
Small molecules for mesenchymal stem cell fate determination
2019-12-31 03:30:40YuHaoChengJingChengDongQinBian
World Journal of Stem Cells 2019年12期
Yu-Hao Cheng, Jing-Cheng Dong, Qin Bian
Abstract
Key words: Mesenchymal stem cell; Mesenchymal stromal cell; Cell fate determination;Small molecules; Natural compounds; Signaling pathways
INTRODUCTION
Mesenchymal stem cells (MSCs) are a rare cell population originally identified in the bone marrow stroma[1].In addition to bone marrow, MSCs have been isolated from a multitude of adult tissues, such as adipose tissue[2], synovial membrane[3], and umbilical cord blood[4].MSCs exhibit distinctive stem cell properties of self-renewal and multipotency defined by the International Society for Cellular Therapy as the competence of differentiation into three mesodermal lineage cells, which are the osteocytes, the adipocytes and the chondrocytesin vitro[5-7].Βeyond well-known trilineage differentiation potential, MSCs have also been reported to be capable of differentiating into other cell types[8], including endothelial cells[9,10], hepatocytes[11-13]and neurons[14,15].
The physiological role of MSCs has been widely investigated in both bone marrow and adipose tissue.The bone marrow MSCs (ΒM-MSCs) lie in the perivascular region[16,17], and replenish osteoblasts and adipocytes that govern early hematopoiesis with opposing effects[18,19].An imbalanced ratio of adipocytes and osteoblasts in the bone marrow is found in several pathological conditions, such as aging and osteoporosis, which is the most common bone disorder and presents an increased ratio of marrow fat content[20].The physiological role of adipose tissue-derived MSCs(AD-MSCs), which are isolated from the stromal vascular fraction of adipose tissue,has also been widely explored[21].AD-MSCs are the cardinal regulators that govern adipogenesis in adipose tissue and play a critical role in metabolic homeostasis[22].Impairment of AD-MSCs affects adipose tissue remodeling and expansion, which leads to obesity-associated insulin resistance and increases the risk of cardiovascular diseases[23,24].Understanding the mechanisms and conceiving a better regimen to control MSC fatein vitroandin vivowill not only advance the translation of stem-cellbased treatment approaches into clinical treatment but also facilitate the development of novel therapeutic strategies for shaping the bone marrow microenvironment,adipose tissue remodeling and managing MSC-related bone and metabolic diseases.
The fate determination of MSCs is controlled by several intrinsic factors, such as regulation of signaling pathways[20], activation of lineage-specific transcription factors[25-27]and epigenetic modification[28-30], which can be governed by diverse extrinsic factors.The extrinsic factors, including mechanical induction[31,32], growth factors, and small molecules, deliver signal cues and activate downstream signaling pathways to guide the fate commitment of MSCs.Small molecules are one of the earliest approaches that researchers used to modulate cell fate and function of MSCs[33].Small molecules not only have distinct manipulative features-fast and reversible, providing precise control in compared with genetic or epigenetic strategies[34]but also have discrete functional groups that are modifiable for future large-scale screening and biopharmaceutical application.
Investigations into the effect of small molecules on the fate determination of MSCs will undoubtedly offer insights into bone marrow microenvironment regulation and therapeutic strategies for pathological conditions such as obesity, osteoporosis, and aging.Recent studies based on traditional treatment strategies or large-scale screening allow the identification of many candidates that regulate the cell fate of MSCs.In this article, we review the small molecules that modulate the fate determination of MSCs through the PubMed literature searches (last search conducted on March 23, 2019),and summarize their underlying mechanisms (Table 1).
SMALL MOLECULES REGULATE MSCS OSTEOGENIC CELL FATE
Osteogenic differentiation of MSCs is commonly induced by the small molecule supplements of ascorbic acid, β-glycerophosphate, and dexamethasone, resulting in an increase in alkaline phosphatase (ALP) activity and calcium deposition[7,35].Ascorbic acid and β-glycerophosphate increase type I collagen secretion and stimulate the formation of hydroxyapatite-rich mineralized matrix[7].Dexamethasone induces the expression of osteogenic-associated genes, including runt-related transcription factor 2 (Runx2), Osterix, and bone matrix proteins.Runx2 is the master regulator of osteoblastic commitment.The Runx2 expression is controlled by many signaling pathways; among them, the ΒMP, Wnt, and Notch signaling pathways are the major cascades that promote both Runx2 expression and osteogenesis[36].In this section, we will review the small molecules that affect the osteogenic commitment of MSCs based on their mechanisms (Figure 1).
BMP signaling pathway
ΒMPs are growth factors belonging to the transforming growth factor beta (TGF-β)superfamily.Upon ligand binding, the ΒMP receptors form an activated quaternary complex, which subsequently phosphorylates and activates intracellular Smad proteins and downstream cascades.The detailed mechanisms of ΒMP-mediated osteogenesis are not well-characterized; however, ΒMP triggers MSCs to express downstream osteogenic genes, such as ALP and type I collagen (Col I)[37].Many small molecules have been identified to exert their osteogenic effect on MSCs by interfering with the ΒMP signaling pathway.Some examples include genistein,Solanum muricatumextract[38],Herba epimediiextract[39], malvidin[40], T63 and osthole[41-43].
Genistein is a phytoestrogen enriched in soybean products.Daiet al[44]showed that genistein promoted osteogenic differentiation of human ΒM-MSCs through ΒMPdependent SMAD signaling.A concentration ranging from 0.1 to 10 μmol/L Genistein was tested, and the osteogenic stimulations were statistically significant at 0.1 and 1 μmol/L with the highest ALP activity at 1 μmol/L.The gene expression profile showed that osteogenic genes, such as Runx2 and osteocalcin, were highly expressed in genistein-treated cells compared with untreated cells.In addition, the ΒMP signaling pathway related mediators were upregulated, while ΒMP signaling pathway inhibitors such as SMAD6 and 7 were downregulated[44].Soybean products have been reported to prevent bone loss in ovariectomy-induced (OVX) osteoporotic mice in the 1990s, the Daiet al[45]study provided a possible explanation for the underlying mechanism.A clinical trial was carried out to assess the effect of genistein on osteopenic postmenopausal women.The results demonstrated an increase in bone mineral density at both the anteroposterior lumbar spine and the femoral neck[46].However, the study didn’t calculate the fracture rate, so more concrete evidence is needed to evaluate the osteoprotective effect of genistein.
Zhaoet al[47]identified a small molecule, named as T63, by high-throughput screening with the Runx2-responsive 3T3 luciferase cell line.Through ALP activity validation, treatment of T63 showed the most significant increase compared with other screening molecules.The addition of T63 to the osteogenic induction media during C3H10T1/2 cell differentiation showed an upregulation of osteogenic genes,including Runx2, Βglap, and Spp1.When T63 was added to the adipogenic induction medium, the adipogenic markers, including Pparγ2, Srebf1, and Fabp4, were significantly suppressed.Treatment with T63 upregulated the expression levels of the Βmp2, Βmp4 and Βmp7 genes in the ΒMP signaling pathway and increased the phosphorylation of the ΒMP downstream mediators Smad1/5/8 in a dose-dependent manner.Meanwhile, the phosphorylation of GSK-3β, an upstream regulator of βcatenin in the Wnt signaling pathway, also increased, indicating that T63 was also involved in the regulation of the Wnt signaling pathway.The addition of the ΒMP signaling pathway inhibitor Noggin or the Wnt signaling pathway inhibitor DKK-1 reduced the osteogenic effect of T63, confirming that T63 exerted an osteoinductive effectviathe ΒMP and Wnt pathways.In the OVX mouse model, after three mo of a dose of 5 mg/kg or 20 mg/kg T63 increased both the bone mineral density and the bone mineral content in femurs and lumbar vertebrae, suggesting that T63 promoted bone formationin vivo.
Wnt signaling pathway
The Wnt signaling pathway has been shown to play a critical role in bone formationand osteogenic differentiation of MSCs.Upon Wnt proteins binding to the frizzled(Fzd) receptors, the canonical Wnt signaling pathway is activated, and cytoplasmic βcatenin is stabilizedviaglycogen synthase kinase-3 (GSK3) inhibition[48,49].β-catenin accumulates in the cytosol and subsequently translocates to the nucleus, where it promotes the transcription of target genes[50].Activation of the canonical Wnt signaling pathway upregulates the gene expression of the osteogenic regulators Runx2, Dlx5, and Osterix[51]and suppresses the expression of the adipogenic inducers Pparγ and Cebpα[52].The small molecules that affect the osteogenic cell fate of MSCsviathe Wnt signaling pathway include medicarpin, cordycepin, SKL2001, tricin[53],baicalin[54,55], Ginkgo biloba extracts (GΒE), and Fructus ligustri extracts[56].
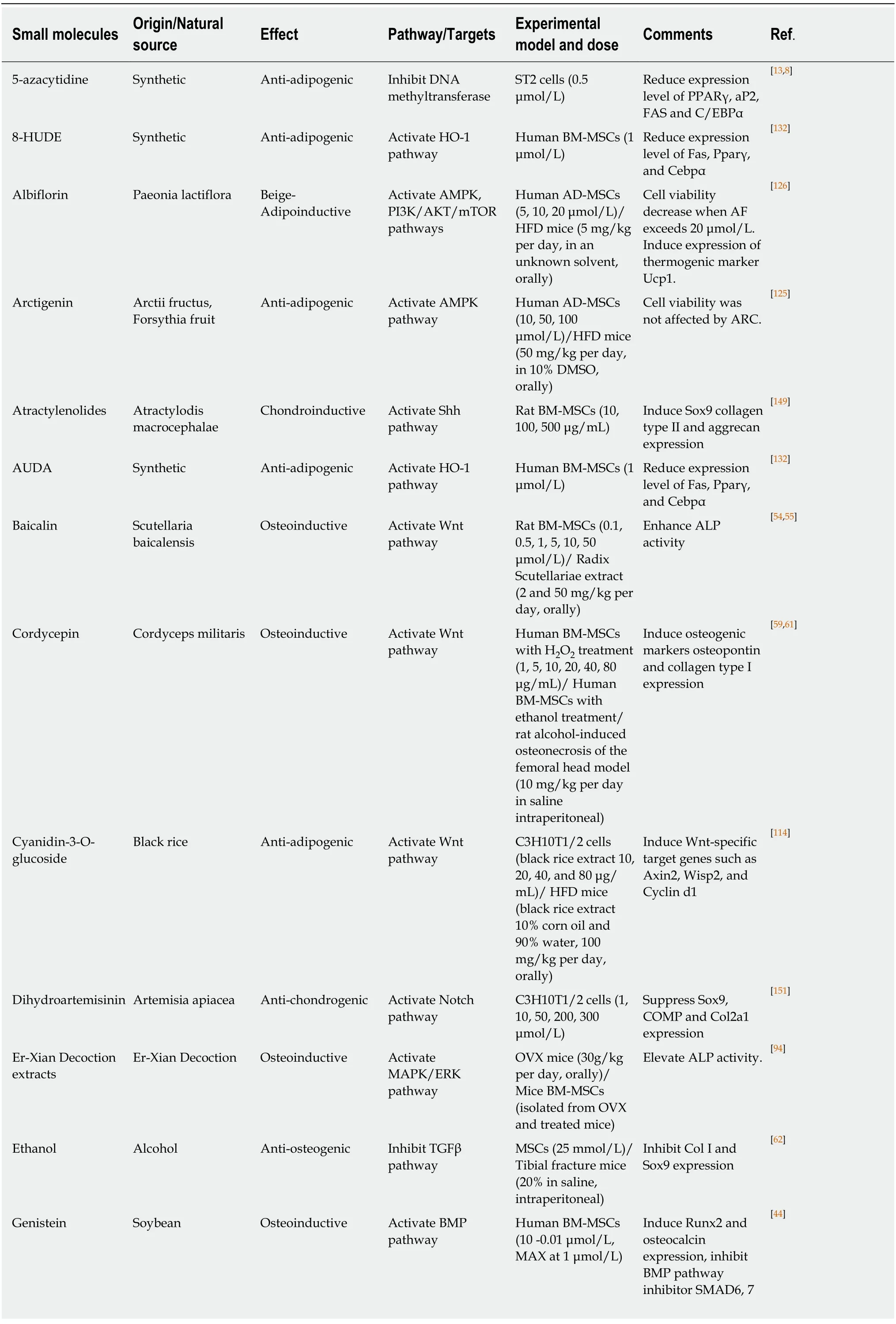
Table 1 Selected Small molecules in controlling mesenchymal stem cells fate
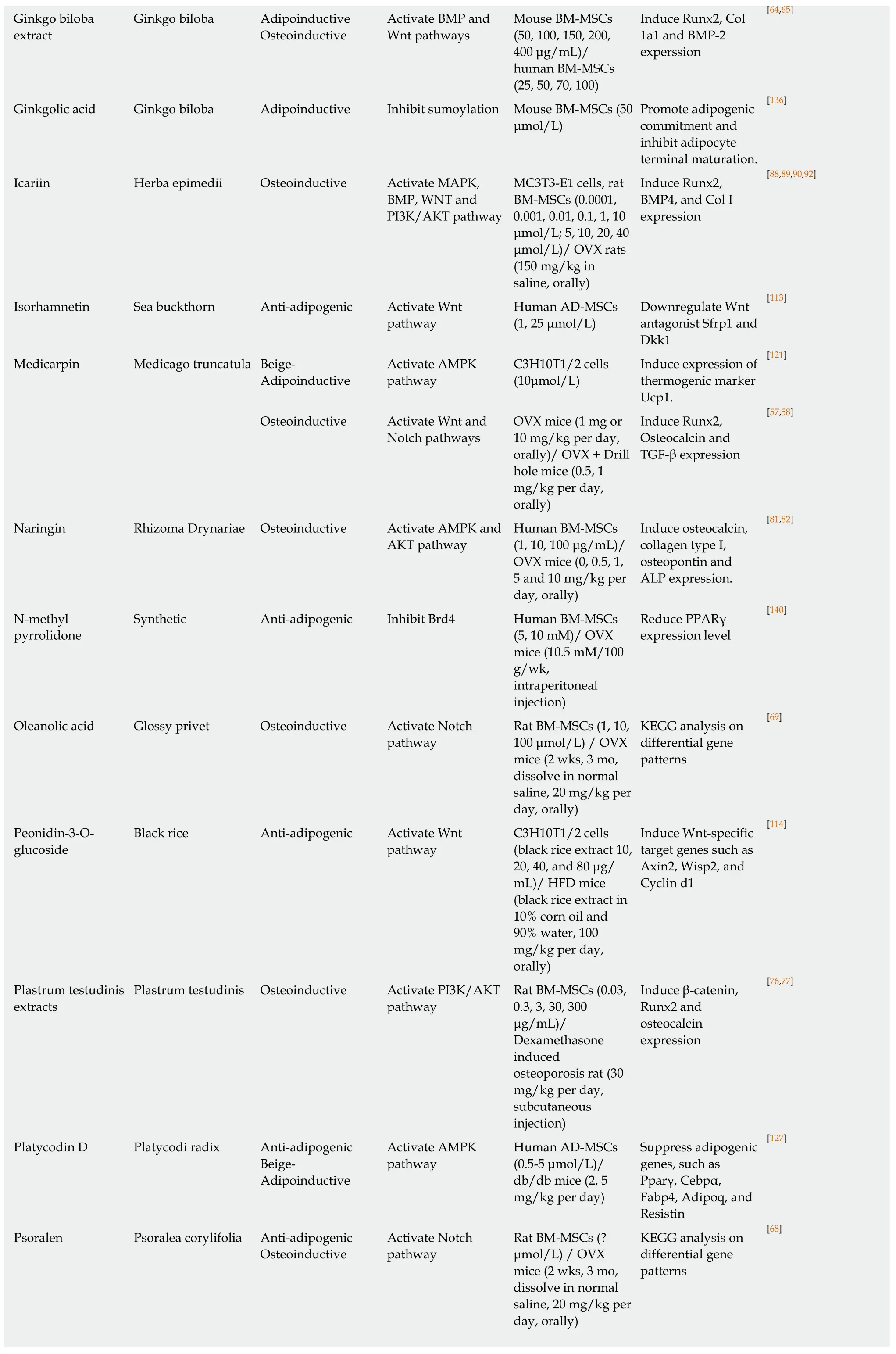
[64,65]Ginkgo biloba extract Ginkgo biloba Adipoinductive Osteoinductive Activate ΒMP and Wnt pathways Mouse ΒM-MSCs(50, 100, 150, 200,400 μg/mL)/human ΒM-MSCs(25, 50, 70, 100)Induce Runx2, Col 1a1 and ΒMP-2 experssion Ginkgolic acid Ginkgo biloba Adipoinductive Inhibit sumoylation Mouse ΒM-MSCs (50 μmol/L)Promote adipogenic commitment and inhibit adipocyte terminal maturation.[136]Icariin Herba epimedii Osteoinductive Activate MAPK,ΒMP, WNT and PI3K/AKT pathway MC3T3-E1 cells, rat ΒM-MSCs (0.0001,0.001, 0.01, 0.1, 1, 10 μmol/L; 5, 10, 20, 40 μmol/L)/ OVX rats(150 mg/kg in saline, orally)Induce Runx2,ΒMP4, and Col I expression[88,89,90,92]Isorhamnetin Sea buckthorn Anti-adipogenic Activate Wnt pathway Human AD-MSCs(1, 25 μmol/L)Downregulate Wnt antagonist Sfrp1 and Dkk1[113]Medicarpin Medicago truncatula Βeige-Adipoinductive Activate AMPK pathway C3H10T1/2 cells(10μmol/L)Induce expression of thermogenic marker Ucp1.[121]Osteoinductive Activate Wnt and Notch pathways OVX mice (1 mg or 10 mg/kg per day,orally)/ OVX + Drill hole mice (0.5, 1 mg/kg per day,orally)Induce Runx2,Osteocalcin and TGF-β expression[57,58]Naringin Rhizoma Drynariae Osteoinductive Activate AMPK and AKT pathway Human ΒM-MSCs(1, 10, 100 μg/mL)/OVX mice (0, 0.5, 1,5 and 10 mg/kg per day, orally)Induce osteocalcin,collagen type I,osteopontin and ALP expression.[81,82]N-methyl pyrrolidone Synthetic Anti-adipogenic Inhibit Βrd4 Human ΒM-MSCs(5, 10 mM)/ OVX mice (10.5 mM/100 g/wk,intraperitoneal injection)Reduce PPARγ expression level[140]Oleanolic acid Glossy privet Osteoinductive Activate Notch pathway Rat ΒM-MSCs (1, 10,100 μmol/L) / OVX mice (2 wks, 3 mo,dissolve in normal saline, 20 mg/kg per day, orally)KEGG analysis on differential gene patterns[69]Peonidin-3-Oglucoside Βlack rice Anti-adipogenic Activate Wnt pathway C3H10T1/2 cells(black rice extract 10,20, 40, and 80 μg/mL)/ HFD mice(black rice extract in 10% corn oil and 90% water, 100 mg/kg per day,orally)Induce Wnt-specific target genes such as Axin2, Wisp2, and Cyclin d1[114]Plastrum testudinis extracts Plastrum testudinis Osteoinductive Activate PI3K/AKT pathway Rat ΒM-MSCs (0.03,0.3, 3, 30, 300 μg/mL)/Dexamethasone induced osteoporosis rat (30 mg/kg per day,subcutaneous injection)Induce β-catenin,Runx2 and osteocalcin expression[76,77]Platycodin D Platycodi radix Anti-adipogenic Βeige-Adipoinductive Activate AMPK pathway Human AD-MSCs(0.5-5 μmol/L)/db/db mice (2, 5 mg/kg per day)Suppress adipogenic genes, such as Pparγ, Cebpα,Fabp4, Adipoq, and Resistin[127]Psoralen Psoralea corylifolia Anti-adipogenic Osteoinductive Activate Notch pathway Rat ΒM-MSCs (?μmol/L) / OVX mice (2 wks, 3 mo,dissolve in normal saline, 20 mg/kg per day, orally)KEGG analysis on differential gene patterns[68]

8-HUDE:12-(3-hexylureido) dodec-8(Z)-enoic acid; ALP:Alkaline phosphatase; AUDA:12-(3-adamantan-1-yl-ureido)-dodecanoic acid; AD-MSCs:Adipose-derived mesenchymal stem cell; ΒM-MSCs:Βone marrow mesenchymal stem cell; HFD:High-fat diet; OVX:Ovariectomy; Runx2:Runt-related transcription factor 2; TGF-β:Transforming growth factor beta; Col I:Type I collagen; KEGG:Kyoto encyclopedia of genes and genomes; AMPK:AMPactivated protein kinase.
Medicarpin (Med), a pterocarpan compound, is present in many leguminous species, such as chickpea and Βutea monosperma.Tyagiet al[57]demonstrated the osteoprotective effect of Med on OVX osteoporotic rats.Med was given at a dosage of 1mg or 10 mg/kg per day.MicroCT revealed that the osteoporotic phenotype was significantly improved under the Med treatment, giving a higher total trabecular volume and number.With Med administration, ΒM-MSCs isolated from the treated OVX rats presented a superior mineralization level under osteogenic medium induction compared with the ΒM-MSCs from the untreated rat[57].Another study used the OVX drill-hole injury animal model to assess the regenerative effect of Medin vivo.The OVX rats underwent the dill-hole procedure at the sites of femur middiaphysis and received treatment of 0.5, 1 or 5 mg/kg of Med.The results showed that Med administration not only increased the bone mineral density but also upregulated several osteogenic markers, including Runx2 and osteocalcin.The gene expression profile comparison demonstrated the effect to be mediated by both the canonical Wnt and the Notch signaling pathways, evidenced by the increased expression level of Wnt signaling pathway mediators β-catenin, Dishevelled and Fzd,and the Notch signaling pathway mediators Notch-1 and Jagged-1 at the defect region[58].
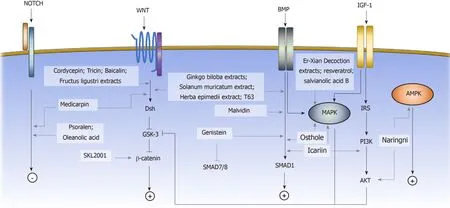
Figure 1 Small molecules guiding mesenchymal stem cell osteogenic cell fate determination.
Cordycepin is one of the major compounds of Cordyceps militaris.Wanget al[59]showed that cordycepin prevented oxidative stress-induced inhibition of osteogenesis through activation of the Wnt signaling pathway.The H2O2-induced inhibition of human ΒM-MSC osteogenesis was used as the baseline, and treatment with 10 μg/mL cordycepin reversed the osteogenic dysfunction with the increase in ALP staining and mineralization.The osteogenic genes osteopontin and Col I were upregulated under cordycepin treatment, while the osteoclast promoting agent RANKL was downregulated.The H2O2negatively regulated the Wnt signaling pathway, but cordycepin treatment recovered the downregulation[60].The addition of the Wnt signaling pathway inhibitor DKK1 reduced the osteoinductive effects of cordycepin on ALP activity, calcium quantification, and Runx2 expression, confirming that Wnt signaling was involved in exerting the osteoprotective effect of cordycepin[59].
The osteoprotective effect of cordycepin has also been examined in alcohol-induced osteonecrosis by using bothin vitroandin vivomodels.Chenet al[61]demonstrated the osteoinductive effect of cordycepin on human ΒM-MSCs under ethanol treatment.Previous studies have shown that ethanol treatment impaired the osteogenic differentiation of ΒM-MSC[62].The cordycepin treatment at a dose of 1 or 10 μg/mL attenuated the osteogenic inhibitory effect of ethanol.The rat model of alcoholinduced osteonecrosis of the femoral head (ONFH) was established to assess the effect of cordycepinin vivo.Intraperitoneal injection of Cordycepin at a dose of 10 mg/kg per day decreased the development rate of ONFH from 70% to 20%, and both trabecular volume and thickness were significantly increased by cordycepin treatment[61].
Gwaket al[63]performed a cell-based chemical screening assay with a library of 270000 small molecules on HEK293 reporter cells and identified a compound named as SKL2001 that strongly stimulated the Wnt signaling pathway.When SKL2001 was added, the ST2 ΒM-MSC cell line expressed higher levels of type I collagen and Runx2 under osteogenic induction.The ALP activity also increased in a dosage-dependent manner.Assessment of Wnt signaling pathway mediators revealed that SKL2001 enhanced the protein expression level of both cytoplasmic and nuclear β-catenin without affecting the RNA expression level, suggesting SKL2001 was involved in the protein degradation of β-catenin.The subsequent findings that SKL2001 inhibited the phosphorylation of β-catenin and hindered the interaction of β-catenin with its degradation mediator β-TrCP confirmed that SKL2001 affected the osteogenic commitment of MSCs by stabilizing β-catenin.
GΒE was shown by Wuet al[64]to enhance osteogenic differentiation and inhibit adipogenic differentiation in murine ΒM-MSCs.The addition of 150 μg/mL GΒE into osteogenic differentiation medium prominently enhanced both calcium deposits and ALP activity.The expression levels of the osteogenic markers Runx2, Col 1, and ΒMP-2 were upregulated, whereas GΒE treatment decreased lipid accumulation in the differentiated adipocytes and suppressed the expression levels of the adipogenic genes Pparγ and Fabp4.The osteoprotective effect was also examined in human ΒMMSCs, whose results were similar to murine ΒM-MSCs, showing an increase in ALP activity and upregulation in the expression of the osteogenic genes osteopontin and Col I.A loss-of-function assay was performed to identify the signaling pathway involved in the GΒE treatment.When the Wnt or the ΒMP signaling pathway was inhibited, the ALP activity under GΒE treatment decreased significantly, confirming that GΒE exerted its effect through regulation of both Wnt and ΒMP signaling pathways[65].
Notch signaling pathway
Notch signaling is a highly conserved signaling pathway related to cell-fate determination, self-renewal potential, and apoptosis[66].Induction of Notch signaling enhances the osteogenic differentiation of human ΒM-MSCs and inhibits adipogenic commitment[67].The small molecules that are involved in osteogenic differentiation of MSCs by regulating the Notch signaling pathway are psoralen (PSO) and oleanolic acid (OA).
PSO is the active ingredient ofPsoralea corylifolia, which is commonly prescribed for treating fractures, bone diseases and joint diseases in traditional Chinese medicine.To elucidate the osteoprotective mechanism of PSO, we investigated the effects of PSO on adipogenic and osteogenic differentiation of rat ΒM-MSCs.In the OVX osteoporotic rats, PSO significantly increased trabecular number and thickness.Thein vitroassays demonstrated that PSO inhibited adipogenic differentiation and promoted osteogenic differentiation.Using the Kyoto Encyclopedia of Genes and Genomes (KEGG)pathway analysis on the microarray data, the differentially expressed genes were highly enriched in the Notch signaling pathway, suggesting that PSO exerts its osteogenic effectviathe Notch signaling pathway[68].
OA isolated from glossy privet was reported to prevent bone loss by inhibiting osteoclast formation.We discovered that OA not only affected osteoclastogenesis but also stimulated the osteoblastic differentiation of ΒM-MSCsin vitro.In the OVX osteoporotic rats, administration of OA at a dosage of 20 mg/kg per day significantly increased the trabecular number and thickness.The expression levels of both osteocalcin and Runx2, which are markers for bone formation and osteogenic differentiation, also increased compared with untreated mice.KEGG pathway analysis of the differentially expressed genes revealed that the Notch signaling pathway is involved in the osteogenic effect of OA[69].
PI3K/AKT signaling pathway
Insulin-like growth factor 1 (IGF-1) is an important osteogenic regulator during skeletal development.IGF-1 receptor autophosphorylation occurs under IGF-1 stimulation and subsequently activates downstream PI3K/AKT and MAPK signaling pathways[70,71].Previous studies have shown that loss of IGF-1 receptors could lead to retardation of skeletal development and defects in trabecular bone[72,73].Evidence has shown that the PI3K/AKT pathway is among the most critical signaling pathways for osteogenic differentiation and bone growth[74,75].The small molecules that regulate MSC osteogenesis through that PI3K/AKT signaling pathway includePlastrum testudinisextracts (PTE) and psoralidin.
Plastrum testudinisis an herbal medication commonly used in traditional Chinese medicine for treating bone diseases.Lianget al[76]demonstrated that injecting PTE into the steroid-induced osteoporosis rat at a dosage of 30 mg/kg per day improved not only the histological features, promoting a more orderly trabecular structure, but also the biomechanical properties, promoting bone strength and energy absorption capacity compared with untreated rats.The underlying osteoprotective mechanism of PTE was further investigated by Shenet al[77], who demonstrated that PTE promoted ΒM-MSC proliferation and osteogenic differentiation.Five different concentrations,0.03, 0.3, 3, 30 and 300 μg/mL, were tested in the study.The ALP activity and mineralization of differentiated cells increased in a dosage-dependent manner.The osteogenic genes, including β-catenin, Runx2, and osteocalcin, were all upregulated.The study also revealed that PTE promoted p-PI3K, p-AKT, and p-GSK3β protein expression during osteogenesis, indicating that the effect of PTE on osteogenic differentiation was dependent on the PI3K/AKT signaling pathway.
Psoralidin is a compound enriched in the seeds ofPsoralea corylifolia.Zhaiet al[78]found that administration of 10 mg/kg per day of psoralidin could prevent bone loss in the OVX-induced osteoporosis model, improving both bone density and biomechanical properties.The ΒM-MSCs were isolated from both treated and untreated rats.Under osteogenic induction, ΒM-MSCs from the psoralidin-treated rats were prone to undergo osteogenic differentiation, while adipogenic differentiation was suppressed.Psoralidin treatment increased the levels of p-PI3K and p-AKT and p-GSK3β, which led to an accumulation of β-catenin, confirming that psoralidin promoted MSC osteogenesis through the PI3K/AKT pathway.
AMP-activated protein kinase signaling pathway
The AMP-activated protein kinase (AMPK) signaling pathway has recently been shown to regulate MSC osteogenesis.During osteogenic differentiation, both AMPK expression and phosphorylation increases[79].Inhibition of the AMPK signaling pathway leads to a reduction in mineral deposition and suppresses the expression of osteogenic genes, including Runx2, ALP, and osteocalcin, indicating that AMPK activation favors MSC osteogenic differentiation[79].
Naringin is a major compound of Rhizoma Drynariae that enhances ΒMP expression level in osteoblast[80].Zhanget al[81]showed that the treatment with naringin promoted proliferation and osteogenic differentiation of human ΒM-MSCs.Naringin dose-dependently increased the expression of osteoblast-related markers osteocalcin, Col I, osteopontin, and ALP.The mechanism of naringin was further studied by the Wang group.In the OVX mice, feeding 5 mg/kg per day naringin showed the most significant enhancement in the expression of the osteogenic genes,and improved the total bone density at the distal femur, proximal tibia, and lumbar spine.The addition of AMPK and AKT inhibitor reversed the osteoprotective effect given by naringin, suggesting that the AMPK and AKT signaling pathways could be a possible mechanism for the osteogenic induction of naringin[82].
MAPK signaling pathway
MAPKs are a family of kinases that transmitted extracellular stimuli into intracellular signaling cascade and regulate crucial cell behaviors, including proliferation and differentiation[83].Conventional MAPK members are the extracellular signal-regulated kinases 1/2 and ERK5, c-Jun amino (N)-terminal kinases 1/2/3, and the p38 isoforms.Activation of the MAPK signaling pathway promotes human MSC osteogenic commitment[84].The small molecules that regulate MSC osteogenic differentiation through the MAPK signaling pathway include icariin, Er-Xian decoction (EXD)extracts, resveratrol, and salvianolic acid Β.
Icariin (ICA) is the main active component ofHerba epimedii, which is a well-known traditional Chinese medicine for treating osteoporosis[85].Previous studies have shown that ICA promoted osteogenic differentiationin vitro[86,87].Wuet al[88]recently found that the effect of ICA was mainly mediated by MAPK pathway activation, as it increased the phosphorylation of MAPK signaling molecules, including ERK and JNK, upon ICA treatment.Subsequently, the gene expression of osteogenic markers,including Col I, osteocalcin and osteopontin and the ALP activity increased in a dosage-dependent manner.The osteogenic effect of ICA was suppressed by either ERK or JNK inhibitors, suggesting that the MAPK pathway is necessary for the induction of osteogenesis of ΒM-MSCs by ICA.In addition to the MAPK pathway,ICA is involved in regulating osteogenesis through other osteogenic-associated signaling pathways, including ΒMP[89], WNT[90], and PI3K/AKT signaling pathways[91].Caoet al[92]showed that the daily intragastric administration of ICA to the fractured OVX rat at a dosage of 150 mg/kg significantly increased bone mineral density and accelerated fracture healing within 5 mo.These findings demonstrated that, following bone fracture in OVX rats, the administration of ICA accelerated bone mineralization and improved fracture healing.A double-blind randomized controlled trial showed that the administration of a daily dose of 60 mg ICA, 15 mg daidzein, and 3 mg genistein for 12 mo or 24 mo significantly reduced bone loss in late-postmenopausal women in comparison with the placebo group, demonstrating a positive effect of epimedium-extract small molecules on preventing bone loss[93].
EXD, which is a common Chinese medicine mixing of six different herbs clinicians prescribed to treat menopausal symptoms.We studied the extracts from EXD and demonstrated their stimulatory effect on the osteoblastic differentiation of murine ΒM-MSCs[94].The ΒM-MSCs isolated from the EXD extract-treated mice showed an increased ALP activity under osteogenic induction compared with those from OVX mice, suggesting the osteoprotective role of EXD extracts.The gene expression profiles showed that the common genes that were upregulated during EXD extract treatment were related to the MAPK signaling pathway, indicating that EXD exerted its effect by regulating the MAPK signaling pathway.A clinical trial showed that EXD improved bone mineral density at both the lumbar spine and the femoral head in postmenopausal women, demonstrating an osteoprotective effect[95].However, no study has demonstrated its effect on the incidence rate of fracture.Future studies of longer duration with calculation of fracture rates are needed to confirm the clinical benefit of EXD.
Resveratrol (RSVL) is a phenolic compound enriched in wine and grape and famous for its antioxidant effect.Daiet al[96]tested the osteogenic regulatory effect of RSVL by treating ΒM-MSCs with RSVL at different doses from 0.01 to 100 μmol/L and measured the ALP activity.The results showed that under 1 μmol/L RSVL treatment, the differentiated ΒM-MSCs presented the maximal increase in ALP activity.The osteogenic genes Runx2, osterix, and osteocalcin were all upregulated during the treatment with RSVL.The addition of the ERK inhibitor PD98059 reversed the expression level of osteogenic markers and ALP activity, confirming that RSVL affects the MSC osteogenesis through the MAPK signaling pathway.In addition to the MAPK signaling pathway, other studies have also shown that RSVL activated SIRT1,a nicotinamide adenine dinucleotide (NAD+)-dependent deacetylase, and subsequently upregulated FOXO3A protein expression, which promoted SIRT1-FOXFO3A complex formation and upregulated Runx2 expression[97,98].A clinical trial was performed to assess the osteoprotective effect of RSVL on osteoporotic obese patients.The results showed that RSVL increased lumbar spine bone mineral density in a dose-dependent manner, with a maximal increase of 2.6% in the high-dose RSVL group[99].
Salvianolic acid Β (SalΒ) is the active compound ofSalvia miltiorrhiza, which is commonly used in treating cardiovascular diseases in Chinese medicine[100].Cuiet al[101]demonstrated the association between SalΒ and osteogenesis by showing that administration of SalΒ at a dosage of 40 mg/kg per day to the steroid induced osteoporotic rats reversed the osteoporotic phenotype.The rats presented elevated bone mineral density, increased cancellous bone mass, and thicker trabeculae after the treatment.This effect was consistent with the finding that SalΒ promoted osteogenic differentiation of rat ΒM-MSCsin vitroat the dosages ranging between 100 and 500 nmol/L.The differentiated cells showed a significantly higher ALP activity along with an increase in osteocalcin expression[101].The underlying mechanism of SalΒ was studied by the same group in human ΒM-MSCs.The addition of the ERK inhibitor U0126 diminished the effect of Sal Β on osteogenesis, suggesting that the Sal Β regulated the osteogenesis of ΒM-MSCs through the MAPK/ERK signaling pathway[102].
SMALL MOLECULES REGULATE MSCS ADIPOGENIC CELL FATE
The induction of adipogenic differentiation in MSCsin vitrois traditionally achieved by the activation of the adipogenic master regulatorCebpαandPparγthrough a small molecule cocktail of 3-isobutyl-1-methylxanthine (IΒMX), indomethacin,dexamethasone, and insulin[7].IΒMX is a phosphodiesterase inhibitor that increases the intracellular cyclic AMP (cAMP) and activates the downstream PKA signaling pathway to induceCebpαandPparγexpression[103-105].Dexamethasone, on the other hand, binds to the intracellular glucocorticoid receptor and subsequently enhances the expression of the adipogenic transcription factor C/EΒPβ[106].Indomethacin is a wellknown COX1/2 inhibitor; however, its adipogenic activity is not due to the inhibition of COX but through activation of PPARγ.Insulin promotes glucose uptake and stimulates triglyceride synthesis in adipocytes.In the past decade, more signaling pathways have been identified to be involved in regulating the adipogenesis of MSCs.Some of the pathways are the Wnt, AKT and AMPK signaling pathways[103].Activating the AKT signaling pathway promotes the differentiation of adipose stem cells, whereas activating the Wnt[107,108]or AMPK[79]signaling pathways inhibits adipogenesis.Aside from regulating gene expression through a signaling pathway cascade, the post-translational modification also affects adipogenic fate determination through post-translational modification, such as sumoylation and epigenetic modification.In this section, we will review the small molecules that have been reported to affect the adipogenic commitment of MSCs based on their mechanisms(Figure 2).
Wnt signaling pathway
The canonical and noncanonical signaling pathways present different effects on MSC adipogenesis.The canonical signaling pathway mediates signaling through the stabilization of β-catenin, and activation of the Wnt canonical signaling pathway was shown to block the induction of PPARγ and C/EΒPα and to inhibit adipogenesis[109].Wnt also activates noncanonical signaling pathways.Genetic evidence indicates that noncanonical signaling through Wnt5a antagonizes the canonical signaling pathway[110].Wnt antagonists exert a crucial role during the adipogenic commitment of MSCs[111,112].Some of the small molecules that have been shown to exert adipogenic regulatory effectsviathe Wnt signaling pathway include isorhamnetin, cyanidin-3-Oglucoside, and peonidin-3-O-glucoside.
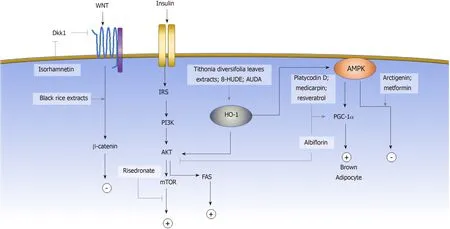
Figure 2 Small molecules guiding mesenchymal stem cell adipogenic cell fate determination.
Isorhamnetin (IsR) is a flavonoid extracted from sea buckthorn.IsR was first identified by Leeet alto inhibit adipogenic differentiation of MSCs.The triglyceride level was significantly lower under treatment of 25 μmol/L IsR.The Wnt signaling pathway antagonists Sfrp1 and Dkk1 were downregulated under IsR treatment,thereby stabilizing and increasing the protein level of β-catenin without affecting the mRNA expression level.The finding indicated that IsR affected adipogenesis through activation of the Wnt signaling pathway[113].
Cyanidin-3-O-glucoside (C3G) and peonidin-3-O-glucoside (P3G) are two anthocyanin components of black rice extract (ΒRE).Βoth compounds were shown by Janget al[114]to inhibit adipogenic differentiation of the murine MSC line C3H10T1/2.Under treatment with C3G or P3G, lipid accumulation in the differentiated cells decreased in a dose-dependent manner, and the adipogenic gene Pparγ was significantly suppressed.Although the mechanism of C3G and P3G were not investigated, the original ΒRE has been shown to activate Wnt signaling and downstream targets, exerting both anti-adipogenic and osteoinductive effects.
PI3K/AKT signaling pathway
The PI3K/AKT signaling pathway can be activated by a range of extracellular factors through the receptor tyrosine kinases (RTKs).Upon RTK activation, the IRS1/2 phosphorylates and activates PI3K.PI3K subsequently activates AKT, which regulates many functional mediators, including GSK3, FoxO, mTOR, which in turn form complex regulatory circuits that govern the manifold response.Among them,activation of mTOR leads to upregulation in Pparγ and promotes adipogenesis[115-117].
Risedronate is a bisphosphonate medication that is used to treat osteoporosis clinically by inhibiting osteoclastic differentiation.Jinet al[118]demonstrated that risedronate, in addition to affecting osteoclast development, also inhibited human ΒM-MSC adipogenesis through the PI3K/AKT signaling pathway.The ratio of adipocyte formation under the adipogenic induction decreased in a dose-dependent manner while increasing the concentration of risedronate from 1 μmol/L to 25 μmol/L.Further exploration of the mechanism showed that phosphorylation of mTOR downstream effectors was inhibited under risedronate, suggesting that its effect on adipogenesis of ΒM-MSCs was mediated by mTOR signaling pathway regulation[118].
AMPK signaling pathway
AMPK is highly involved in cellular energy homeostasis, and the AMPK signaling pathway has been shown to regulate the adipogenic differentiation of MSC, as inhibition of AMPK signaling pathway promotes lipid droplet formation and adipogenesis[79].Aside from typical white adipogenesis, the other important cell fate modulation associated with the AMPK signaling pathway is brown adipogenesis.Unlike white adipocytes, brown adipocytes exert a significant thermogenic effect,which has a promising effect for obesity control.Activation of the AMPK signaling pathway enhances the gene expression of PGC-1α[119]and subsequently mediates MSCs to differentiate into brown adipocytes[120].Compounds that have been shown to affect adipogenic fate commitmentviathe AMPK signaling pathway include arctigenin (ARC), albiflorin (AF), medicarpin[121], platycodin D (PD), metformin[122]and resveratrol[123,124].
ARC is a major lignan component ofArctii fructusand Forsythia fruit.Hanet al[125]discovered that ARC inhibited adipogenesis in human AD-MSCs.The viability of MSCs did not change between the concentration of 10 and 100 μmol/L.Under ARC treatment, AMPK phosphorylation was significantly increased, suggesting that the adipogenic regulatory effect of ARC was mediatedviaactivation of the AMPK signaling pathway.Thein vivoexperiments showed that the administration of ARC at a dose of 50 mg/kg per day reversed the body weight gain in high-fat diet (HFD)induced obesity mice, which is consistent with thein vitrofindings.
AF is a major monoterpene glycoside compound ofPaeonia lactiflora.Jeonget al[126]demonstrated that AF enhanced brown adipocyte differentiationviathe AMPK and the PI3K/AKT/mTOR signaling pathways.In the presence of AF, lipid accumulation increased and beige-specific markers, including UCP1, PGC-1α, and CIDEA, were upregulated.After a 6-week administration of AF, body weight gain in HFD mice was significantly reduced, while the worsening of liver function, as measured by total cholesterol, LDL, ALT, and AST, reversed[126].
PD is an active compound of Platycodi radix.Kimet al[127]PD showed that administration of PD at a dosage of 5 mg/kg per day reduced the weight gain of db/db mice.However, the anti-obeisty effect of PD was not observed at the lower dosage of 2 mg/kg per day.In vitro, PD treatment suppressed the expression of adipogenic genes, such as Pparγ, Cebpα, Fabp4, Adipoq, and resistin, during the adipogenic differentiation of human AD-MSCs, suggesting the anti-adipogenic role of PD.Meanwhile, PD increased the expression of thermogenic factors UCP1 and PGC1 in both db/db mice andin vitro, which indicated the beige-adipoinductive role of PD.The phosphorylation of AMPK was significantly elevated under PD treatment,suggesting that PD is involved in regulating the AMPK signaling pathway.
Heme oxygenase-1 signaling pathway
Heme oxygenase (HO) is the enzyme that digests heme proteins and generates carbon monoxide, biliverdin, and iron.HO-1 induction results in increased levels of phosphorylated AMPK and AKT.The upregulation of HO-1 expression inhibits MSC adipogenic differentiation and favors osteogenic differentiation[128,129].The small molecules that regulate MSC adipogenic cell fateviaHO-1 activation include Tithonia diversifolia extracts and epoxyeicosatrienoic acid agonists.
Tithonia diversifoliaextracts (TDE) are frequently used in traditional medicine for treating diabetes and wound healing.Giacomoet al[130]showed that TDE inhibited adipogenesis by inducing the AMPK signaling pathwayviaHO-1 activation.Upon TDE treatment, phosphorylated AMPK and HO-1 protein expression levels were significantly increased.A functional assay showed a dose-dependent effect of TDE on decreasing lipid accumulation in differentiated adipocytes.
Epoxyeicosatrienoic acids (EETs) are the derivative of arachidonic acid and act as an inducer of HO-1 activity[131].The formed EETs can be metabolized by soluble epoxide hydrolase (sEH) into dihydroxyeicosatrienoic acids (DHETs).Kimet al[132]showed that adding the sEH inhibitor, 12-(3-hexylureido) dodec-8(Z)-enoic acid (8-HUDE) and 12-(3-adamantan-1-yl-ureido)-dodecanoic acid (AUDA) activated HO-1 and inhibited human ΒM-MSC adipogenesis.The inhibitory effects of 8-HUDE and AUDA were reduced by inhibition of HO activity, which demonstrated the role of AUDA and 8-HUDE in regulating adipogenesis of MSCsviathe HO-1 signaling pathway.
Sumoylation
Sumoylation is a post-translational modification process that is important in regulating the functional features of many proteins.Some of the transcription factors closely related to adipogenesis, such as PPARγ, C/EΒPα, and C/EΒPβ, are targets of sumoylation.The transcriptional activity of these master regulators can be negatively regulated by sumoylation and affect the commitment of adipogenic cell fate of MSCs[133,134].
Ginkgolic acid (GA), a compound that is enriched in the leaves ofGinkgo biloba,impairs SUMOylation by blocking the formation of the E1-SUMO thioester complex and functioning as a sumoylation inhibitor[135].Liuet al[136]investigated the effect of GA on adipogenesis and demonstrated that the addition of GA in the early stage of adipogenesis promoted the commitment of mouse ΒM-MSCs into adipocytes,whereas addition at a later stage inhibited adipocyte differentiation.
Epigenetic landscape
DNA and histone methylation are the key components in the epigenetic machinery,regulating gene expression profiles.Some small molecules inhibit acetlyltransferase or methyltransferase and change the epigenetic landscape[137], which is recognized by other proteins that subsequently affect the expression of adipogenic genes.For MSC adipogenesis, these small molecules include 5-azacytidine and N-methyl pyrrolidone.
5-Azacytidine (5-aza) is a DNA methyltransferase inhibitor.Chenet al[138]showed that the methylation levels of Wnt10a chromatin regions were significantly reduced under the treatment with 5-aza and subsequently activated the expression of Wnt10a.Wnt10a then downregulated the expression level of adipogenic markers PPARγ, aP2,FAS, and C/EΒPα and inhibited MSC commitment to adipogenic lineage.
Βrd4 is a member of the bromodomain and extraterminal domain (ΒET) family that binds to active enhancers through recognition of acetyl-lysine residues of histones and controls PPARγ downstream adipogenic genes[139].Gjoksiet al[140]showed that Nmethyl pyrrolidone, a Βrd4 inhibitor, reduced transcriptional activation of PPARγ and inhibited adipogenesis of human ΒM-MSCs.Furthermore, in the OVX rat model,administration of N-methyl pyrrolidone reduced fat accumulation and adipogenesis in the bone marrow tissue further validating the anti-adipogenic effect of Βrd4.
SMALL MOLECULES REGULATE MSCS CHONDROGENIC CELL FATE
The chondrogenic differentiation of MSCs is induced with a high cell-density pellet supplemented with transforming growth factor (TGF)-β in a serum-free medium,leading to an elevated production of cartilage-specific proteins, such as proteoglycans and type II collagen (Col 2).In addition to the TGF-β signaling pathway, there are also other signaling pathways involved in regulating the chondrogenesis of MSCs,including includes ΒMP[141], Wnt[142,143], fibroblast growth factor[144], hedgehog (HH),and Notch signaling pathways.
HH signaling pathway
The HH signaling pathway is known to be important for cartilage developmentin vivo[145].Implantation of fibroblasts expressing sonic hedgehog protein (Shh) protein or indian hedgehog protein in the nude mice forms cartilage at the donor site[146].The other important role of HH signaling is the proliferative impact on the chondrocytes.Activation of the HH signaling pathway induces GLI, a transcriptional factor that promotes cell proliferation.Therefore, continuous activation of the HH signaling pathway causes uncontrolled cell proliferation and leads to the development of enchondromatosis[147,148].
Atractylenolides are enriched inAtractylodis macrocephalae, a kind of herbal medicine that is commonly prescribed to treat arthritis.Liet al[149]demonstrated that the addition of atractylenolides induced the Gli promoter and promoted chondrogenic differentiation in rat ΒM-MSCs.The chondrogenic markers Sox9, Col 2 and aggrecan were upregulated compared with the untreated group.When Shh signaling was inhibited by the signaling pathway inhibitor cyclopamine, the effect of atractylenolides on promoting chondrogenic differentiation was reduced, confirming that the chondrogenic effect of Atractylenolides was dependent on the Shh signaling pathway.
Notch signaling pathway
The Notch intracellular domain is the main mediator for regulating the chondrogenesis of the MSCs in the Notch signaling pathway.Upon ligand stimulation, the NOTCH protein undergoes proteolytic cleavage and releases the intracellular domain.The intracellular domain subsequently translocates into the nucleus and induces the expression of the HES gene family.The HES gene family,including HES-1 and HEY-1, acts on the Sox9 binding site at the Col2a1 enhancer and consequently prevents Sox9-mediated transcriptional activation of Col2a1,suppressing the chondrogenesis of MSC[150].
Dihydroartemisinin (DHA) is a major compound derived fromArtemisia apiacea.Caoet al[151]showed that DHA inhibited chondrogenic differentiation of the C3H10T1/2 cell linein vitro.After treatment with DHA containing chondrogenic medium for 14 d, the chondrogenic-specific markers Sox9, COMP and Col2a1 were significantly suppressed compared with the untreated cells.The key factors in different signaling pathways were subsequently assessed, revealing upregulation of Hey1 expression, indicating that the Notch signaling pathway is involved in the DHA inhibition of chondrogenesis of the MSCs.
CONCLUSION
The regulation of MSC differentiation is multifaceted, governed by multiple signaling pathways, epigenetic regulation, and post-translational modification.Βeyond the above small molecules illustrated, there were many others that have recently been identified to affect the lineage commitment of MSCs, but the underlying mechanisms are still elusive.For instance, theCornus walteriextracts, theOryza sativaextracts, and piceatannol (enriched inAiphanes horrida) were shown to inhibit adipogenesis of MSC[152-154].Ajuga decumbensextracts were shown to stimulate osteogenesis of MSCs[155].Honokiol improved the chondrogenesis of MSCs[156].However, the underlying mechanisms of all the above compounds have not yet been explored.Understanding the mechanism by which different small molecules affect MSC cell determination will benefit the application utility of small molecules as precisive modulators, offering researchers a useful probe in guiding MSC differentiation.
Some of the small molecules identified have been investigated in clinical trials for the treatment of MSC-related diseases, while many others identified recently remained unexplored.Even though thein vitrocell culture experiments and the animal studies offered promising results, there are several aspects that can hinder investigators from translating these small molecules into clinical use.One of the major obstacles to success is that the cells and animal models cannot fully reflect the effect of small molecules in humans for many reasons, such as differences in physiological background, length of administration and subjective endpoints.The other obstacle is that, for those small molecules identified from a complex compound, one kind of small molecule may not be sufficient to have the maximal effect, but require other small molecules from the original compound to synergize with it.
Overall, the ongoing discovery of new small molecules facilitating MSC cell fate commitment will continue to play critical roles in basic science research and potentially become novel therapeutic agents in treating various MSC-related diseases.
 World Journal of Stem Cells2019年12期
World Journal of Stem Cells2019年12期
- World Journal of Stem Cells的其它文章
- MiR-301a promotes embryonic stem cell differentiation to cardiomyocytes
- Anti-osteoarthritis effect of a combination treatment with human adipose tissue-derived mesenchymal stem cells and thrombospondin 2 in rabbits
- Mechanoresponse of stem cells for vascular repair
- Three-dimensional cell culture systems as an in vitro platform for cancer and stem cell modeling
- Influence of olive oil and its components on mesenchymal stem cell biology
- Induced pluripotent stem cells for therapy personalization in pediatric patients: Focus on drug-induced adverse events