
小黄鱼(Larimichthys polyactis)精子的生理特性及超低温冷冻保存研究*
2020-02-06 06:01:30郑学斌王景倩谢庆平竺俊全
海洋与湖沼 2020年1期
郑学斌 杜 晨 王景倩 詹 炜 谢庆平 楼 宝① 竺俊全①
(1.宁波大学教育部应用海洋生物技术重点实验室 宁波 315211;2.浙江省农业科学院 杭州 310016)
精子活力是评价精子质量的重要指标,也是精子生理特性研究的主要内容(季相山等,2007)。鱼类精子活力受盐度、pH、离子等环境因子的影响,研究精子活力与环境因子的关系,有助于筛选人工授精用激活液及精子冷冻保存用稀释液(Islamet al,2011;Öğretmenet al,2016;王鑫伟等,2017)。目前,鱼类精子活力及其影响因子的研究已在大黄鱼(Larimichthys crocea)(朱冬发等,2005)、黄姑鱼(Nibea albiflora)(闫家强等,2010)、眼斑拟石首鱼(Csiaenopso cellatus)(魏平等,2009)、大弹涂鱼(Boleophthalmus pectinirostris)(洪万树等,1997)、夏牙鲆(Paralichthys dentatus)(王学颖等,2016)、真鲷(Pagrosomus major)(江世贵等,2000)、黄鳍鲷(Sparus latus)(黄晓荣等,2008)等海水鱼类中研究报道,研究较多的影响因子有盐度、pH、离子及葡萄糖等。鱼类精子活力检测常用传统的显微观察法,该法主观性较强,易产生人为误差,近年来,计算机辅助精子分析系统(Computerassisted sperm analysis,CASA)已在鱼类精子活力(运动率、运动速率、鞭打频率等)的检测中得到广泛应用,它可避免传统显微观测方法所造成的人为误差(Kimeet al,2001;刘清华等,2006;季相山等,2007;Kowalskiet al,2019)。
鱼类精子的超低温冷冻保存技术在人工繁殖、遗传育种及种质资源保存等方面具有重要应用价值(Cabritaet al,2010)。目前,已取得精子冷冻保存成功的海水鱼类有大黄鱼(程顺等,2013)、黄姑鱼(金春华等,2011)、黑鲷(Sparus macrocephalus)(叶霆等,2009)、大菱鲆(Scophthalmus maximus)(Chereguiniet al,2003)、鲻鱼(Mugil cephalus)(Balamuruganet al,2017)、太平洋鳕(Gadus macrocephalus)(Wanget al,2016)、褐石斑鱼(Epinephelus bruneus)(Limet al,2013)等种类。
小黄鱼(Larimichthys polyactis)属鲈形目、石首鱼科、黄鱼属,广泛分布于渤海、黄海、东海及朝鲜半岛西岸海区,曾与大黄鱼、曼氏无针乌贼(Sepia maindroni)、带鱼(Trichiurus japonicus)并称为我国“四大海产”(李建生等,2009)。近年来,小黄鱼自然资源因过度捕捞及水环境污染而衰退,并出现个体小型化、低龄化及性成熟提前等问题(林龙山等,2004;郭旭鹏等,2006;Liuet al,2017),因此,其种质资源保护、繁殖生物学、人工繁殖及增养殖技术研究引起研究者重视,如性腺发育(吴佩秋,1981;Limet al,20 10)、精卵发生(吴佩秋,1980;Kanget al,2013)、精子生理特性(Leet al,2011b)、精子冷冻保存(Leet al,2011a)、人工育苗及养殖(徐献明等,2014;陈睿毅等,2016)等方面均已见研究报道。但对人工养殖的小黄鱼精子活力及超低温冷冻保存的研究未见报道。本研究检测分析了养殖小黄鱼精子的运动特征及盐度、pH、离子、葡萄糖等环境因子的变化对其精子活力的影响;探究了稀释液、抗冻剂、稀释比及降温高度等对精子冷冻保存效果的影响,旨在了解小黄鱼精子的生理特性及超低温冷冻保存效果,为其人工繁殖及种质细胞保存提供参考。
1 材料与方法
1.1 材料
人工培育的性成熟小黄鱼取自浙江省宁波市象山港湾水产苗种有限公司,挑选成熟度好的雄鱼60尾,体长 13.7±1.13cm,体质量 35.38±7.82g。
1.2 方法
1.2.1 精液的采集 用干毛巾擦净小黄鱼泄殖孔周围体表水分,轻压鱼腹使精液自然流出,吸取无污物精液于4°C冰盒内保存,供后续实验。
1.2.2 精子密度测定 取适量精液与HBSS稀释液以1∶1000比例混匀,滴一滴于血球计数板(计数区16×25格,总体积0.1mm3)上,加盖玻片后于显微镜下观察并测定精子密度(n=3)。实验重复3次,下同。
1.2.3 精子运动特征检测 取0.5μL精液,加入500μL自然海水激活,并用计算机辅助精子分析系统(CASA)测定激活20s后精子的运动率(percentage of motile,MOT,运动精子占所有精子的百分率)、直线运动速率(straight line velocity,VSL)、曲线运动速率(curvilinear velocity,VCL)、平均路径运动速率(average path velocity,VAP)、侧摆幅值(amplitude of lateral head displacement,ALH)、鞭打频率(beating cross frequency,BCF)、平均移动角度(mean angular displacement,MAD)等指标(视野数3,捕捉精子数≥200),并人工记录精子的运动时间(moving time,MT)及寿命(life span,LS)(精子激活开始至90%精子原地颤动的时间为运动时间,90%精子停止运动的时间为寿命)。
1.2.4 盐度对精子活力的影响实验 在过滤海水(盐度27,pH 8.1)中添加蒸馏水或海水晶,配制成盐度为 5、10、15、20、25、30、35、40的实验溶液,检测精子在各实验溶液中的活力(运动率,运动时间及寿命,下同)。
1.2.5 pH对精子活力的影响实验 在过滤海水(盐度27,pH 8.1)中滴加1mol/L的HCl或NaOH,配制成 pH 为 5.0、6.0、7.0、7.5、8.0、8.5、9.0、10.0、11.0的实验溶液,检测精子在各实验溶液中活力。
1.2.6 葡萄糖对精子活力的影响实验 在去离子水中添加葡萄糖,配制成浓度梯度为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0mol/L的实验溶液,检测精子在各实验溶液中的活力。
1.2.7 KCl、NaCl、MgCl2及CaCl2溶液对精子活力的影响实验 在去离子水中添加NaCl、KCl、MgCl2及CaCl2,配制成浓度梯度为0.1、0.2、0.3、0.4、0.5、0.6、0.7、0.8、0.9、1.0mol/L的实验溶液,检测精子在各实验溶液中的活力。
1.2.8 不同海水对精子活力的影响实验 参考魏平等(2009)方法配制全人工海水(每1000mL人工海水中含 19.29g NaCl、4.24g MgSO4、0.84g CaCl2、0.51gKCl、0.12g NaHCO3),并用相同摩尔数的MgCl2、Na2SO4和NaCl依次替换 ASW中的 CaCl2、MgSO4和NaHCO3,配制成缺Ca2+、缺Mg2+和缺HCO3-的人工海水,检测精子在各实验溶液中的活力。
1.2.9 稀释液种类对冷冻保存效果的影响实验 分别用 MFRS、ASP、Hank’s、MPRS、HBSS、Cortland为稀释液(各稀释液成分见表1),稀释比1∶3,并添加10%二甲基亚砜(dimethyl sulfoxide,DMSO)(终浓度,下同)为抗冻剂,0.25mL麦细管为冻存管,用二步降温法于液氮面上3.5cm处平衡5min后投入液氮中保存,12h后取出,40°C水浴解冻6s后测定精子活力。

表1 小黄鱼精子超低温冷冻保存稀释液的组成成分Tab.1 Composition of extender for cryopreservation of L.polyactis sperm
1.2.10 抗冻剂种类及浓度对冷冻保存效果的影响实验 以HBSS为稀释液,稀释比1∶3,分别以5%、10%、15%及20%的DMSO、乙二醇(ethylene glycol,EG)、丙二醇(propyleneglycol,PG)、甘油(glycerol,Gly)作为抗冻剂,0.25mL麦细管为冻存管,用二步降温法于液氮面上3.5cm处平衡5min后投入液氮中保存,12h后取出,40°C水浴解冻6s后测定精子活力。
1.2.11 稀释比对冷冻保存效果的影响实验 以HBSS为稀释液,分别以1∶1,1∶3,1∶5,1∶7,1∶9稀释精液,添加10%DMSO为抗冻剂,0.25mL麦细管为冻存管,二步降温法于液氮面上3.5cm处平衡5min后投入液氮中保存,12h后取出,40°C水浴解冻6s后测定精子活力。
1.2.12 降温高度对冷冻保存效果的影响实验 以HBSS为稀释液,稀释比1∶3,并添加10%DMSO为抗冻剂,0.25mL麦细管为冻存管,用二步降温法分别于液氮面上2、3.5、5、7.5、10cm处平衡5min后投入液氮,12h后取出,40°C水浴解冻6s后测定精子活力。
1.3 数据处理
数据统计与分析采用Excel和SPSS19.0软件完成,用Excel进行作图,用单因素方差分析(One-Way ANOVA)进行显著性分析(SPSS19.0)。统计结果以平均值±标准差(Mean±SD)表示。
2 结果
2.1 小黄鱼精子密度
经检测,小黄鱼精液中精子密度为(6.92±1.20)×109/mL。
2.2 小黄鱼精子运动特征
小黄鱼精子经自然海水激活后各运动参数见表2(n=10),曲线运动速率和平均路径运动速率较高,直线运动速率较低。

表2 小黄鱼精子的运动参数(n=10)Tab.2 The parameters of sperm movement of L.polyactis(n=10)
2.3 盐度对小黄鱼精子活力的影响
水温16°C、pH8.1,小黄鱼精子在不同盐度溶液中的活力(运动率、运动时间、寿命)见图1。精子在盐度10—40溶液中均有一定比例被激活,在盐度25—30溶液中,精子运动率大于45%,在盐度25溶液中,精子运动率、运动时间及寿命分别达(53.43±5.08)%、8.17±0.41min及11.37±0.86min,与对照组无显著差异。精子在盐度为5的溶液中未被激活。
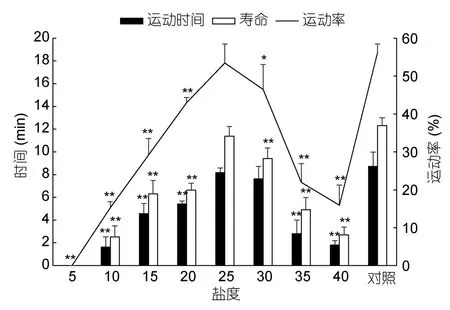
图1 盐度对小黄鱼精子活力的影响Fig.1 Effects of salinity on the motility of L.polyactis sperm
2.4 pH对小黄鱼精子活力的影响
水温16°C、盐度27,小黄鱼精子在不同pH溶液中的活力见图2。精子在pH 5.0—11.0的实验溶液中均有一定比例被激活,在pH 7.5—8.5溶液中,精子运动率大于50%,在pH 8.0溶液中,精子运动率、运动时间及寿命分别达(59.12±8.62)%、8.32±0.33min及11.64±0.74min,与对照组无显著差异。

图2 pH对小黄鱼精子活力的影响Fig.2 Effects of pH value on the motility of L.polyactis sperm
2.5 葡萄糖溶液对小黄鱼精子活力的影响
水温16°C,小黄鱼精子在葡萄糖溶液中的活力见图3。在0.3—1.0mol/L葡萄糖溶液中,均有部分精子被激活,在0.7—0.9mol/L溶液中,精子运动率与对照组无显著差异,在0.8mol/L溶液中,精子运动率、运动时间及寿命分别达(56.65±5.01)%、8.51±0.46min及12.19±0.67min。在0.1或0.2mol/L葡萄糖溶液中,精子未被激活。

图3 葡萄糖溶液对小黄鱼精子活力的影响Fig.3 Effects of glucose solutions on the motility of L.polyactis sperm
2.6 KCl、NaCl、MgCl2及 CaCl2溶液对小黄鱼精子活力的影响
水温 16°C,小黄鱼精子在 KCl、NaCl、MgCl2及CaCl2溶液中的活力见表3、表4、表5。精子在0.1—0.9mol/L的KCl溶液中均有部分被激活,在0.4或0.5mol/L KCl溶液中,精子的运动率相对较高(约45%),但均显著低于对照组;在0.1mol/L KCl溶液中,精子未被激活。精子在0.2—1.0mol/L的NaCl溶液中均有一定比例被激活,在0.4—0.6mol/L NaCl溶液中,精子活力较好,在0.5mol/L NaCl溶液中,精子运动率、运动时间及寿命分别达(53.80±12.17)%、7.79±1.45min及10.77±0.88min,与对照组差异不显著;在0.1mol/L NaCl溶液中,精子未被激活。精子在0.3—1.0mol/L MgCl2溶液中均有一定比例被激活,在0.7—1.0mol/LMgCl2溶液中,精子运动率大于40%,在0.9mol/LMgCl2溶液中,精子运动率、运动时间及寿命分别达(58.39±1.83)%、8.04±0.32min 及 11.71±0.54min,与对照组差异不显著;在0.1—0.2mol/L MgCl2溶液中,精子未被激活。精子在0.1—0.8mol/L CaCl2溶液中均有部分被激活,在0.2—0.3mol/L CaCl2溶液中,精子运动率,运动时间及寿命相对较高,但均显著低于对照组;精子在0.9—1.0mol/L CaCl2溶液中未被激活。
2.7 不同海水对小黄鱼精子活力的影响
水温16°C,小黄鱼精子在各人工海水及自然海水中的活力见表6。小黄鱼精子在自然海水中的运动率显著高于各人工海水,运动时间及寿命也比各人工海水更长;而在各离子缺陷型人工海水中,精子的运动率与全人工海水相比无显著差异。
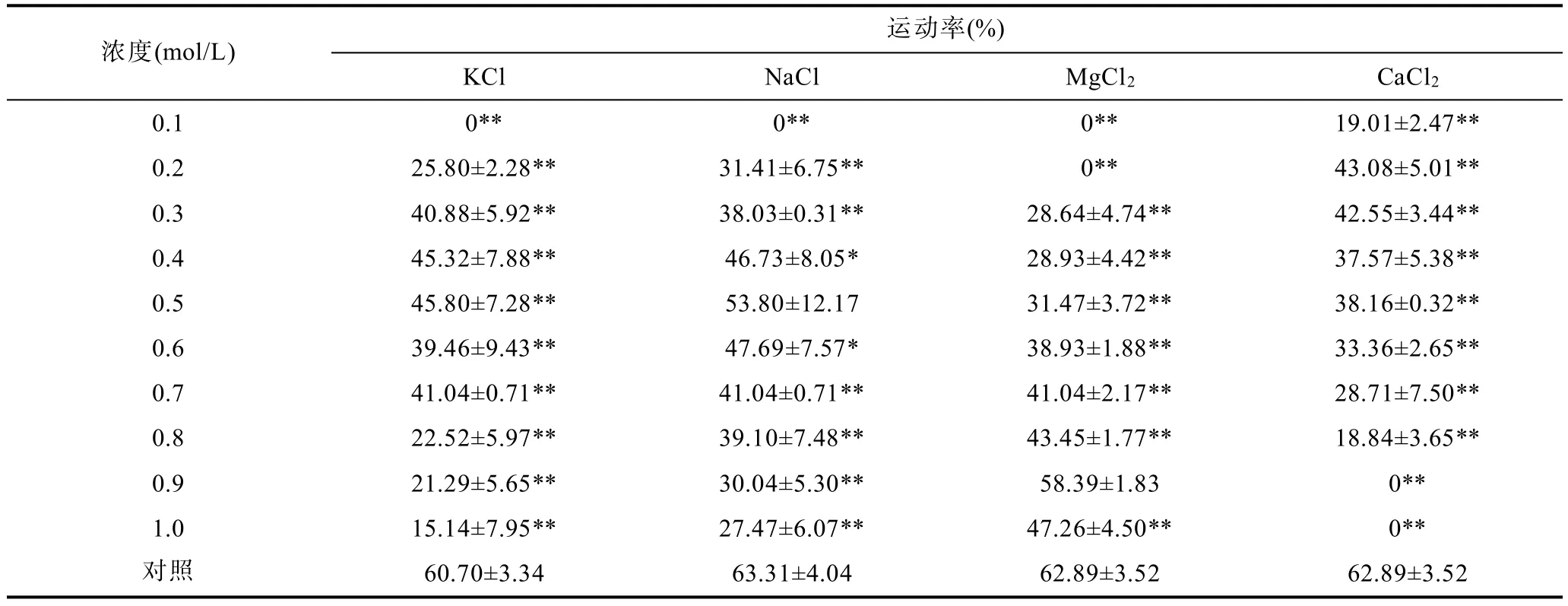
表3 KCl、NaCl、MgCl2、CaCl2溶液对小黄鱼精子运动率的影响Tab.3 Effects of KCl,NaCl,MgCl2,CaCl2solutions on the percentage of L.polyactis sperm motile

表4 KCl、NaCl、MgCl2、CaCl2溶液对小黄鱼精子运动时间的影响Tab.4 Effects of KCl,NaCl,MgCl2,CaCl2solutions on the moving time of L.polyactis sperm
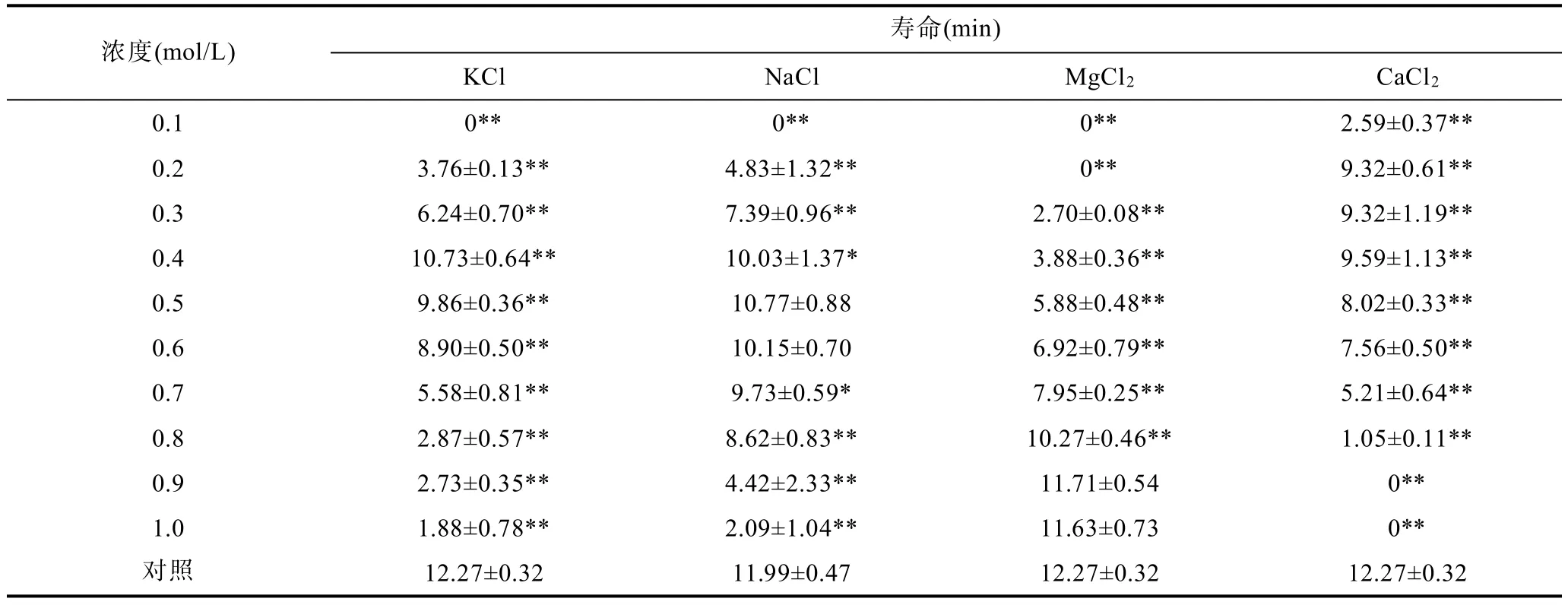
表5 KCl、NaCl、MgCl2、CaCl2溶液对小黄鱼精子寿命的影响Tab.5 Effects of KCl,NaCl,MgCl2,CaCl2solutions on the life span of L.polyactis sperm

表6 不同海水对小黄鱼精子活力的影响Tab.6 Effects of different seawater on the motility of L.polyactis sperm
2.8 稀释液种类对精子冷冻保存效果的影响
以MFRS等6种溶液为稀释液,10%DMSO为抗冻剂冷冻保存小黄鱼精子,结果显示,精子在HBSS稀释液中冷冻保存效果最好,冻精运动率为(41.38±5.03)%,其次是 MFRS,运动率(39.56±6.08)%,在ASP、Hank’s、Cortland及MPRS稀释液中冷冻保存效果较差。对照组鲜精运动率为(59.93±3.41)%,显著高于各冻精组(图4)。
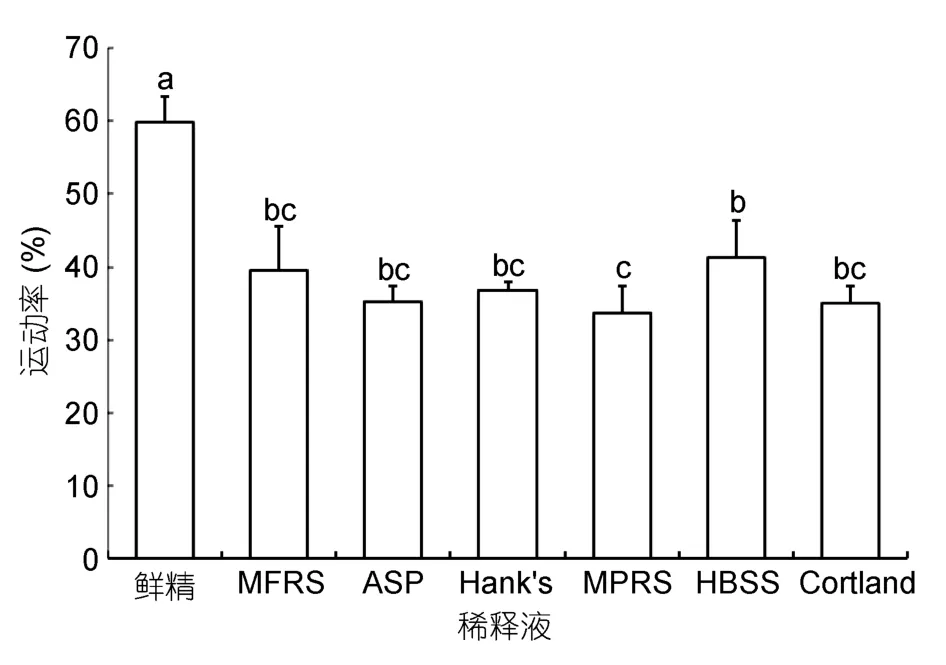
图4 稀释液种类对小黄鱼精子冷冻保存效果的影响Fig.4 Effects of different extenders on cryopreservation of L.polyactis sperm
2.9 抗冻剂种类及浓度对小黄鱼精子冷冻保存效果的影响
以HBSS为稀释液,不同浓度的四种抗冻剂对精子冷冻保存效果的影响见图5。10%DMSO及15%PG对精子冷冻保存效果较好,运动率分别为(43.57±2.59)%和(43.06±6.77)%,5%、10%EG,5%、15%、20%DMSO及10%、15%、20%PG保存效果次之,而不同浓度Gly保存效果较差,运动率显著低于10%DMSO及15%PG。对照组鲜精运动率为(59.27±1.38)%,显著高于各冻精组(图5)。
2.10 稀释比对小黄鱼精子冷冻保存效果的影响
以HBSS为稀释液,10%DMSO为抗冻剂,不同稀释比冷冻保存小黄鱼精子,结果显示,精子经1∶3或1∶5稀释后冷冻保存效果较好,运动率分别为(43.59±1.59)%和(42.83±3.72)%,其次为 1∶7,而 1∶1及1∶9冷冻保存效果较差。对照组鲜精运动率为(57.55±2.44)%,显著高于各冻精组(图6)。

图5 抗冻剂种类及浓度对精子冷冻保存效果的影响Fig.5 Effects of different cryoprotectants on cryopreservation of L.polyactis sperm

图6 稀释比对小黄鱼精子冷冻保存效果的影响Fig 6 Effects of milt/diluent ratios on cryopreservation of L.polyactis sperm
2.11 降温高度对精子冷冻保存效果的影响
以HBSS为稀释液,10%DMSO为抗冻剂,不同液氮面高度降温5min后冷冻保存小黄鱼精子,结果显示,液氮面3.5cm处降温5min后冷冻保存效果较好,冻精运动率为(45.67±0.95)%,其次为2cm,而5、7.5或10cm保存效果较差。对照组鲜精运动率为(57.70±2.72)%,显著高于各冻精组(图7)。

图7 降温高度对小黄鱼精子冷冻保存效果的影响Fig 7 Effects of the height above surface of liquid nitrogen on cryopreservation of L.polyactis sperm
3 讨论
3.1 精子的运动特征
鱼类精子的运动特征因种的不同存在差异,现将部分鱼类精子的运动参数列于表7,对其进行比较。由表7可知,鱼类精子的运动速率一般介于几十至几百微米每秒,如高体雅罗鱼(Siddiqueet al,2016)精子的运动速率约为30μm/s,条纹锯鮨(韩龙江等,2014)精子的运动速率约为80μm/s,而鲤鱼(Cejkoet al,2015)精子的运动速率较高,可达250μm/s以上。本研究中,小黄鱼精子的VSL、VCL及VAP分别为11.96±5.21、25.38±7.19及 22.33±6.88μm/s,低于大西洋鳕(Buttset al,2010),欧洲鳗鲡(Gallegoet al,2013)及条纹锯鮨(Gallegoet al,2013)等海水鱼类,而与淡水的高体雅罗鱼(Siddiqueet al,2016)相近。研究表明,养殖大西洋鳕精子的运动速率要显著低于野生大西洋鳕,且两种环境条件下,大西洋鳕精子的运动速率均随繁殖时间的推移呈逐渐上升趋势(Buttset al,2010),小黄鱼精子是否存在此类现象,值得进一步研究。鱼类精子的运动轨迹常为光滑的曲线,因此精子VCL和VAP的值基本一致(Rurangwaet al,2004)。表7中,鲤鱼(Cejkoet al,2015)、高体雅罗鱼(Siddiqueet al,2016)、西伯利亚鲟(Siddiqueet al,2016)、大西洋鳕(Buttset al,2010)等鱼的运动速率均以VCL值最大,且VCL和VAP值较接近(表7),表明这些鱼类精子多呈曲线式运动。本研究中,小黄鱼精子VCL和VAP值也较接近,且VSL较小,表明小黄鱼精子的运动方式也是以曲线运动为主。
3.2 环境因子对精子活力的影响
盐度是影响海水鱼类精子活力的主要因子,且精子适宜的盐度范围往往与其生境或繁殖环境相关(江世贵等,2000)。据研究,眼斑拟石首鱼(魏平等,2009)、黄姑鱼(闫家强等,2010)及大黄鱼(朱冬发等,2005)精子运动适宜的盐度分别为20—35、25—35及 19.61—24.87,而大弹涂鱼(洪万树等,1997)精子运动适宜的盐度范围较低(15—20)。本研究中,小黄鱼精子运动适宜的盐度为25—30,与黄姑鱼(闫家强等,2010)及眼斑拟石首鱼(C.ocellatus)(魏平等,2009)精子运动适宜的盐度相似,小黄鱼繁殖期的海区盐度一般为27—32,是其精子运动适宜的盐度范围。

表7 部分鱼类精子运动特征比较Tab.7 Comparison in kinetic characteristics of sperm in some fish species
pH对鱼类精子活力影响也较大。据报道,鱼类精浆pH大多介于7.3—8.5之间,多数鱼类精子在中性或偏碱性的水体中激活效果更好(邓岳松等,1999;Kowalskiet al,2019)。据研究,大黄鱼(朱冬发等,2005)精子运动适宜的pH范围是7.5—8.0,精子在pH 7.5溶液中运动率最高、运动时间最长;黄姑鱼(闫家强等,2010)精子运动适宜的pH范围是7.5—8.5,在pH 8.0时,精子活力最好;此外,真鲷(江世贵等,2000)、黄鳍鲷(S.latus)(黄晓荣等,2008)、欧洲鲈鱼(Dicentrarchus labrax)(Öğretmenet al,2016)等海水鱼类精子也都在中性偏碱的激活液中活力较高。本研究,小黄鱼精子运动适宜的pH范围是7.5—8.5,pH为8.0时,精子运动率、运动时间及寿命分别达(59.12±8.62)%、8.32±0.33min及 11.64±0.74min,表明小黄鱼精子也是在偏碱性的环境中活力较好,因此,在配制小黄鱼人工授精激活液或精子冷冻保存抗冻液时,应首选pH偏碱性(8.0左右)的配方。
据报道,适宜浓度的葡萄糖溶液对江鳕(Lota lota)(王位莹等,2015)、高体雅罗鱼(张涛等,2017)、中华鲟(Acipenser sinensis)(刘鉴毅等,2007)及乌原鲤(Procypris mterus)(吴清毅等,2011)等淡水鱼类精子活力有促进作用,可延长精子的运动时间或寿命;葡萄糖溶液对眼斑拟石首鱼(魏平等,2009)、黄姑鱼(闫家强等,2010)、大黄鱼(朱冬发等,2005)等海水鱼类精子活力无明显促进作用,而对夏牙鲆(王学颖等,2016)精子活力有一定提升。本研究中,小黄鱼精子在0.7—0.9mol/L葡萄糖溶液中保持较高活力,在0.8mol/L时,精子运动率、运动时间及寿命分别达(56.65±5.01)%、8.51±0.46min及 12.19±0.67min,但与对照组各活力参数基本持平,表明葡萄糖对小黄鱼精子活力无明显促进作用。
K+、Na+、Ca2+及Mg2+是激活鱼类精子的重要因子,也是精浆中的主要离子成分(Alaviet al,2006;Figueroaet al,2016),不同种鱼类精子对各种离子的敏感性不同(Alaviet al,2006),可能会导致其激活适宜的离子浓度有所差异。据研究,大弹涂鱼(魏平等,2010)精子激活效果较好的K+、Na+、Ca2+及Mg2+浓度分别为0.3、0.15、0.1、0.3mol/L,眼斑拟石首鱼(魏平等,2009)精子激活效果较好的K+、Na+、Ca2+浓度均为0.6mol/L,而大黄鱼(朱冬发等,2005)精子激活效果较好的K+、Na+浓度分别为0.5和0.3mol/L,本研究小黄鱼精子激活效果较好的K+、Na+、Ca2+及Mg2+浓度分别为0.4、0.5、0.2、0.9mol/L,与上述几种鱼类有所不同。
本研究显示,小黄鱼精子在不同离子激活液中的运动率均低于自然海水,这表明单一金属阳离子对小黄鱼精子的激活效果要低于含复合金属阳离子的自然海水;小黄鱼精子在单一离子缺陷型人工海水和全人工海水中的运动率非常接近(介于48%—53%之间),表明单一离子缺失的人工海水并没有对其精子活力造成明显的影响。王学颖等(2016)研究发现夏牙鲆精子在自然海水中的运动率要显著高于各人工海水,自然海水的渗透压为939mosm/kg,而缺Ca2+、缺Mg2+、缺HCO3-人工海水或全人工海水的渗透压介于1029—1131mosm/kg之间,因此渗透压可能是导致精子运动率不同的因素之一。本研究中的小黄鱼精子在自然海水中的运动率显著高于各人工海水,这可能是自然海水与人工海水的离子成分不同及渗透压不同所致。
3.3 精子超低温保存的适宜条件
不同种鱼类精子超低温冷冻保存适宜的条件(稀释液、稀释比、抗冻剂种类及浓度等)不同(王鑫伟等,2017)。表8是部分海水鱼类精子冷冻保存条件及其效果比较。
不同规格的麦管(0.25、0.5mL)和冻存管(2、5mL)在鱼类精子冷冻保存研究中均有应用,麦管具有体积小、降温速率快、稳定性高等特点,适用于精液量较少的冷冻保存研究(表8)。而冻存管体积较麦管大、降温速率相对较慢,适用于精液量较多的冷冻保存研究(Liuet al,2015;Nomuraet al,2018)。本研究的养殖小黄鱼雄鱼个体较小、精液量较少,因此使用0.25ml麦管进行了冷冻保存研究,以确定其适宜的冷冻条件。今后,若培育较大规格的小黄鱼雄鱼,精液量多时,可采用体积较大的冻存管进行精子冷冻保存研究,以便于冷冻保存大量精液。
不同种鱼类精子冷冻保存用适宜的稀释液及其稀释比不同。Fabbrocini等(2000)和Gwo等(1991)用1%NaCl溶液为稀释液,分别以1∶6和1∶9的稀释比冷冻保存金头鲷及细须石首鱼精子效果较好;Chereguini等(2003)用 Mounib’s稀释液,以 1∶2 的稀释比冷冻保存大菱鲆精子,获得与鲜精相近的受精率;Degraaf等(2004)调整了 Mounib’s稀释液成分,并以1∶3的稀释比冷冻保存条纹锯鮨精子,获得与鲜精相近的运动率和受精率。此外,HBSS、Hank’s、Ranger、Cortland等稀释液在庸鲽、夏牙鲆、紫红笛鲷、大黄鱼等海水鱼类精子冷冻保存研究中获得较好的效果(表8)。本研究,以10%DMSO为抗冻剂,以HBSS为稀释液、1∶3或1∶5的稀释比,超低温冷冻保存小黄鱼精子,获得了较好的运动率,但与鲜精运动率相比仍有一定差距。

?

?
超低温冷冻过程会导致精子结构损伤,适宜浓度的抗冻剂可有效保护精子结构(陈松林等,1992;Kowalskiet al,2019)。DMSO、Gly、EG、PG等是鱼类精子冷冻保存常用的抗冻剂,其中DMSO具有渗透速度快、分布均匀、毒性低等特点(李纯等,2000),在庸鲽、金头鲷、细须石首鱼等海水鱼类精子冷冻保存中起到较好的抗冻保护效果(表8)。抗冻剂适宜的使用浓度一般在5%—20%之间(表8)。本研究的小黄鱼精子冷冻保存采用10%DMSO及15%PG为抗冻剂,冻精解冻后运动率分别为(43.57±2.59)%和(43.06±6.77)%,与鲜精运动率(57.70±2.72)%较为接近,表明10%DMSO和15%PG适用于小黄鱼精子超低温冷冻保存。
精子冷冻保存的降温方法主要有液氮面高度降温法或程序降温法,两者使用的仪器设备和操作方法有所不同,但都是控制降温速率。由表8可知,使用程序降温法超低温冻存鱼类精子,其适宜的降温速率因鱼的种类不同存在差异,但多数在10—20°C/min;少数鱼如绿鳍马面鲀降温速率较高(40°C/min)。使用液氮面高度降温法超低温冻存鱼类精子,多数在液氮面3—5cm处降温3—5min后投入液氮保存。液氮面3.5cm处的温度约-76°C(Limet al,2013),如果在此高度降温5min,平均降温速率为15.2°C/min。本研究的小黄鱼精子冷冻保存也是在液氮面3.5cm处平衡5min后投入液氮,其降温速率在15°C/min左右。今后,应开展程序降温法超低温冻存小黄鱼精子试验,以确定其最佳降温速率,提高冻存效果。
4 结论
小黄鱼精子主要以曲线运动为主,精子适宜的盐度为25—30,适宜的pH为7.5—8.5,适宜浓度的葡萄糖溶液对精子活力无明显促进作用。小黄鱼精子在KCl和CaCl2溶液中受到一定程度抑制,活力明显低于对照组。单一离子缺陷型人工海水对小黄鱼精子活力影响不大,但激活效果不及自然海水。小黄鱼精子经HBSS稀释液1∶3稀释,添加10%DMSO及15%PG为抗冻剂,液氮面3.5cm处降温5min后投入液氮保存效果较好,冻精运动率与鲜精较接近。
猜你喜欢
快乐作文(5.6年级)(2023年4期)2023-05-04 23:23:16
今日农业(2022年1期)2022-06-01 06:18:10
现代畜牧科技(2021年11期)2021-12-21 06:11:08
小读者(2020年4期)2020-06-16 03:34:08
今日农业(2020年24期)2020-03-17 08:58:00
海峡姐妹(2020年1期)2020-03-03 13:36:02
水利技术监督(2017年2期)2017-05-17 05:19:34
食品与生活(2016年10期)2016-10-13 18:57:23
中国畜牧兽医文摘(2015年9期)2015-12-29 03:38:09
莫愁·家教与成才(2015年8期)2015-05-30 10:48:04
