
干旱和盐胁迫对不同种源地麻花艽幼苗生长和生理特性的影响
2020-02-03 03:16程庭峰陈世龙王久利师生波沈建伟周党卫
干旱地区农业研究 2020年6期
程庭峰,王 环,陈世龙,王久利,师生波,沈建伟,周党卫,5
(1.中国科学院西北高原生物研究所高原生物适应与进化重点实验室,青海 西宁 810008;2.中国科学院西北高原生物研究所分析测试中心,青海 西宁 810008;3.青海省作物分子育种重点实验室,青海 西宁 810008;4.青海民族大学生态环境与资源学院,青海 西宁 810007;5.中国科学院大学,北京 100049)
麻花艽(GentianastramineaMaxim.) 为龙胆属多年生草本植物[1],《中国药典》2015 年版将其和同为秦艽组(Sect.CruciataGaudi)的大叶秦艽 (G.macrophyllaPall.)、粗茎秦艽 (G.crassicaulisDuthie ex Burk.)、小秦艽 (G.dahuricaFisch.)一起列为中药秦艽的4种基源植物,秦艽是一种传统的中藏药材,距今已有2000多年的历史,具有祛风湿、清湿热、止痹痛和退虚热等功效[2]。研究发现生长在不同环境下的植物,经过自然选择和适应,能够在形态和生理特性等方面产生变异,最终形成特定的地理种源;麻花艽在我国主要分布于青藏高原及其周边地区,推测不同种源麻花艽在长期的生长过程中也已经形成了适应多种生境的遗传差异[3-4]。近年来,随着市场需求量的增加,麻花艽野生资源遭到严重破坏,因此有必要开展麻花艽的人工栽培研究[5]。
盐胁迫和干旱胁迫是植物最容易遭受的逆境胁迫,能够通过渗透胁迫造成植物损伤,导致植物体内活性氧积累,是植物生长的限制性因素之一。我国存在大量的干旱盐渍土地,其中又以西北地区(青海、甘肃、新疆)为主[6-8]。当前麻花艽人工种植在青海等地已经广泛开展,因此,对胁迫下不同种源麻花艽生理特性的研究,将有助于抗旱耐盐种质资源的筛选和培育,保证野生资源的可持续开发利用[9]。
目前,相关学者已经对麻花艽、秦艽等植物胁迫下的萌发特性进行了研究,但胁迫对其生理特性的影响却鲜有报道[10-11]。本试验以不同采集地点的麻花艽种子为研究对象,分别通过PEG6000、NaCl以及两者混合模拟干旱胁迫、盐胁迫和混合胁迫,分析不同胁迫对不同种源麻花艽幼苗的形态、可溶性蛋白、丙二醇以及抗氧化酶活性的影响差异,阐明干旱和盐胁迫对麻花艽生理特性的影响,为其驯化栽培提供理论依据。
1 材料和方法
1.1 材料与处理
麻花艽种子于2018年分别取材自青海省海北站(HBZ)、德令哈(DLH)、河北乡(HBX)、下藏科(XZK)和班玛县(BMX)5个区域。晾干后选取籽粒饱满、大小均匀、成熟的种子作为实验材料。
将麻花艽种子置于300 μmol·L-1的赤霉素中浸泡24 h,分别用无菌水漂洗3次,75%的酒精消毒1 min、2.5%的NaClO溶液消毒10 min后,再用无菌水漂洗5次。将消毒后的种子接入30 mL MS固体培养基中,所有的培养瓶放到人工气候培养箱中进行培养。大约3周后植株长至 2 cm 时,分别接入5 mL 10% PEG、150 mmol·L-1NaCl以及10% PEG+150 mmol·L-1NaCl混合溶液模拟干旱胁迫、盐胁迫和混合胁迫,7 d后将材料取出,检测麻花艽幼苗的株高、根长、鲜重以及鲜重下的可溶性蛋白和丙二醛含量、抗氧化酶活性等生理指标。
1.2 各生理指标的测定及其方法
可溶性蛋白含量测定采用考马斯亮蓝 G-250染色法;丙二醇(MDA)测定采用硫代巴比妥酸(TBA)比色法;超氧化物歧化酶(SOD)活性采用氮蓝四唑(NBT)法测定;过氧化物酶(POD)活性采用愈创木酚比色法测定;过氧化氢酶(CAT)活性采用紫外吸收法测定。
1.3 数据分析
实验数据通过SPSS 20.0进行统计分析,采用Excel 2010进行绘图。生长指标以 3次重复的平均值及其标准误差表示。利用相对可溶性蛋白、相对MDA和相对抗氧化酶活性等指标,通过隶属函数法对幼苗抗性进行综合评价。
隶属函数值的计算公式:
R(Xi) = (Xi-Xmin)/(Xmax-Xmin)
反隶属函数值计算公式:
R(Xi) = 1-(Xi-Xmin)/(Xmax-Xmin)
式中,Xi为实验材料在某一胁迫下的测定值;Xmax和Xmin分别为该胁迫下所有材料中该指标的最大值和最小值,计算完所有指标的隶属值后,通过累加求平均,平均值越大,表明其抗性越强[12-13]。
2 结果与分析
2.1 不同胁迫对麻花艽幼苗生长的影响
双因素方差分析结果显示,采集地、胁迫及两者的交互显著影响麻花艽的生长(表1)。
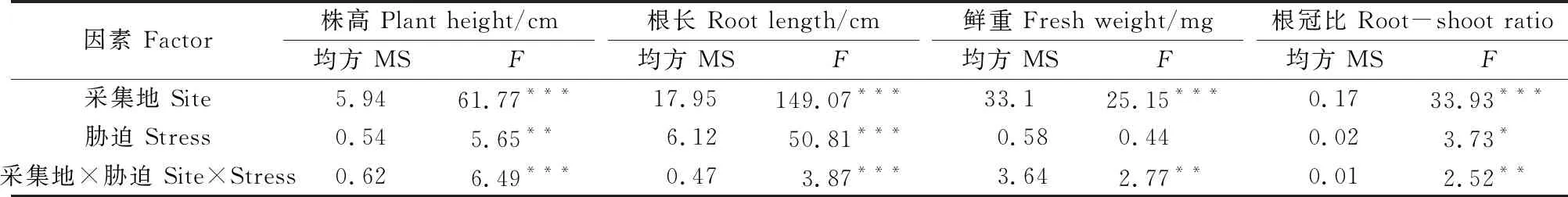
表1 麻花艽幼苗生长的双因素方差分析结果
不同胁迫对麻花艽幼苗的生长具有不同影响,如表2所示。与对照相比,除河北乡和盐胁迫下的海北站外,3种胁迫下幼苗的株高都出现降低,且混合胁迫下抑制作用更明显;所有材料的根长在胁迫下都出现降低;河北乡幼苗的鲜重在干旱胁迫下出现显著增加(P<0.05),其余地区材料有变化但无显著性差异;海北站幼苗的根冠比在干旱胁迫下出现显著降低(P<0.05),其余材料出现变化但与对照间也无显著性差异。
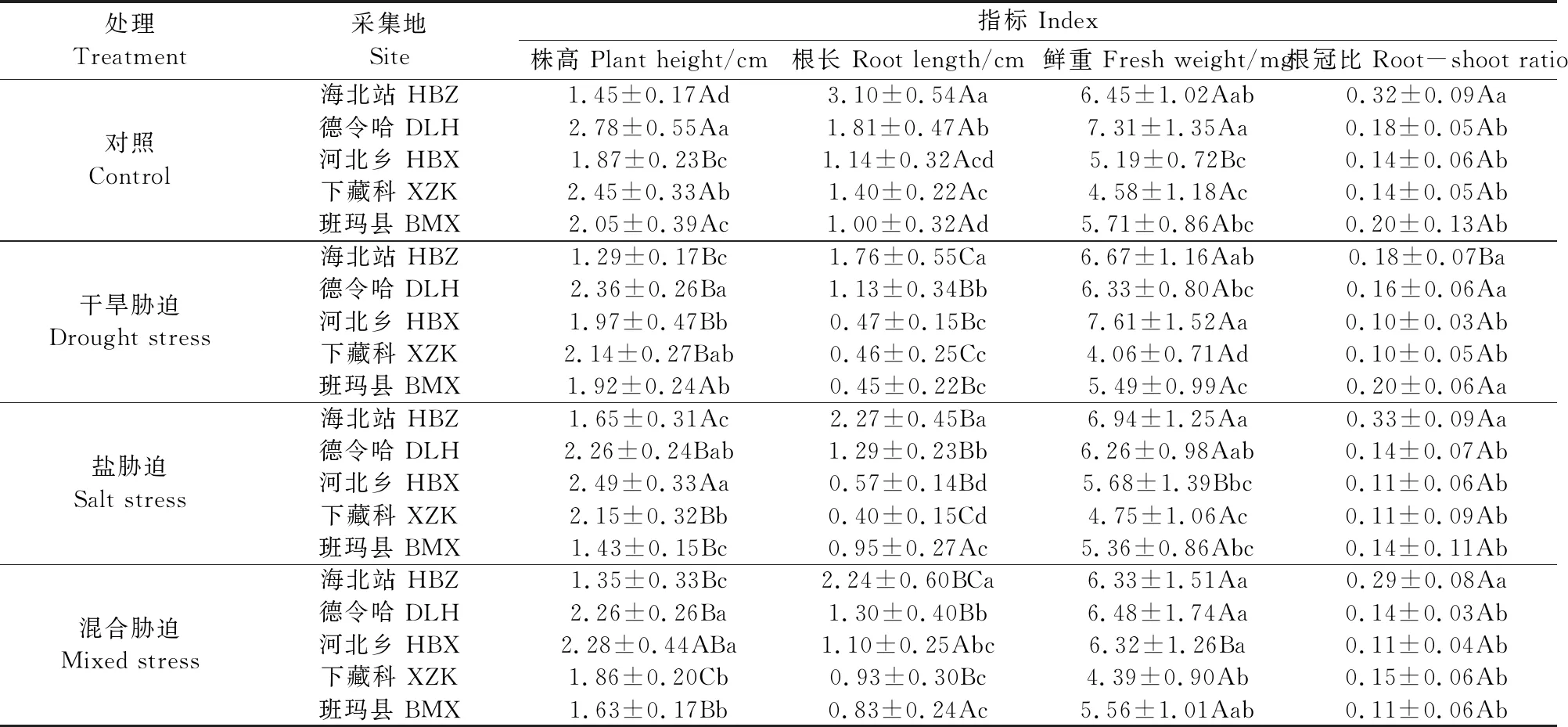
表2 胁迫对不同种源地麻花艽幼苗生长的影响
不同种源麻花艽种子对胁迫的响应也具有一定的差异,干旱胁迫下德令哈地区幼苗的株高最高,为2.36 cm;盐胁迫和混合胁迫下河北乡和德令哈地区幼苗的株高均显著高于其他地区(P<0.05),且河北乡均为最高,分别为2.49 cm和2.28 cm。3种胁迫条件下,海北站根长最长。干旱胁迫下河北乡地区幼苗鲜重最大,为7.61 mg;盐胁迫下海北站鲜重最大,为6.94 mg;混合胁迫下德令哈鲜重最高,为6.48 mg。干旱胁迫下班玛县幼苗根冠比最大,为0.20;盐和混合胁迫下海北站根冠比最大,分别为0.33和0.29。
2.2 不同胁迫对麻花艽幼苗丙二醇含量的影响
如图 1所示,与对照相比,除干旱胁迫下的德令哈外,其他采集地麻花艽幼苗的MDA含量均出现增加,3种胁迫条件下班玛县MDA增加量均为最高,分别增加了1 509.75%,605.29%和1 175.76%。其中海北站、德令哈和河北乡幼苗的MDA含量在混合胁迫下增加更为明显,分别增加了833.92%,23.93%和242.05%,下藏科幼苗在盐胁迫下增加最为明显,增加了600.29%。

图1 胁迫对麻花艽幼苗丙二醇含量的影响
2.3 不同胁迫对麻花艽幼苗可溶性蛋白含量的影响
如图 2所示,不同地理种源麻花艽幼苗可溶性蛋白的含量在胁迫下呈现出不同的变化趋势。与对照相比,干旱胁迫下海北站、德令哈和下藏科均显著增加,分别增加了23.87%,46.52%和10.00%;盐胁迫下德令哈的幼苗可溶性蛋白增加了12.15%,其余地区都出现降低,降幅不显著;混合胁迫下所有材料可溶性蛋白都出现降低,其中海北站、德令哈和河北乡显著下降,分别下降了21.96%,18.32%和12.80%。
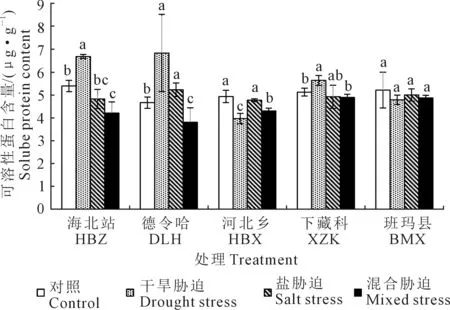
图2 胁迫对麻花艽幼苗可溶性蛋白含量的影响
2.4 不同胁迫对麻花艽幼苗抗氧化酶活性的影响
如图3所示,胁迫下不同地理种源麻花艽幼苗的抗氧化酶活性也出现不同变化。与对照相比,干旱胁迫下所有材料的SOD均出现降低,盐胁迫下仅下藏科增加了17.23%,混合胁迫下仅德令哈地区增加了6.45%,其余地区均表现为降低(图3A)。胁迫下POD活性也出现不同变化,干旱胁迫下除河北乡外都出现增加,其中海北站增加最明显,相较于对照增加了234.39%;盐胁迫下除海北站增加了340.53%外,其余地区POD活性都出现下降;混合胁迫下除下藏科增加了88.18%倍外,其余地区也都出现下降(图3B)。胁迫下多数地区幼苗的CAT活性出现降低,干旱和盐胁迫下仅海北站分别增加了86.39%和127.86%,其余地区都出现下降;混合胁迫下海北站、德令哈和下藏科均出现显著性增加,分别增加了12.83%、51.20%和26.57%(图3C)。

图3 胁迫对麻花艽幼苗SOD、POD和CAT活性的影响
2.5 不同种源麻花艽幼苗的综合抗性评价
不同地理种源麻花艽的抗性隶属函数分析如表3所示,其中相对可溶性蛋白、相对抗氧化酶活性计算采用隶属函数公式,MDA与幼苗生长表现为负相关,其计算采用反隶属函数公式。抗旱性综合评价顺序为:德令哈>海北站>下藏科>河北乡>班玛县,耐盐性顺序为:海北站>下藏科>德令哈>班玛县>河北乡,混合胁迫抗性顺序为:德令哈>下藏科>河北乡>海北站>班玛县。
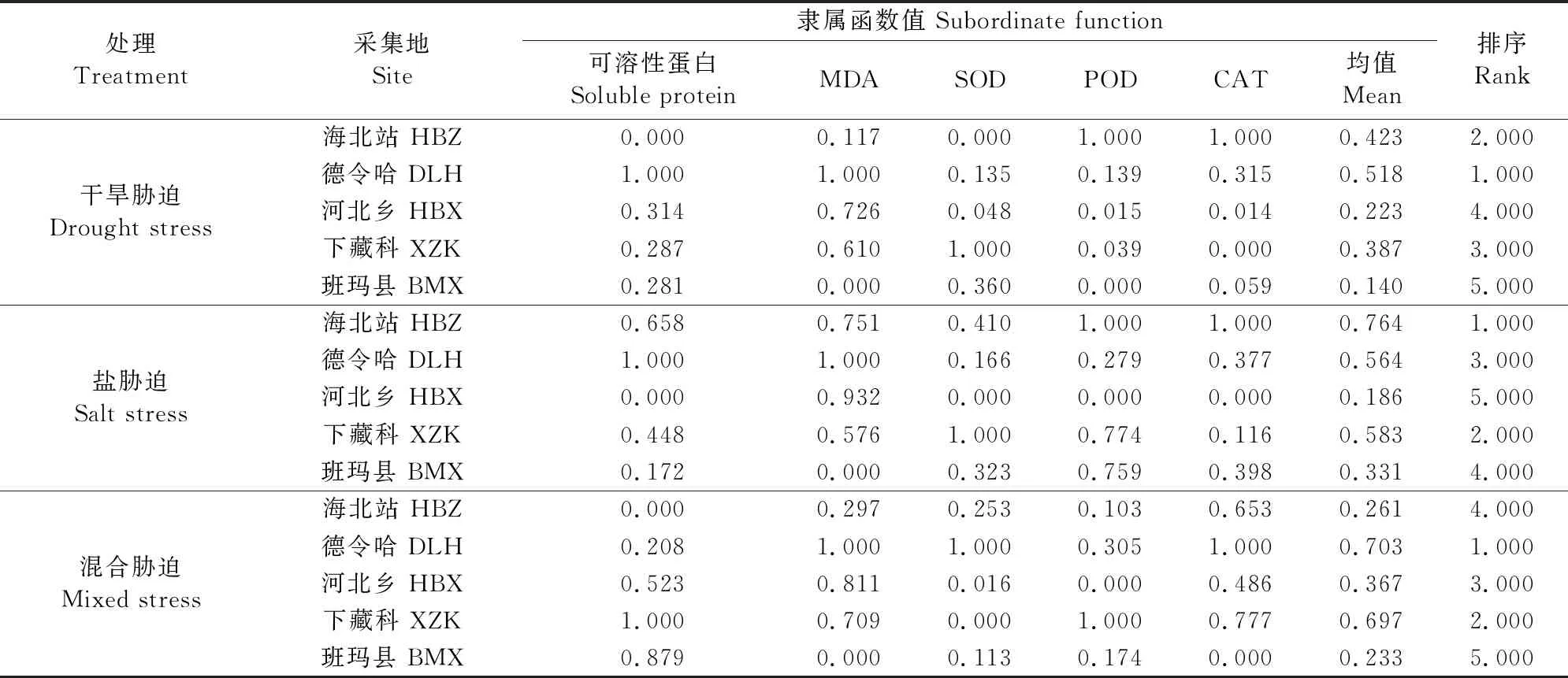
表3 不同地理种源麻花艽的抗性隶属函数值及综合评价
3 讨 论
3.1 不同胁迫对麻花艽生长的影响
胁迫能抑制植物的生长,本研究发现不同胁迫下大多数材料的生长受到明显抑制,鲜重无显著性变化,且混合胁迫下对株高的抑制更为明显,这与李静怡等[14]对天仙子NaCl和PEG胁迫的结果相一致。虽然胁迫下植物的生长受到抑制,但相关研究也发现,植物在水分胁迫下倾向于将更多的资源分配给根系,通过提高根冠比来适应干旱的环境[15]。本研究中植物胁迫下的根冠比并未出现明显增加,但海北站幼苗的根冠比在胁迫下显著高于其他地区,说明其对胁迫环境具有一定的适应性。
3.2 不同胁迫对麻花艽生理特性的影响
MDA能够造成细胞膜的损伤,其含量高低可以作为植物抗性强弱的判定指标之一,多数植物在胁迫下MDA含量都会出现明显升高[16]。本研究发现班玛县麻花艽幼苗在3种胁迫条件下MDA含量都出现明显积累,远大于其他地区,表明胁迫对其影响较大。而德令哈的MDA变化却不明显,干旱胁迫下更是出现明显下降,表明其对胁迫的耐受性较强。胁迫下植物可以通过增加可溶性蛋白等渗透调节物质的含量降低自身对水分的需求。本研究发现干旱胁迫下海北站和德令哈地区幼苗可溶性蛋白含量出现明显增加,盐胁迫下仅德令哈地区出现增加,说明该地区植物对胁迫表现出一定的生理适应。李为民等[7]发现混合胁迫能够导致野生大豆渗透调节物质可溶性糖含量的降低,本研究发现混合胁迫下所有地区可溶性蛋白含量都出现降低,推测可能是由于强胁迫下,高浓度的Na+导致细胞内外的离子平衡受到破坏,影响了可溶性蛋白的积累。
植物在胁迫条件下会出现活性氧的积累,其细胞膜结构会遭到破坏,导致生长受到抑制,植物会通过在体内建立一套相应的抗氧化保护酶系统(SOD,POD和CAT等)以维持其体内的活性氧动态平衡,实现对胁迫的耐受[7]。本研究中,大部分材料在盐和混合胁迫下抗氧化酶活性都出现降低;干旱胁迫下POD活性出现增加,说明麻花艽对干旱具有一定的耐受性。陈锋[17]对祁连山3种药用植物抗旱性的研究也发现,干旱胁迫下麻花艽的SOD和POD出现升高,但随着胁迫时间的增长,其抗氧化酶活性迅速降低,由此推测本研究中麻花艽幼苗抗氧化酶活性的降低,可能是由于强胁迫超过了保护酶系统的承受极限,导致抗氧化酶系统受到损伤[18]。虽然大部分麻花艽材料抗氧化酶活性出现了降低,但部分地区还是明显升高,例如单纯干旱和盐胁迫处理下,海北站地区幼苗的POD和CAT活性出现明显增加,混合胁迫下德令哈地区的SOD和CAT增加也最为明显。说明这两个地区的麻花艽在胁迫下抗氧化酶活力得到增强,其对胁迫的耐受性也强于其他地区的麻花艽。
3.3 不同种源麻花艽的抗逆性评价
模糊隶属函数法现已广泛应用于品种的抗逆性评价上,能够综合所有指标对植物进行客观全面的评价,避免了单一指标评价的片面性[12-13,19]。本研究中,通过对麻花艽种质的抗逆性评价,发现德令哈和海北站的麻花艽抗旱性较强,海北站、下藏科和德令哈的抗耐盐性较强,这也和形态指标表现相一致,胁迫下海北站和德令哈的麻花艽具有较高的株高、根长和根冠比。有趣的是,虽然海北站麻花艽具有较强的抗旱耐盐性,但在混合胁迫下,其抗性却远低于德令哈,说明麻花艽对逆境的响应是一个复杂的过程,并不是单一累加的结果[14]。不同种源麻花艽抗性的差异也与种质所处的环境气候条件相吻合,德令哈属于柴达木盆地荒漠区,其年降水量较少,而班玛县则属于川西藏东高山峡谷针叶林区,受印度洋季风的影响,其年降水量较多[20],说明各地理种源麻花艽经过长期的自然选择和环境适应对胁迫响应产生了一定的差异,原产地生长于寒冷干旱地区的植物, 对水分缺乏不敏感,例如德令哈、海北站地区。而温暖湿润地区, 例如班玛县的麻花艽,其对水分缺乏则较为敏感[21]。
4 结 论
综上所述,胁迫对麻花艽的生长具有一定的影响,但其能够通过植物形态、生理生化等机制的调节实现对胁迫的耐受,且不同地理种源麻花艽对胁迫耐受性存在一定的差异,其中海北站和德令哈地区的麻花艽由于长期受自然环境的影响具有较强的抗性,主要通过增加渗透调节物质含量、提高抗氧化酶活性等机制,实现了对胁迫的耐受。后续应加强对具体调节机制的研究,扩大材料的选取范围,为麻花艽优质种质资源的引进栽培提供理论依据。
猜你喜欢
飞天(2022年5期)2022-05-18
意林彩版(2022年1期)2022-05-03
绿色科技(2021年21期)2021-11-26
阅读与作文(小学高年级版)(2021年8期)2021-09-12
种子(2021年3期)2021-04-12
星星·散文诗(2019年5期)2019-07-19
读友·少年文学(清雅版)(2018年1期)2018-09-11
阅读与作文(小学高年级版)(2017年7期)2017-08-04
福建林业(2016年4期)2016-08-16
诗歌月刊(2014年12期)2015-04-14
