
基于树轮气候相关分析法的杉木径向生长与气候因子关系
2020-01-10 06:18:12胡亚楠黄智军吴鹏飞马祥庆
福建农林大学学报(自然科学版) 2020年1期
张 辉, 颜 耀, 胡亚楠, 黄智军, 吴鹏飞, 马祥庆
(1.福建农林大学林学院;2.国家林业局杉木工程技术研究中心,福建 福州 350002)
杉木[Cunninghamialanceolata]是我国人工林面积最大的用材林树种,分布于我国秦岭、淮河以南17个省区.根据森林资源清查数据[1],杉木人工林面积为854万hm2,蓄积量占全国人工林的29.2%,在我国林业生产中占有重要地位.近年来随着气候变化的加剧,对杉木人工林产量和质量造成的影响日趋严重,如何揭示杉木生长对气候变化的响应和提升杉木的抗逆性成为林业生产中急需解决的重大课题[2].
树木年轮生态学是研究树木径向生长对气候变化适应性的主要方法[3],广泛应用于全球气候变化研究领域.通过研究树木径向生长对气候因子的不同响应,提高对树木生态学的认识,且通过树木年轮剖析获取的信息具有连续性强、分辨率高、环境变化指示意义明确等优点[3].因此通过树木年轮长时间生长序列与每月气候因子之间的相关分析,可以找出限制树木径向生长的关键气候因子和每年(月或季节)树木受影响最大的时间[4].
近年来,我国相关杉木年轮生态学研究逐渐成为热点[5].朱安明等[6]研究表明年平均温度升高,抑制杉木生长.李建等[7]研究表明湿润度指数是限制杉木径向生长的主要因素.刘兰妹等[8]研究表明杉木径向生长对温度和降水的响应存在差异.但早材宽度、晚材宽度和整轮宽度对月份水平气候因子响应方面的研究较缺乏[3].由于早材形成于春季,高效的水分运输,使细胞分裂强烈,扩大时间长,产生大量宽而薄的早材细胞;晚材形成于夏季和秋季,有限的淀粉和水分含量,使细胞分裂减慢,而扩大时间缩短,产生数量较少、窄而厚的晚材细胞[9-10].因此早、晚材宽度包含整轮宽度所无法获取的环境气候信息[9,11],对进一步了解树木径向生长对其环境变化的响应机制具有重要意义.
鉴于此,本研究以树轮气候相关分析方法,利用亚热带杉木中心产区60年生杉木年轮数据,通过建立杉木树轮自回归年表(ARS),分析杉木径向生长(整轮宽度、早材宽度、晚材宽度)与主要气候因子(温度、降水)的相关关系,揭示不同气候因子对杉木径向生长影响的差异,为杉木人工林高效经营和杉木年轮气候学研究提供依据.
1 试验地概况
试验地位于福建省三明市三元区福建农林大学莘口林场(26°11′—26°13′N ,117°30′—117°33′E),属于亚热带季风气候,年无霜期240~300 d,年降水量1 600~1 800 mm.试验地海拔250~300 m,1958年春造林,初植密度2 000株·hm-2[12].2017年林分平均胸径19.68 cm,平均树高18.08 m,林分现存密度1 109株·hm-2.土壤为山地红黄壤,容重1.09 g·cm-3,总孔隙度54.42%,最大持水率44.71%,pH 4.08,林下主要植被有粗齿桫椤(Alsophiladenticulate)、芭蕉(Musabasjoo)、芒萁(Dicranopterisdichotoma)等.
2 研究方法
2.1 样品采集及预处理
依据国际树木年代学研究的标准方法[13]采集样品.优势木可以指示林分的立地条件,标准木可以指示整个林分的生长状况,因此本研究首先通过对9个20 m×20 m样地进行调查,选取18棵杉木优势木、9棵杉木标准木.优树木为每一样地最高的2株,平均标准木胸径与林分平均胸径相差不超过半个径阶,其树高与林分平均高相差不超过5%.
于2017年12月分别伐倒选择的优势木和标准木,以2 m等截距取下圆盘,进行树干解析.带回实验室并用扎带绑紧防止裂缝,晾干处理.选用1.3 m处无病虫害轮盘,按照标准逐一使用300、600、800目砂纸打磨,测量整轮宽度并定年;之后每个轮盘在无裂缝处均匀切割3~6根树芯,树芯宽约2 cm,共采集到27棵树104根树芯,再使用量角显微镜测量纤维丝角度,用双片轮锯切割样品薄片,薄片平均厚度为1.019 mm,酒精蒸馏48 h,以去除油脂等杂质,X光照射、拍片.
2.2 宽度测量及年表建立
首先目视交叉定年,消除伪年轮(细胞前后渐变过程为伪轮)和缺轮,保证同一轮盘不同方向同一年轮相互照应;采用精度为0.001 mm的第六代年轮分析仪测量树木年轮宽度,并利用COFECHA程序对交叉定年结果进行质量控制和检验[14],以保证年轮的准确性.然后采用Dendro2003 X射线年轮系统分析仪,准确测量整轮宽度、早材宽度和晚材宽度[15-16],避免传统人为区分早、晚材的误差.再次使用COFECHA程序,以确定是否将序列输入年表,剔除异常与主序列关系不显著的样本序列,共挑选出23棵树43根芯建立整轮宽度年表,选出21棵树42根芯建立早材宽度年表,选出19棵树36根芯建立晚材宽度年表.数据分析主序列年轮长度为56 a.通过ARSTAN程序的负指数函数、样条函数去除年龄因素、立地条件、林分差异等非气候因素对杉木径向生长的影响[17-18],最终建立标准年表(STD)、残差年表(RES)和自回归年表(ARS).由于ARS年表不仅能够消除个体之间生长差异影响,而且保留群体共有的高频和低频变化信息[17-20],因此本研究选用ARS年表.
2.3 气象数据统计分析
气象资料来自离样地最近的永安气象观测站(25°58′N,117°21′E),该区域属于亚热带季风气候.从国家气象科学数据共享服务平台(http://data.cma.cn)下载气象因子数据,从气象站记录的多年气候平均状况(图1)来看,该区域存在明显的干湿季节变化,秋冬季低温少雨,5—6月降雨达到峰值,7—9月受海洋偏南气流影响进入高温少雨的伏旱期,多年平均气温19.5 ℃,1月平均气温9.5 ℃,7月平均气温28.4 ℃,极端最高气温40.5 ℃,极端最低气温-7.6 ℃,多年平均降水量1 560.2 mm,年平均相对湿度78.6%.本研究选取月平均温度、月平均最高温、月平均最低温、月平均降水量为主要气候因子,研究时间为1961—2016年.通过对永安气象站1961—2016年的年均温度和年均降水量进行变化趋势分析(图1)发现,研究区年平均气温呈显著上升趋势(P<0.001),以每10年0.26 ℃的速率递增;年均降水量变化趋势不显著(P=0.780).
由于年轮径向生长的“滞后效应”[21-22],即长期的径向生长受到上一年温度、降水量的影响,序列敏感度也体现出树轮径向生长受前一年的影响,因此选取上一年生长季早期3月至当年生长季晚期10月的气候因子与树轮宽度年表进行相关分析,分析限制亚热带地区三明杉木径向生长的主要气候因子.
为阐明早材宽度、晚材宽度对整个年轮形成过程中的影响比重和指标间气候差异,采用SPSS 25.0中Pearson方法对3个径向生长指标年表进行相关分析.本研究采用树轮气候相关分析法,以DendroClim 2002软件[23]中Evolutionary and Moving Response and Correlation模块的向前滑动方式(窗口年限为18 a)进行滑动相关分析,检测在气候变暖背景下杉木径向生长与气候因子关系的稳定性.利用CANOCO 5.0软件中的冗余分析(RDA)方法,检验量化轮宽指数与气候因子的相关性.通过Excel 2010进行气候数据提取,采用ArcMap10.2、Origin2018作图.
3 结果与分析
3.1 杉木年表统计特征
由表1可知:3个杉木自回归年表平均敏感度为0.181~0.297,均大于0.1可信度水平,平均敏感度反映树轮对气候信息的敏感程度,表明三明地区杉木的径向生长变化包含较多气候信息且限制作用明显;年表的信噪比为29.088~35.002,样本总体代表性均大于0.85,表明年表可靠性高.
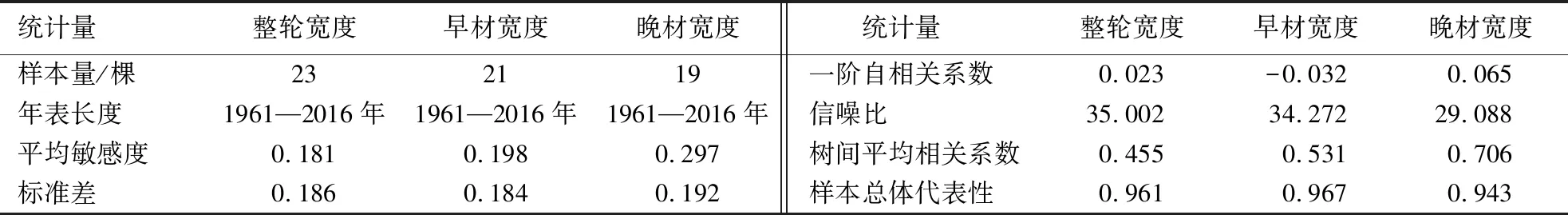
表1 杉木人工林ARS年表的统计特征Table 1 Statistics on the characteristics of ARS chronology

表2 杉木年轮指标间相关系数1)Table 2 Correlation coefficients of ring-related indices
1)*、**表示在0.05、0.01水平显著相关.
由表2可知,整轮宽度与早材宽度、晚材宽度之间存在极显著正相关,其中早材宽度与整轮宽度的相关系数为0.93,表明两者存在高度一致性.早材宽度与晚材宽度之间也存在极显著正相关.
3.2 杉木径向生长与气候因子的关系
杉木ARS年表与逐月气候因子的相关分析结果(图3)表明,不同径向生长指标均受到温度、降水的影响,但不同指标间响应程度存在一定差异.当年2月平均温度、平均最高温与早材宽度呈显著负相关,而与整轮宽度和晚材宽度呈负相关,未达显著水平;多数早材宽度或晚材宽度与气候因子相关的显著性水平大于整轮宽度与气候因子相关的显著性水平;其中当年7月最高温度与晚材宽度呈极显著负相关,而与早材宽度和整轮宽度呈负相关,未达显著水平;当年10月平均降水量、8月最高温度与晚材宽度呈显著正相关,而与早材宽度和整轮宽度呈正相关,未达显著水平.综上所述,说明在某一月份早材宽度或晚材宽度比整轮宽度指示的气候环境信息更准确.
整轮宽度和早材宽度对气候因子的响应存在较高一致性;当年9月平均温度、最低温度与晚材宽度呈极显著负相关,而与早材宽度和整轮宽度呈显著负相关.当年6月平均温度、8月最低温度与整轮宽度和早材宽度呈显著负相关,而与晚材宽度呈负相关,但未达显著水平.
杉木径向生长还受上一年气候要素的影响.其中,早材宽度与前一年12月平均降水量以及晚材宽度与前一年8月平均温度呈显著负相关,整轮宽度、早材宽度与前一年8月最低温度呈显著负相关.
3.3 杉木径向生长与气候因子的稳定性
选取当年2—10月温度、降水进行滑动相关分析,在气候变化条件下,检验杉木各径向生长指标与气候因子关系动态稳定性.结果(图4)表明,杉木径向生长3个指标均与当年9月平均最高温度呈显著负相关,并逐渐增强.整轮宽度与当年10月平均最高温度呈显著正相关,并逐渐减弱.晚材宽度与当年9月平均最低温度呈显著负相关,并逐渐增强,而与当年5月平均最低温度呈显著负相关,并逐渐减弱.
3.4 杉木径向生长与气候因子的冗余分析
杉木不同径向生长指标自回归年表与主要气候变量的冗余分析结果如图5所示.相关分析中显著水平较好的9个气候变量,在冗余分析中对杉木径向生长的影响均达到显著水平.当年7月平均最高温度和当年10月平均降水量对杉木径向生长的影响较强;其中,当年3、10月平均降水量与杉木径向生长呈显著正相关,而当年温度以及5月平均降水量与杉木径向生长呈显著负相关.晚材宽度与当年3月平均降水量呈极显著正相关,说明晚材宽度对早期生长季降水的影响存在“滞后效应”.
4 讨论与小结
4.1 杉木径向生长对气候因子的响应
在生长季气候因子对树木径向生长的影响较为复杂,树木旺盛的生命活动与旺盛的光合作用和呼吸作用紧密相连,气候因子成为影响净生产力的关键因素[24].研究表明树木径向生长主要受小环境温度和降水的干扰[6-8,25-26].
本研究发现当年2月平均温度和最高温度明显抑制早材径向生长,这可能是因为此时较高温度促进土壤水分蒸发、叶片蒸腾,并提高蒸气压而限制杉木的新陈代谢,不利于其径向生长[27].当年7月平均温度明显抑制晚材宽度径向生长,这可能是温度在7月份达到峰值(图1),其新陈代谢加速,而气孔导度和光合速率受到影响,不利于有机物储存和积累,从而限制径向生长,这与Yu et al[28]对华北麻栎(Quercusacutissma)研究的结论一致.因此证实了早、晚材宽度能够提取整轮宽度提取不到的气候环境信息,这与Souto-herrero et al[29]的研究结果一致.本研究还发现整轮宽度、早材宽度受上一年8月最低温度的明显抑制,可能是因为上一年生长季晚期较高的低温有利于呼吸作用,减少有机物储存,进而影响来年的径向生长养分供应[30].
本研究发现当年3月平均降水量不仅促进早材径向生长,而且对于晚材径向生长也有促进作用.一方面,可能是降水补充土壤水分,提高水分利用效率,有利于形成层细胞的分裂和伸长过程,使细胞个体增大[28],促进早材径向生长.另一方面,可能是降水有利于树木的光合作用和呼吸作用,在生长季前期积累养分和激素,进而影响后期晚材年轮径向生长的能力[29],表明晚材径向生长对当年生长季早期生长的影响存在“滞后效应”.当年10月平均降水量明显促进早材和晚材的径向生长,可能是因为生长季后期杉木形成层还没有完全停止活动,充足的水分有利于杉木的光合作用和新陈代谢,从而促进晚材生长[31];也可能是因为充足的水分有利于早、晚材管胞吸水,扩大了管胞体积[32].因此,通过综合分析早、晚材宽度能够提取杉木径向生长对气候环境的响应信息.
本研究还发现,杉木不同径向生长指标中早材宽度与整轮宽度的相关系数达到0.93,二者对气候因子的响应也呈高度一致性,说明早材宽度对整轮宽度的形成起着更重要的作用.这与兰彦平对北京地区板栗(Castaneamollisima)研究结果不一致[33],可能是因为杉木为速生树种,且南北气候存在差异.另外,冗余分析发现当年7月平均最高温度和当年10月平均降水量对杉木径向生长的影响较大,这可能是此区域在7月份温度达到最高水平,抑制了树木的光合作用,不利于径向生长.而在10月份生长季晚期较大的降水量提高了树木水分利用效率,促进树木径向生长.
4.2 杉木径向生长与气候因子关系的稳定性
研究发现,不同海拔树木径向生长与气候因子的响应关系的稳定性存在差异[22,32,34].本研究发现,杉木不同径向生长指标与气候因子关系的稳定性也不同,说明早材宽度、晚材宽度和整轮宽度不仅包含不同的气候环境信息,而且对气候因子的响应稳定性也有差异.
在气候变暖背景下,本研究发现杉木整轮宽度与当年10月平均最高温度呈显著正相关,并逐渐减弱.各径向生长指标与9月平均最高温度呈显著负相关,并逐渐增强.这可能是因为9月过高温度抑制光合作用,降低形成层活性,影响了细胞分化[11];而10月温度相对较低,适当提高温度有利于细胞分裂代谢,而气温呈明显的增长趋势,因此相关关系的稳定性发生变化.晚材宽度与当年10月平均降水量的正相关关系逐渐增强,这可能是因为在气候变暖背景下,秋季更加干旱,而降水有效缓解了干旱,促进晚材生长.并且较好的水分条件有利于延长树木晚材生长期.
此外本研究发现,杉木径向生长对当年生长季前期气候因子的响应敏感性较低,对生长季晚期气候因子的响应敏感性较强.这可能是因为杉木喜温暖湿润的气候环境,怕旱,当年3—7月温度逐渐升高降水充足,气候因子限制作用不明显.7月之后,降水显著减少,干旱逐渐增强(图1a),树木生长受到明显胁迫[13].而晚材宽度与当年5月平均最低温度呈显著负相关,并逐渐减弱.这可能是当年5月最低温限制了生长素的合成,而生长素的变化容易在树木生长季后期做出反应[9],随着温度升高,这种限制作用可能消失.
本研究结果表明:杉木不同径向生长指标受气候因素的影响,且存在明显差异;早材宽度和晚材宽度与其形成季节的气候因子存在显著相关.表明综合分析杉木整轮宽度、早材宽度和晚材宽度能够更加准确提取杉木径向生长对气候环境的响应信息.晚材生长对当年早材生长季气候因子存在“滞后效应”.在显著升温条件下,杉木径向生长的3个指标与气候因子的关系稳定性出现增强或减弱的变化趋势.
猜你喜欢
中国农业科技导报(2024年1期)2024-04-29 00:42:04
红蜻蜓·低年级(2021年12期)2022-01-19 05:18:24
天津医科大学学报(2021年1期)2021-12-05 11:11:05
小天使·一年级语数英综合(2018年7期)2018-09-12 10:13:26
滇池(2017年5期)2017-05-19 03:22:18
中国医学影像技术(2017年11期)2017-01-16 12:54:05
考试周刊(2016年63期)2016-08-15 14:33:26
故事作文·低年级(2016年5期)2016-05-14 08:45:49
西南农业学报(2016年6期)2016-04-16 05:13:01
科技视界(2016年1期)2016-03-30 14:08:41
