
小冰麦附加系高代材料中染色体的组成和变异研究
2020-01-03 05:54李亚莉董艳辉侯丽媛王育川吴新明
山西农业大学学报(自然科学版) 2019年6期
李亚莉,董艳辉,侯丽媛,王育川,吴新明
(1.山西省农业科学院 生物技术研究中心,山西 太原 030031;2.新疆生产建设兵团 第六师农业科学研究所,新疆 五家渠 831300)
天蓝冰草学名中间偃麦草(Thinopyrumintermedium),具有许多可供小麦育种利用的优良农艺性状,如耐寒、耐旱、抗真菌病和病毒病等[1~3],基因组原位杂交兼具直观、有效等优点,在小麦遗传育种研究中,该技术被广泛用于对小麦背景下外源遗传物质的追踪和鉴定[4~6]。目前,国内外已创制出许多小麦-天蓝冰草的中间材料, 即八倍体小冰麦、六倍体小冰麦和小冰麦异附加系[7]。我国小麦育种学家利用小冰麦异附加系进一步创制和鉴定了一批有利用价值的易位系材料,且育成优质小麦新品种[8]。小冰麦异附加系自创制以来到现在已有二十多年,对其后代材料先后从细胞和分子水平进行过检测和分析,讨论染色体的变异以及亲本的来源[9,10],但对于其高代材料中外源染色体的断裂和重组过程未见相关报道,为了进一步利用小冰麦材料改良小麦品种,本研究以小冰麦的高代材料为研究对象,即小冰麦异附加系 TAI系列,应用荧光原位杂交技术筛选有小片段插入、融合的后代,为创制异源新种质材料奠定基础。
1 材料和方法
1.1 试验材料
试验材料TAI系列(小冰麦异附加系),他们的天兰冰草染色体分别自异源八倍体小冰麦中2、中3、中4、中5所携带的天兰冰草染色体组,试验材料由中国科学院遗传所提供,试验于2017-2018年先后在中国科学院遗传所和山西省农科院进行。
1.2 试验方法
1.2.1 中期染色体的制备
根尖细胞中期染色体的制备、探针标记方法以及原位杂交程序均按照 Han等[8]描述的方法进行。首先,每份材料中分别选取约20粒种子,将挑选好的籽粒饱满的种子在垫有湿滤纸的培养皿中发芽,待根长长到 2~3 cm 时,剪取生长旺盛的根尖,首先根尖被果胶酶和纤维素酶的混合溶液37 ℃消化50 min,然后用70%的酒精洗2次,捣碎,离心,100% 的乙酸溶解,滴片。
1.2.2 原位杂交技术
植物总DNA的提取采用CTAB 法[11],利用微量紫外分光光度计检测样品 DNA浓度,并统一稀释至终浓度为 100 mg·L-1。
原位杂交程序按照Han等[11]描述的方法进行,在GISH分析中,提取中间偃麦草的基因组DNA,用fluorescein-12-dUTP标记为绿色,小麦的着丝粒逆转录转座子用Texas red-5-dCTP标记为红色,用中国春基因组DNA作为封阻;在FISH分析中,pAs1探针的信号标记为红色,pSc119.2 探针的信号标记为绿色;在Mc-GISH 分析中,染色体上绿色是中间偃麦草的信号,黄绿色是普通小麦A组的信号,蓝色是B组的信号,红色是D组的信号。细胞学镜检采用 OLYMPUSBX60荧光显微镜观察,用Photo-metrics SenSys CCD 成像。
2 结果与分析
2.1 TAI-I 和TAI-II系列高代材料染色体组成分析
对TAI-I 和TAI-II 系列的高代材料进行Mc-GISH分析,发现在TAI-I 系列的高代材料中小麦染色体条数为42,附加2条外源染色体;TAI-II 系列的高代材料中,TAI-23材料中小麦染色体条数是40,含有4条外源染色体,其余材料中均为小麦染色体条数是42,附加2条外源染色体(表1)。
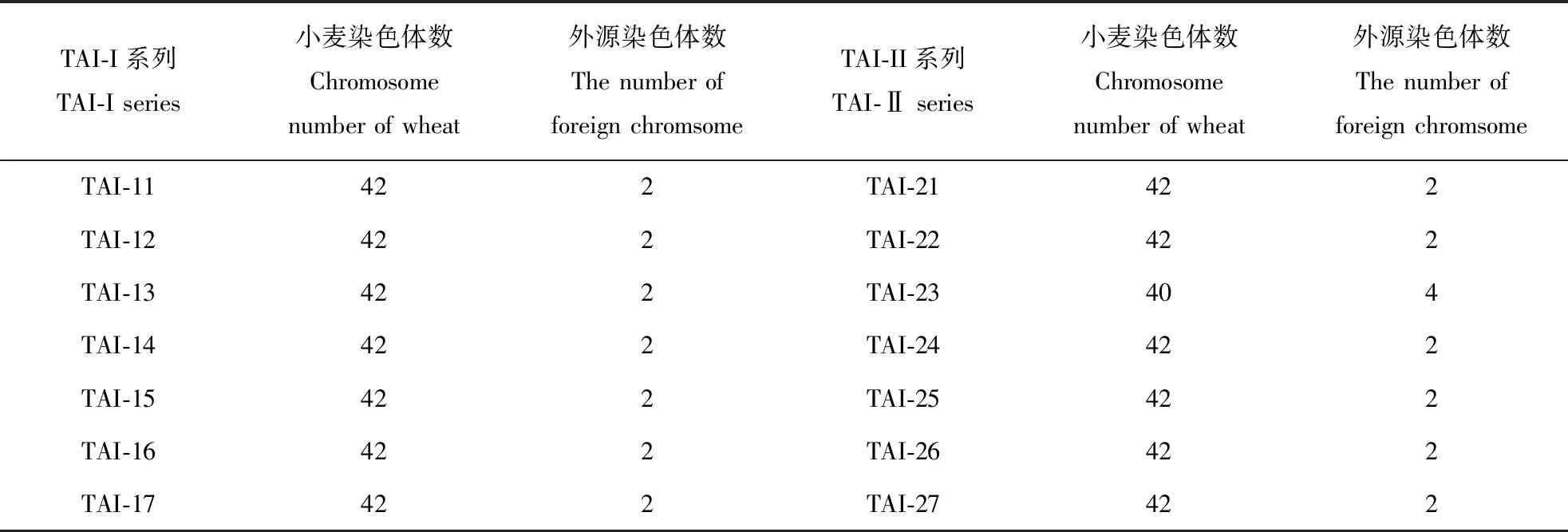
表1 TAI-I 和TAI-II 系列的高代材料染色体条数及其变异Table 1 The number of chromosomes and chromosomal changes in TAI-I and TAI-II series
材料创制过程中,我们将小冰麦的异附加系自交得到高代材料,始终保留能稳定遗传、籽粒饱满且染色体数在42条左右的后代连续种植,收集自交后代材料。本文应用荧光原位杂交技术对遗传稳定、籽粒饱满的高代材料进行检测,继续保留小麦染色体数为42的后代。检测结果发现大部分材料小麦染色体数稳定在42条,并附加了2条外源染色体,只有TAI-II系列的TAI-23包含小麦染色体数为40条,附加4条外源染色体。
2.2 TAI-I系列材料中外源染色体的变异
在TAI-I系列材料中,每份选取约20粒,首先进行GISH分析,观察小麦染色体和外源染色体的条数,发现所有材料小麦染色体均为42条,另附加2条外源染色体;之后,选取染色体分布清楚的制片,应用FISH 和 Mc-GISH技术,检测小冰麦附加系的高代材料中小麦和偃麦草染色体在重组过程中发生的染色体变异。
检测发现TAI-12、TAI-15、TAI-16三份材料中,小麦染色体和外源染色体均未发生任何改变,但是TAI-11和TAI-13两份材料中均附加两条B/D/Th.intermedium易位染色体,且TAI-13中2条6BS染色体丢失;TAI-17中附加2条D/Th.intermedium易位染色体,TAI-14附加两条端着丝粒染色体,染色体很短,几乎是其他染色体的一半,具体的染色体分布和变异如图1所示,箭头所指是发生变异的附加外源易位染色体。
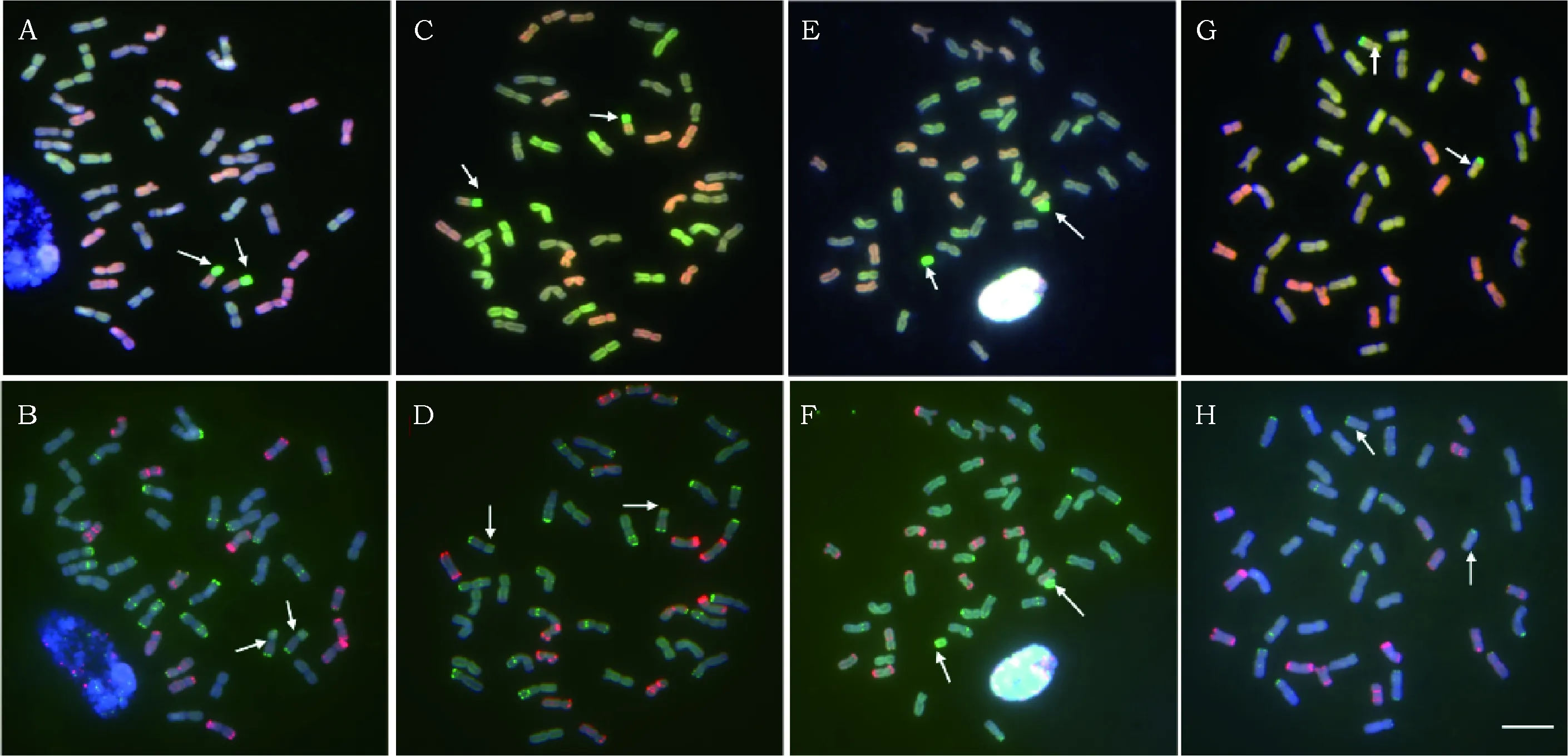
A,B,C,D,E,F,G,H分别是TAI11-3,TAI13-1,TAI14-2,TAI17-5的体细胞染色体分布模式,箭头指向发生断裂和重组的染色体。图A、C、E、G中,染色体上绿色是中间偃麦草的信号,黄绿色是普通小麦A组的信号,蓝色是B组的信号,红色是D组的信号;图B、D、F、H中,pAs1探针的信号标记为红色,pSc119.2探针的信号标记为绿色。Bar=10 μmPatterns of A and B areTAI11-3, patterns of C and D areTAI13-1, patterns of E and F are TAI14-2, patterns of G and H areTAI17-5, Arrows indicate the truncation and translocation chromosomes. In A、C、E、G, genome of Thinopyrum intermedium are labeled in green, the A genome DNA is labeled in yellow green, the D genome DNA is labeled in red, the B genome DNA is labeled in blue; In B、D、F、H, pAs1 probe is labeled in red, pSc119.2 probe is labeled in green. Bar=10 μm.图1 TAI11-3,TAI13-1,TAI14-2,TAI17-5的根尖中期细胞染色体的GISH和FISH分析Fig.1 GISH和FISH analysis of metaphase chromosomes in root-tip of TAI11-3 and TAI13-1,TAI14-2,TAI17-5
2.3 TAI-II系列材料中外源染色体的变异
TAI-II系列材料中,每份选取约20粒,首先进行GISH分析,观察小麦染色体和外源染色体的条数,发现所有材料小麦染色体大部分为42条,附加2条外源染色体,只有TAI-23和TAI-27两份材料不仅附加两条易位染色体,而且小麦染色体同时发生丢失和代换;之后选取染色体分布较好的制片,应用FISH 和 Mc-GISH技术,进一步检测TAI-II系列材料中所有染色体发生的变异。
检测发现TAI-21、TAI-24、TAI-25、TAI-26四份材料中,小麦染色体和外源染色体均未发生任何改变,即包含42条小麦染色体且附加2条偃麦草染色体,没有发现小麦和偃麦草的易位染色体。但是,TAI-22的材料中附加两条外源染色体,在杂交过程中,小麦和外源染色体发生易位,形成D/Th.intermedium易位染色体,且易位染色体从近着丝粒处断裂成小染色体;对小麦染色体进行检测分析发现,小麦的2 A染色体置换了5D染色体。TAI-23和TAI-27的材料中,小麦染色体40条,外源附加染色体4条,其中,TAI-23的材料中,小麦的6B染色体被中间偃麦草的染色体置换,小麦的3BS丢失,附加2条D/Th.intermedium易位染色体;TAI-27的材料中,2条D/Th.intermedium易位染色体从近着丝粒处断裂成小染色体,小麦的5D染色体被2 A染色体置换。 具体的染色体分布和变异如图2所示,箭头所指的是附加的易位染色体。
3 讨论
小麦和偃麦草的远缘杂交高代材料中,外源染色体会发生变异,形成易位染色体或者断裂形成小染色体[12]。TAI系列是为了导入偃麦草的优良抗性基因,通过远缘杂交创制的育种中间材料,通常包含42条小麦染色体和2条中间偃麦草染色体[13]. 之后对这些材料进行了更深入的细胞学检测,所有材料在鉴定过程中发现不仅外源染色体和小麦染色体发生交换、重组,小麦染色体的不同亚组内也发生染色体丢失、代换的现象。同时,由于染色体在分离、重组过程中发生错分裂,导致染色体断裂、重组,在TAI-14、 TAI-22和 TAI-27的亲本材料中,均发现了端粒染色体和小染色体[14,15]。
本研究应用GISH, FISH 和 Mc-GISH 技术, 继续筛选TAI系列的后代材料,发现TAI-12、TAI-15、TAI-16、TAI-21、TAI-24、TAI-25、TAI-26附加的2条中间偃麦草染色体仍然没有发生断裂、重组等变异,其它材料中外源染色体和小麦染色体发生交换、重组,新形成的易位染色体又发生整臂断裂等现象,这些染色体变异主要是由于减数分裂期间,中间偃麦草染色体发生错分裂产生小染色体,或者产生的端着丝粒染色体和小麦染色体发生融合,形成易位染色体[16,17],而新形成的易位染色体不稳定,TAI-22材料在着丝粒区域发生断裂整臂丢失(图2 M)。对比所有TAI系列材料的染色体组成发现,有些材料后代比较稳定,虽附加2条外源染色体,但是小麦和偃麦草染色体均能正确分离和配对,后代比较稳定,但是,有些材料随着繁育代数的增加,外源染色体会发生断裂,然后和小麦染色体重组,形成易位染色体。
4 结论
综上所述,本研究对TAI 系列的材料进行了更深入的细胞学检测和分析,发现更多的染色体变异情况,包括小麦A、B、D之间的代换和丢失;同时,发现不同材料随着代数的增加,染色体组成也会发生不同程度的变异,下一步将继续鉴定附加染色体的亚型,并考察育成小冰麦TAI材料的农艺性状,为改良小麦品种筛选优良的育种中间材料。
猜你喜欢
中国卫生标准管理(2022年21期)2023-01-03
作物学报(2022年2期)2022-11-06
舰船科学技术(2022年11期)2022-07-15
农业资源与环境学报(2021年5期)2021-10-06
快乐语文(2020年14期)2020-07-04
现代园艺(2018年3期)2018-02-10
上海农业学报(2017年3期)2017-04-10
西部(2016年6期)2016-05-14
今日农药(2016年5期)2016-05-14
医学研究杂志(2015年2期)2015-06-10
