
基于不同位点的广东省柑橘黄龙病菌种群分子多样性分析
2020-01-17 06:33黄洪霞邓晓玲许美容
华南农业大学学报 2020年2期
黄洪霞,郑 正,邓晓玲,许美容
(华南农业大学 农学院/广东省微生物信号与作物病害防控重点实验室,广东 广州 510642)
柑橘黄龙病是柑橘生产上的重要病害,严重制约着我国柑橘产业的发展。根据文献记载,19世纪末在我国广东潮汕地区就已经有黄龙病的发生报道[1]。至今黄龙病已在亚洲、非洲、南美洲、北美洲和大洋洲的近50个国家和地区传播和蔓延[2]。目前已知黄龙病菌属于α-变形菌纲Alphaproteobacterial候选韧皮部杆菌属CandidatusLiberibacter的革兰阴性细菌。与黄龙病相关的CandidatusLiberibacter有 3 个种:亚洲种 (“Ca. L. asiaticus”, CLas)、非洲种 (“Ca. L. africanus”, CLaf)和美洲种 (“Ca. L.americus”, CLam)[3],我国仅有 CLas分布[4-5]。柑橘黄龙病在广东地区已有近百年流行历史,分析广东省内不同地理区域的CLas分子遗传多样性对其余国家及地区的CLas多样性研究具有一定的参考意义。
Jagoueix等[3]从分子水平上确定了黄龙病菌属于细菌,之后对黄龙病菌遗传多样性的研究主要集中在保守基因的单核苷酸多态性(Single nucleotide polymorphisms, SNPs)分析,包括 16S rDNA 序列、16S/23S r RNA 基因间隔区[6-10]、β 操纵子[9]以及外膜蛋白 (Outer membrane protein, OMP)基因,但基于这些相对保守位点的CLas遗传多样性程度很低。自2009年Duan等[11]公布了CLas全基因组序列及2011年Zhang等[12]公布了其中的2个噬菌体SC1和SC2之后,许多高变异的位点被发掘以供黄龙病菌种群多样性研究,这些变异位点包括原噬菌体区域位点、短串联重复序列和转座子等。在CLas基因组中噬菌体多样性研究方面,Liu等[13]通过研究CLas上1个原噬菌体位点(CLIBASIA_05610),发现该基因在广东和云南黄龙病菌群体中的出现频率存在显著差异,在云南群体出现的频率比广东群体更高。Zhou等[14]基于Psy62菌株基因组的原噬菌体区域的串联重复序列鉴定出黄龙病菌亚洲种CLas 2个相关联的高变异基因(hyvⅠ和hyvⅡ),揭示了黄龙病菌在不同国家和地区的种群多样性,首次报道不同CLas种群共存于同一样品中。Zheng等[15]基于2种原噬菌体对我国柑橘黄龙病菌群体进行原噬菌体鉴定,发现我国的黄龙病群体主要携带单一类型的原噬菌体,广东省及周边省份以携带Type II类型原噬菌体的菌株为主,而云南则以含有Type I类型的菌株为主。2018年,Zheng等[16]报道了第3种类型的原噬菌体。在此基础上,李嘉慧等[17]基于原噬菌体类型对我国CLas遗传多样性进行分析,指出我国南方地区的CLas种群分为3个组,且只携带Type II类型原噬菌体的菌株为华南地区的优势种群。在基因组的短串联重复序列多态性方面,Chen等[18]在CLas的CLIBASIA_01645位点上发现广东和美国佛罗里达种群在串联重复序列数目(Tandem repeat number,TRN)存在变化并具有显著差异:佛罗里达的TRN=5(84.4%)的种群占主导地位,而广东种群的TRN值更加多样化,表现为TRN=7(47.6%)。Ma等[19]根据CLIBASIA_01645位点分析我国南方8个省份的CLas种群遗传结构,发现来自广东的CLas种群与其他省份存在明显差异。Katoh等[20]通过分析柑橘黄龙病菌CLas Psy62菌株的全基因组,选择了4个高度多态性的简单重复基因位点(Simple single repeat,SSR),结果表明,使用多个可变数目串联重复序列(Variable number tandem repeat, VNTR)基因位点比仅使用单个VNTR基因位点的分析能更准确地揭示遗传多样性。Islam等[21]开发了7个微卫星DNA(串联重复序列)位点对亚洲、美洲的几个国家(地区)的CLas进行遗传多样性分析,结果发现,亚洲的CLas遗传多样性比美洲丰富。许美容等[22]基于CLas基因组中全套短串联重复序列(Short tandem repeat, STR),通过 PCR 和聚丙烯酰胺凝胶电泳 (Polyacrylamide gel electrophoresis, PAGE)试验分析我国南方6个省份的CLas菌株,结果表明各省的菌株均表现出较高的遗传多样性,且广东省的黄龙病菌株具有一定的遗传特异性。在基于微型反向重复转座元件 (Miniature inverted-repeat transposable element, MITE)的多样性方面,Wang 等[23]在 CLas原噬菌体区域发现了2个具有类似MITE序列特征的微型转座子MCLas-A和MCLas-B,并在佛罗里达州的CLas种群中检测出MCLas-A(B720)和MCLas-B(B630)共存的状态,而在我国的CLas种群中,只含有其中一种转座子。
综上所述,相对于 16S rRNA 基因、16S/23S r RNA基因间隔区、β操纵子以及omp等保守基因,基于噬菌体区域、STR(或SSR、VNTR)及MITE等高变异位点的CLas种群遗传多样性更为丰富,且广东省的CLas种群多特异于其他省份。但至今鲜见基于高变异位点单独对广东省的Clas种群进行分析的报道。本试验在上述研究基础上,利用3种类型的高变异位点:原噬菌体区域、MITE[23]和STR位点[22],对广东省的176个不同地理来源的CLas菌株进行遗传多样性分析。较系统地分析黄龙病发生历史最为悠久的广东省各地市的CLas基因水平的多样性,以明确黄龙病菌在历史病区的遗传分化情况,以期为CLas种群进化提供理论支持。同时,进一步分析与探讨基于柑橘黄龙病菌多种基因位点遗传多样性分析方法的科学性。
1 材料与方法
1.1 病原材料
黄龙病菌株样品来源于广东省清远、韶关、肇庆、梅州、惠州、深圳、江门、茂名、湛江和广州市,采集自带有典型斑驳或者黄化症状的感染黄龙病的柑橘叶片样品,总计176个。每份样品为取自同一株病树同一枝条上的3片具有典型黄龙病症状的叶片。
1.2 DNA 提取
剪取柑橘叶中脉100 mg,参照OMEGA植物DNA提取试剂盒(OMEGA bio-tek)说明书提取总DNA。取1~2 μL提取的DNA溶液,用Thermo NanoDrop One 超微量紫外分光光度计进行DNA质量的检测,合格的DNA样品稀释至约100 ng/μL,保存于−20 ℃冰箱用于下一步试验。
1.3 PCR 及检测
所用引物的特异性已进行多重验证,引物的信息见表1。用Real-time PCR验证所采样品中是否含有CLas及CLas阳性的样品中的噬菌体类型;用常规PCR扩增转座子位点CLIBASIA_05620的基因片段及2个具有STR的基因位点(CLIBASIA_03080和CLIBASIA_01215)。

表1 特异性引物信息Table 1 Information of specific primers
1.3.1 Real-time PCR CLas的检测采用基于 Li等[24]设计的特异性引物和探针HLBas/HLBp/HLBr的Real-time PCR 法,对 Real-time PCR 检测得到的Ct值<30的阳性样品进行后续试验。Real-time PCR 扩增使用 Bio-Rad CFX Connect系统。反应体系共 20 μL,包括 10 μL Bestar qPCR Master Mix(DBI Bioscience),1 μL DNA 模板 (100 ng),0.2 μL PCR 探针 (10 μmol/L),上下游引物 (10 μmol/L)各0.4 μL,8 μL ddH2O。扩增程序为:95 ℃ 2 min;95 ℃10 s,58 ℃ 30 s,40 个循环,在每个 58 ℃ 步骤结束时捕获荧光信号。使用Bio-Rad CFX Manager 2.1软件进行数据分析,基线和阈值设置为软件默认。
1.3.2 常规 PCR 转座子位点的 PCR 反应体系为 25 μL,包括 17.6 μL 的 ddH2O,2.5 μL 的10×DNA 聚合酶缓冲液,2.5 μL 的 2.5 mmol/L 的dNTPs,0.4 μL 的 2.5 U/μL Taq DNA 聚合酶(Tiangen Biotech Co.),上下游引物 (10 μmol/L)各 0.5 μL和 1 μL DNA 模板 (50~100 ng)。扩增程序为:在 94 ℃下初始变性 2 min,扩增 35 个循环 (94 ℃ 30 s,55 ℃30 s和 72 ℃ 30 s),最后 72 ℃ 延伸 7 min。将 PCR产物在10 g/L琼脂糖凝胶中电泳(0.5×TBE缓冲液),然后在凝胶成像系统下观察结果,拍照并保存。STR的常规PCR反应体系为15 μL,反应液体系和扩增程序参照以上扩增MITEs的体系设置,PCR退火温度为57 ℃。PCR产物经10%(w)PAGE电泳(1×TBE缓冲液)后,用硝酸银染色,在白瓷板上拍照并保存。
1.3.3 电泳带型读取及校准 根据琼脂糖凝胶电泳结果,对CLIBASIA_05620~CLIBASIA_05625扩增的部分PCR产物进行胶回收然后克隆测序,根据测序结果分成以下2种带型:B720和B350。参考Wang等[23]的分型方法,将719和731 bp的条带归为B720(约720 bp,含有MCLas-B类型转座子的片段);将 303、291 和 292 bp 归为 B350(约 350 bp,缺失MCLas-A类型转座子的片段)。PAGE电泳的DNA条带读取根据条带相对分子质量由小到大依次读取,并编号。根据电泳图片以及读取的结果,将部分相同相对分子质量的样品重新在同一块胶中进行PAGE电泳,以对数据进一步校准。
1.4 聚类分析
根据Real-time PCR和常规PCR结果计算含有不同类型原噬菌体的CLas菌株在不同地区样本中所占的比例以及转座子位点、短串联重复序列所扩增出来的条带类型。使用Popgene v1.32(https://www.ualberta.ca/~fyeh/popgene.html)软件,基于Nei’ s(1972)genetic distance的分析方法,对广东省10个不同地区的CLas种群进行聚类分析,用MEGA 6.0制图,方法参照文献[25]。
2 结果与分析
2.1 基于噬菌体类型的广东地区柑橘黄龙病菌的种群多样性
通过对176个黄龙病样品进行噬菌体特异性引物Real-time PCR,共鉴定出6种类型的CLas菌株(表2)。85.8%的黄龙病菌株样品中仅能扩增出1种噬菌体,其余样品中含有2种或2种以上的噬菌体。仅含有其中1种噬菌体的151个样品中,有150个菌株(99.3%)只携带Type II类型噬菌体,该类型CLas菌株为广东省优势菌株。同时携带2种不同类型噬菌体的菌株为23个(13.1%),主要为Type I + Type III类型 (15 个,8.5%),其次是 Type I +Type II;同时携带 3 种类型噬菌体 (Type I + Type II +Type III)的菌株数较少。其中携带 Type I + Type III噬菌体的样品主要来自茂名,Type I + Type II类型则主要来自湛江。本研究未检测到只携带Type I类型噬菌体的菌株和均不含有上述3种类型噬菌体的菌株。Type I和Type III噬菌体更倾向于与别的噬菌体复合存在(Type I噬菌体单独存在与复合存在的比率是0∶24;Type III噬菌体单独存在与复合存在的比率是1∶18),而Type II噬菌体则倾向于单独存在(单独存在与复合存在的比率是150∶10)。另外,结合采集的柑橘品种进行研究,结果发现菌株所携带噬菌体的类型与柑橘品种没有相关性。
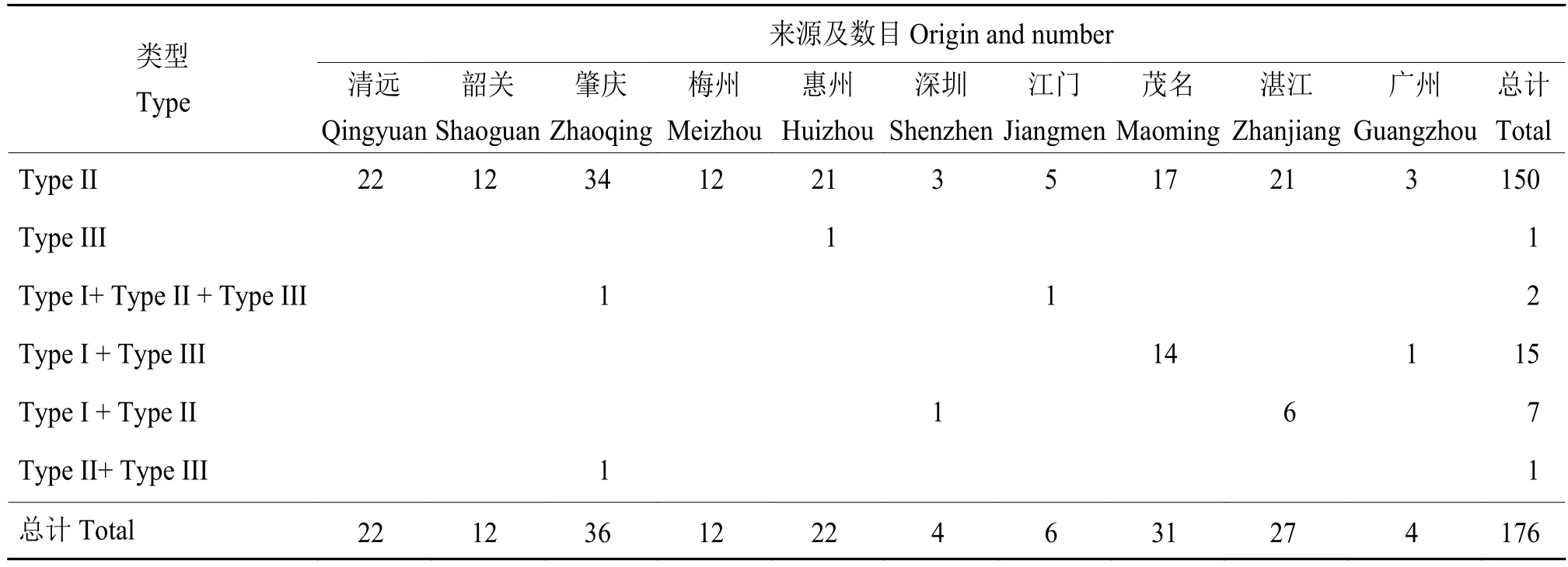
表2 广东省柑橘黄龙病菌株携带噬菌体类型的情况Table 2 Phage types carried by the “Candidatus Liberibacter asiaticus” strains in Guangdong Province
2.2 基于转座子位点的广东地区柑橘黄龙病菌的种群多样性
使用LapPF1-f/LapPF1-r引物对广东省176个CLas菌株的常规PCR所产生的条带类型有4种:B720、B350、B720+B350 和没有条带 (图1)。具有这4种条带的CLas菌株在不同地区样品中的分布见表3。转座子CLIBASIA_05620~CLIBASIA_05625位点扩增结果显示,广东省B350类型条带最多(占76.7%);清远和韶关的CLas样品中只扩增出B350类型的条带,说明其种群结构可能比较单一;而茂名样品扩增出B720+B350的比例较高,占到54.8%,说明其种群变异相对较大;湛江样品扩增出B350和B720的比例相当,均为40.7%(表3)。
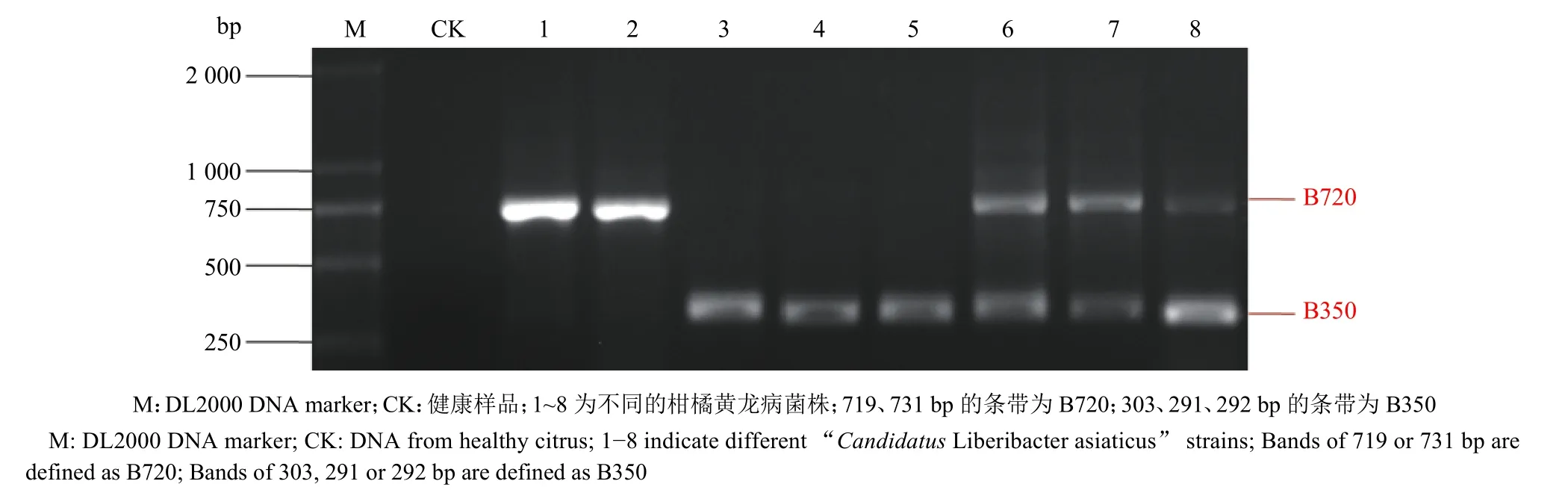
图1 LapPF1-f/LapPF1-r引物对广东部分柑橘黄龙病菌株的PCR扩增条带Fig. 1 PCR amplification bands of partial “Candidatus Liberibacter asiaticus” strains in Guangdong Province with primers of LapPF1-f/LapPF1-r

表3 基于转座子CLIBASIA_05620~CLIBASIA_05625位点对广东省柑橘黄龙病菌株的扩增条带分析Table 3 Analysis of amplified bands of “Candidatus Liberibacter asiaticus” strains in Guangdong Province based on transposon sites in CLIBASIA_05620-CLIBASIA_05625 gene
2.3 基于短串联重复序列的广东地区柑橘黄龙病菌的种群多样性
根据许美容等[22]开发的33个获得有效扩增的STR位点,从中选取2个多态性的位点STR1和STR12。如图2所示,基于STR序列2个不同位点对广东省柑橘黄龙病菌株扩增出来的条带类型均较为多样;如表4所示,STR1位点扩增出9种条带,分别为含有大小不一的单一条带、2种条带组合或没有条带的情况。STR1位点扩增产物为单一条带、不含条带和复合条带的比例分别为133∶4∶39。扩增产物为150~160 bp条带的菌株在广东省内大部分地区的比例都比较高,占56.8%,为优势菌株。其中湛江的菌株多扩增到150~180 bp条带,而茂名菌株则多同时扩增出140~150和150~160 bp条带。
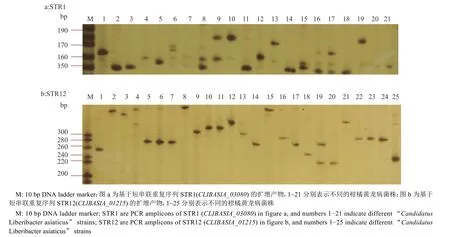
图2 STR1/STR12引物对广东部分柑橘黄龙病菌株的PCR扩增条带PAGE电泳结果Fig. 2 PAGE electrophoresis of PCR amplified bands of partial “Candidatus Liberibacter asiaticus” strains in Guangdong Province with primers of STR1/STR12

表4 基于短串联重复序列CLIBASIA_03080位点对广东省柑橘黄龙病菌株的扩增条带分析Table 4 Analysis of amplified bands of “Candidatus Liberibacter asiaticus” strains in Guangdong Province based on short tandem repeats in CLIBASIA_03080 gene
如表5所示,STR12位点扩增出10种条带。STR12位点比STR1位点多样性更丰富。STR12引物扩增产物单一条带、不含条带和复合条带的比例分别为 141∶19∶16。240~360 bp 的单一扩增条带占多数,而复合有2种类型条带的菌株为少数。其中湛江的样品多为240~280 bp的条带,茂名的菌株则多扩增出280~300 bp的条带。韶关地区的较为单一,使用STR1F/STR1R引物只扩增出150~160 bp这一种条带,使用STR12F/STR12R引物扩增出的条带集中在280~360 bp。另外,结合噬菌体类型和STR 2种位点的结果显示,含有2个及2个以上噬菌体的25个菌株中STR1扩增具有2个条带的样品数目为15个;而单个噬菌体的151个菌株中STR1扩增出多条带的样品数为24个。
2.4 基于 3类多态位点的黄龙病菌株聚类
分别基于原噬菌体区域、转座子位点和短串联重复序列3种多态性位点对176个CLas菌株作出4个聚类图(图3)可见,基于不同位点的CLas种群可分为2大组,但不同位点的具体情况又有所不同。从4个聚类图上看,湛江、茂名和深圳的菌株与其余地区的菌株遗传距离较远;对MITEs和STR1位点,湛江的菌株异于其余地区的菌株。
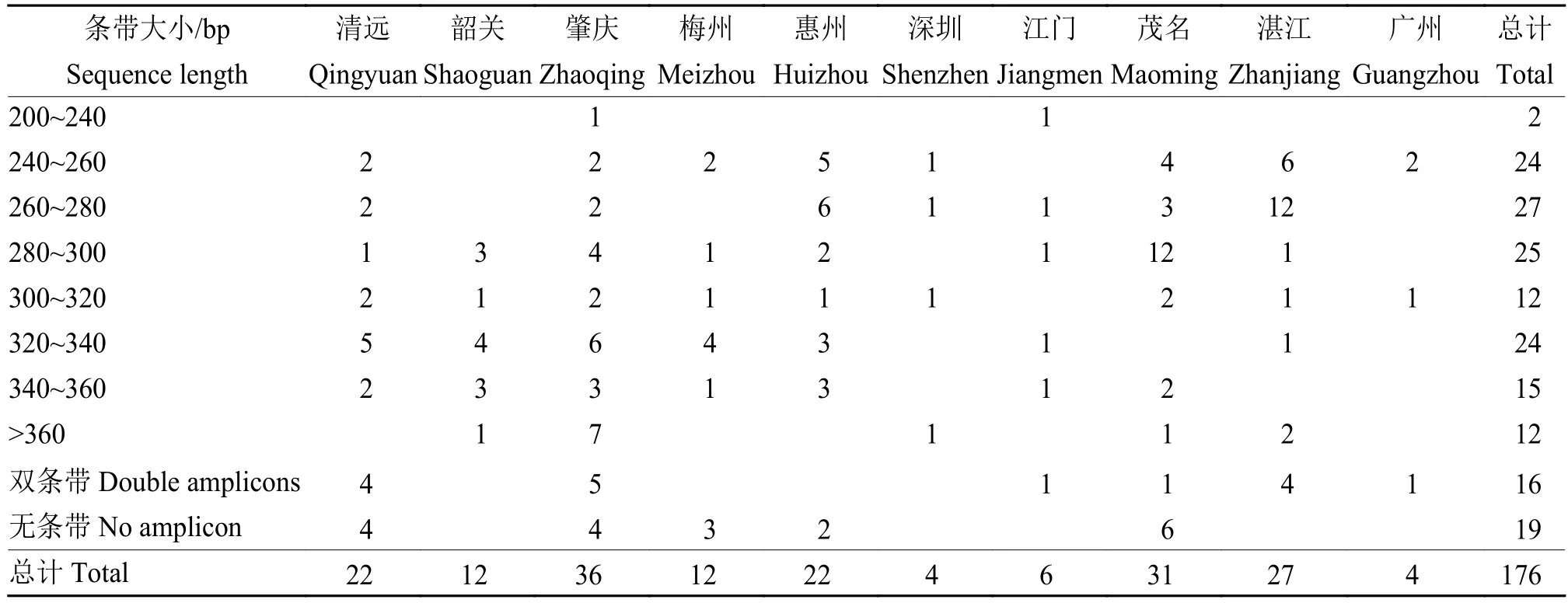
表5 基于短串联重复序列CLIBASIA_01215位点对广东省柑橘黄龙病菌株的扩增条带分析Table 5 Analysis of amplified bands of “Candidatus Liberibacter asiaticus” strains in Guangdong Province based on short tandem repeats in CLIBASIA_01215 gene

图3 基于不同位点的广东地区黄龙病菌种群的聚类分析Fig. 3 The dendrogram analysis of “Candidatus Liberibacter asiaticus” populations in Guangdong Province based on different gene loci
综合所有4个位点的数据对这176个样品作聚类分析(图4)。总的聚类图与每个位点做的聚类图均不完全一致,可见基因水平的遗传多样性是复杂的。只有清远和梅州的样品在4个聚类图以及3种位点总的聚类图中,都聚为一组,说明这2个地方的菌株的遗传距离较近。
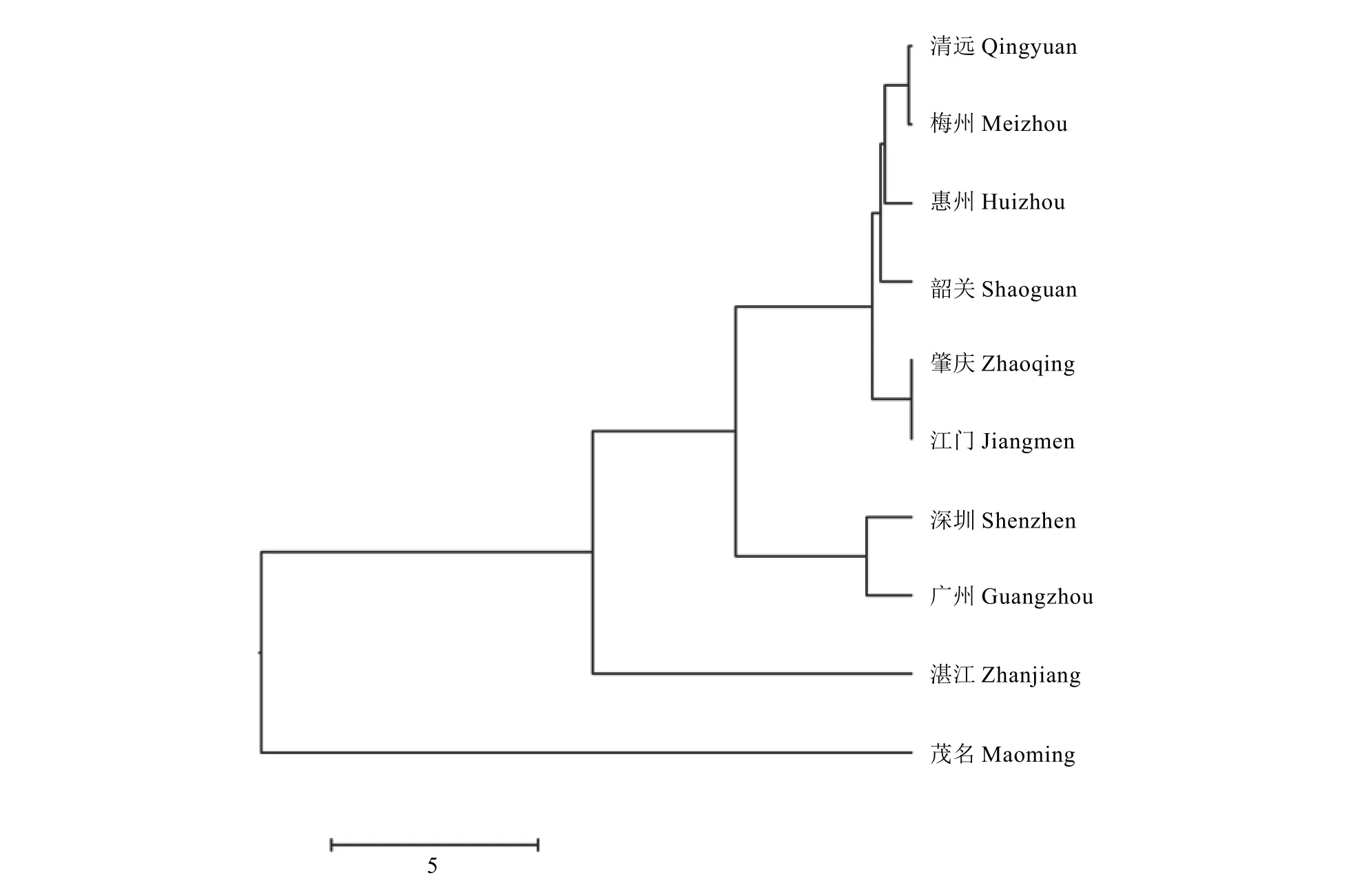
图4 基于6个位点的广东省柑橘黄龙病种群的聚类Fig. 4 The dendrogram of “Candidatus Liberibacter asiaticus” populations in Guangdong Province based on six gene loci
根据这3种不同位点在广东省不同地区黄龙病菌基因组中的Nei’ s (1973) 基因多态性 (H值),对多态性进行排序,基于STR12位点的多态性(0.880 5)>基于 STR1 位点的多态性 (0.633 3)>基于转座子位点的多态性(0.388 8)>基于原噬菌体区域位点的多态性(0.197 8)。从聚类图也可看出基于STR12位点的CLas种群遗传结构较复杂,说明该位点为高度变异位点。
3 讨论与结论
黄永辉等[26]基于omp基因单个位点分析了广东省柑橘产区7个城市黄龙病菌的遗传多样性,发现广东地区的CLas具有很高的同源性,同时也存在一定的遗传变异。Kenta等[10]通过16S rDNA、16S/23S rRNA基因间隔区、omp基因区、trmU-tufB-secE-nusG-rplKAJL-rpoB区域(基因簇区域)和噬菌体型 DNA聚合酶 (Bacteriophage-type DNA polymerase)区域的DNA片段序列研究东南亚不同寄主和不同地理区域的CLas分离株的遗传多样性和亲缘关系,发现噬菌体型DNA聚合酶区域序列多样性最为丰富,该位点可用于东南亚地区CLas分离株的分子鉴定。全基因组序列分析也证明CLas的噬菌体区域的碱基变异最大。本研究的176个菌株基于噬菌体类型、MITE及2个STR位点的多样性都较为丰富,一方面说明保守的基因位点不适用于分析CLas种内的多样性,而种间差异分析时则可参考保守的基因序列;另一方面也表明要全面地探清某一物种的基因多样性,应尽可能地用多基因位点综合分析,甚至用全基因组序列。类似地,Katoh等[20]认为单一的串联重复位点无法真实地反映CLas的遗传多样性。本研究结果证明分别基于2个STR位点的序列多态性聚类结果差异较大;基于不同类型位点(噬菌体类型、MITE和STR)的多样性分析结果也各有异同。广东省不同地区的CLas菌株在不同的基因位点的变异程度有一定差异,这可能是不同类型的位点在进化过程中选择压力不同造成的。根据不同多态性位点的基因型对CLas菌株的遗传分析具有一定的差异,进一步说明基于单个位点对CLas的遗传结构进行分析具有较大的局限性。
目前在黄龙病菌基因组中检测到3种原噬菌体类型,即 Type I(SC1, NC_019549.1)、Type II(SC2,N C_0 1 9 5 5 0.1)和 T y p e I I I(P-J X G C-3,KY661963.1)[16,27]。李嘉慧等[17]根据这 3 种噬菌体分析了中国8个省份548个CLas菌株的多样性,该研究与本研究的噬菌体类型部分结果是一致的:仅携带Type II噬菌体的菌株占多数(分别占76.7%和85.2%),其次为同时携带Type I和Type III(分别占8.59%和8.52%)及同时携带Type I和Type II(6.13%和3.98%)噬菌体组合的菌株,仅个别菌株携带其余类型的噬菌体或组合。2个研究均发现 Type I噬菌体偏向于和 Type II和/或 Type III类型的噬菌体组合存在于广东省的CLas菌株中。da Silva等[28]分析了巴西来源于 São Paulo 州的573个CLas菌株的3个STR位点和噬菌体类型,发现这些CLas菌株的STR复杂,主要含Type I噬菌体,指出基因重组现象可能是导致巴西CLas群体同源性差异的原因。此外,150个含Type II噬菌体的菌株中有88.67%在目的转座子位点为缺失 MClas-A型,STR1位点扩增出150~160 bp 条带的概率为 64%;26 个非 Type II噬菌体菌株中,以上比例相应为7.69%和15%,3种位点的优势菌株的相关性分析表明含Type II类型噬菌体的菌株更可能携带无MClas-A型MITE及150~160 bp条带STR1位点。本研究的广东省内各地区的菌株携带噬菌体类型的多样性低于之前研究中我国不同省份间的菌株多样性,这也证明了与苗木调运相关的柑橘区域化种植对该病害的蔓延传播具有关键影响。结合噬菌体位点和2个STR位点的结果分析发现,含有2个及以上噬菌体菌株中STR1扩增条带数为2个以上的比例高达60%,推测含有2种以上噬菌体的样品可能混有2个以上CLas菌株。在MITE位点上,与Wang等[23]之前报道的MITE类型多样性相比,本研究的菌株中未扩增出B630的条带(含有MCLas-A类型转座子的片段),该类型样品缺失的原因有待进一步研究。
本研究在各地采集的样品数目不同;不同地区的样本有的来源单一(同一果园),有的则来源较为复杂(多个果园);样本也并非来自相同背景(柑橘的品种、树龄、地理条件、栽培管理水平等)的果园;未收集材料的原始来源(引种)信息;聚类分析中每个类型的多态位点未附以不同的分值;以上几点差异可解释部分试验误差。分析地理特异性柑橘品种上的CLas菌株多样性更具有代表性和针对性;对同一地区不同年代的样品进行分析也是研究菌株分子变异的一个思路。
猜你喜欢
新高考·高三数学(2022年3期)2022-04-28
昆明医科大学学报(2022年2期)2022-03-29
林业科学(2022年1期)2022-03-23
植物保护(2021年4期)2021-11-12
浙江农林大学学报(2021年3期)2021-07-12
内蒙古民族大学学报(社会科学版)(2020年1期)2020-11-03
科学24小时(2020年4期)2020-05-14
中国农业科学(2020年7期)2020-04-11
文苑(2018年22期)2018-11-15
陕西画报(2016年1期)2016-12-01
