绘制FeSO4.7H2O标准曲线(右) Fig. 3 BSA standard curve for the determination of total protein content in tissues (left); FeSO4.7H2O standard curve for the determination of ferric ion reducing antioxidant power (right)

图4 马氏珠母贝黄壳、褐壳贝不同组织的总抗氧化能力比较 Fig. 4 Comparison of total antioxidant capacity in different tissues of yellow and brown oysters of Pinctada fucata martensii
3.2 马氏珠母贝黄壳色的遗传与分离
本文中, 从两种壳色的正反交壳色分离结果可认为黄壳色不存在伴性遗传现象。F1和F2中褐×褐家系壳色不分离, 推测褐壳色是隐性纯合子; F1代5个褐×黄家系子代均有褐壳个体产生, 推测半同胞家系中的2个黄壳父本都是杂合子。11个F2代家系中, 2个褐×褐家系(编号12、20))壳色不分离, 3个黄×黄家系有1个(编号3)偏离孟德尔遗传定律, 6个黄×褐(或褐×黄)家系有2个家系(编号6、11)偏离。F2代有5个家系的壳色分离可采用“单位点二等位基因”模型解释, 即黄壳色与褐壳色由单基因控制, 且黄壳色对褐壳色具有显性效应。
贝类壳色遗传存在不同的遗传模式, Ge等[21,29]发现长牡蛎(Crassostrea gigas)壳色遗传也遵循“单位点二等位基因”模型, 金黄色作为背景色对白色背景色为显性。Zheng等[30]研究了华贵栉孔扇贝(Chlamys nobilis)橙色、橙紫色、紫色和棕色四种壳色遗传规律时, 认为其壳色遗传符合“单位点三等位基因”的遗传模式, 橙色可以遮盖其他三种颜色, 橙紫色对棕色和紫色为显性, 紫色对棕色为显性。Winkler等[31]研究了智利扇贝(Argopecten purpuratus)的壳色遗传模式, 认为壳背景色由一个基因控制, 而白色放射带由另一个隐性等位基因控制。Ky等[32-33]认为珠母贝(Pinctada margaritifera)的黑壳、红壳、白壳色符合“单位点三等位基因”遗传模式, 肉色是不同于壳色的另一性状。Kozminskii 等[34]发现单基因遗传假说不足以解释滨螺(Littorina obtusata)的壳色遗传规律, 提出了“两位点二等位基因”假说。
本文中F1代和F2代均有3个家系偏离孟德尔遗传定律, 可能原因是贝的黄壳色是多个性状综合作用的结果, 需要对黄壳色个体进一步区分为黄壳色有/无放射线等类型; 偏离家系还可能与显性上位作用相关; 家系子代死亡率差异也可能对壳色分离比统计有所影响。对于壳色偏分离需更细致深入的研究。
3.3 马氏珠母贝黄壳色和褐壳色个体的总抗氧化能力比较
张博等[25]通过FRAP法测定栉江珧(Atrina pectinata)、毛蚶(Scapharca subcrenata)、台湾日月贝(Amusium japonicum taiwanicum)、菲律宾蛤仔(Ruditapes philippinarum)中鳃、闭壳肌、外套膜3种组织的TAC差异发现, 这4种贝均为鳃最高、闭壳肌次之、外套膜最低。本文测定了黄壳贝和黑壳贝不同组织中的TAC值, 也是黄壳贝鳃和闭壳肌的TAC值较高, 这与张博等[25]的报道基本吻合。在足、心、性腺组织中黄壳贝TAC显著低于褐壳贝(P<0.05); 在鳃、闭壳肌、消化腺组织中黄壳贝TAC高于褐壳贝, 在外套膜组织中黄壳贝低于褐壳贝但差异不显著。Xu等[35]测定了马氏珠母贝不同组织的总类胡萝卜素含量(TCC), 鳃、闭壳肌组织中黄壳贝TCC和TAC含量均高于褐壳贝, 但足和性腺中TCC和TAC的趋势相反, 推测可能是TAC不仅与总类胡萝卜素相关, 还可能与抗氧化酶、自由基、脂质等因素相关。
猜你喜欢
四川蚕业(2022年2期)2022-11-19
中华实用诊断与治疗杂志(2022年1期)2022-08-31
中华实用诊断与治疗杂志(2022年1期)2022-08-31
世界科学技术-中医药现代化(2020年2期)2020-07-25
四川蚕业(2020年3期)2020-07-16
小资CHIC!ELEGANCE(2020年4期)2020-03-09
北京青年周刊(2019年19期)2019-05-28
北京青年周刊(2019年16期)2019-05-20
中南林业科技大学学报(2019年4期)2019-04-08
森林工程(2018年1期)2018-05-14
