
入侵植物瘤突苍耳(Xanthium strumarium)种子萌发期表型代间传递可塑性研究
2019-12-25 07:43:50成清龙
沈阳农业大学学报 2019年6期
曲 波,陈 鲜,张 婷,成清龙
(沈阳农业大学 生物科学技术学院/辽宁省全球变化与生物入侵重点实验室,沈阳110161)
生物入侵这一概念出现较晚,但人类对于生物入侵的研究由来已久。早在19世纪,达尔文就在其著作《物种起源》中多次提到生物的转移和传入现象,但他未强调其危害(如对生物多样性的危害),当时并未引起人们的重视。随着全球经济一体化的发展以及贸易往来和人类活动日渐频繁,外来生物入侵也随之加剧[1-3],并严重地威胁了入侵地生态系统的生物多样性,改变其生态系统的结构和功能[4],这才逐渐引起社会各界人士以及各国政府的重视。
有关生物入侵机制有多种科学假说,如表型可塑性、天敌逃逸、新武器、竞争力增强进化、生长繁殖和防御分配、化感作用、生物阻抗、生态位空缺、资源-天敌相互作用、氮分配等。这些假说从生物的入侵性和环境的可入侵性等多方面解释外来生物的入侵机制。其中BAKER提出的表型可塑性已有半个世纪之久,目前众多研究表明,在动植物界普遍存在表型可塑性,其与生物入侵的关系越来越受到人们的关注[5-13]。
表型可塑性是指生物的一种基因型对不同环境产生不同表型性状的应答[14-17],具有遗传基础,也能发生进化。在自然界中,所有的生物都可能遭遇异质生境,其中一些生物可以主动迁移至较好的环境中,而不能自由移动的生物就需要依赖除运动之外的其他生存策略。植物不仅几乎无法自主选择生长环境,而且多数物种的种子传播距离有限,大多数的种子都落在亲代植株周围。因此,植物子代的生长环境通常与亲代相似,尤其当两代的生境相同且范围大于种子传播的距离时,可能存在亲本生境诱导产生的适应性线索,这些线索可增强子代对该生境的适应能力[18],即生长在与母体相同环境中的子代比生长在其他环境中的子代更适应环境,这意味着植物亲代会“暗示”它们的子代如何适应当地的环境条件,即所谓的“代间传递可塑性”(transgenerational plasticity)。表型可塑性是植物对当前环境条件产生的适应性,而代间传递可塑性是通过个体对环境线索的可塑性应答影响其后代的表型和适应[18-23]。代间传递可塑性也能解释一些非本地种在多变环境中的种群变化[24-25]。
然而,由于缺乏对其适应性意义的可靠检验,无论这种效应是被动反应还是代表一种特殊类型的适应性可塑性,仍然存在争议。本研究以自花授粉的菊科苍耳属植物瘤突苍耳(Xanthium strumarium)[26]全同胞家系为试验材料,设置不同养分条件下的种子萌发试验,探讨植物种子萌发期表型代间传递的可塑性是否与环境因子有关,为丰富植物入侵机制提供参考。
1 材料与方法
1.1 材料
瘤突苍耳果苞(内含2粒种子)于2014年采自辽宁省灯塔市(县级)沈旦堡镇周官村浑河河滩沙地,风干后室温保存。2015年将一部分种子种植于浑河河滩耕地,其土壤类型为河淤土,代表高养分环境,秋季收获种子(记为H);另一部分种植于浑河河滩沙地,代表低养分环境,秋季收获种子(记为L),采集地土壤养分含量如表1。所收获种子置于-20℃冷冻保存。
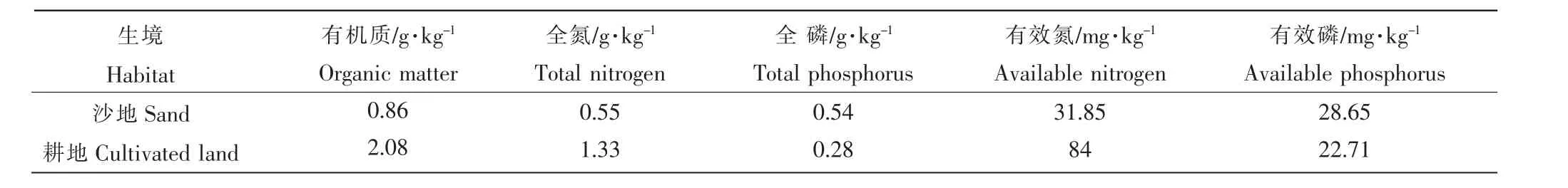
表1 辽宁省灯塔市(县级)沈旦堡镇周官村土壤养分状况Table 1 Soil nutrition in Shendanpu town of Dengta city in Liaoning province
1.2 方法
2017年7月进行种子在不同养分条件下的萌发试验。配制高低两种养分的营养液,高养分为正常浓度的Hoagland's营养液,低养分为氮素浓度减半的Hoagland's营养液,为区分亲代养分处理,分别记为小写字母h和l。分别取亲代低养分(L)、亲代高养分(H)和原种(P)种子5,7和10个家系,每个家系10粒种子。为排除种子在果苞中不同位置对试验结果的影响,所用种子均为去除果苞后的下位种子。用0.5%KMnO4溶液消毒10分钟,然后用蒸馏水冲洗干净,其中5粒置于含高养分营养液的种子萌发袋中(h),另5粒置于低养分营养液的种子萌发袋中(l),种子萌发袋规格为12.5cm×18cm,营养液添加量以保证种子萌发袋中的纸芯湿润,视蒸发量每天酌情补充相应的营养液。从播种第2天开始,每天记录种子萌发数量,第5天用直尺测量每株幼苗主根长度、侧根长度、幼苗高度,用游标卡尺测量每个植株的子叶长度、子叶宽度和子叶厚度。据测量结果计算种子萌发率,发芽势和发芽指数。

式中:Gt为第t天的发芽数;Dt为相应发芽天数
1.3 数据使用SPSS13.0进行分析,使用Excel 2010作图。
2 结果与分析
2.1 不同养分条件的瘤突苍耳亲代种子在不同养分条件下萌发期和幼苗期表型特征
通过数据分析发现,来自高养分亲代的子代除了在低养分条件下的发芽势、子叶厚度、幼苗高度、主根长度低于来自低养分亲代的子代,其他条件下各指标均高于来自低养分亲代的子代。
无论亲代在高养分条件下生长还是在低养分条件下生长,其种子都在高养分条件下有较高的萌发率(图1A),其中当亲代在高养分条件下生长,子代也在高养分条件下生长时发芽率最高,高达94%,当亲代在低养分条件下生长,子代也在低养分条件下生长时,其发芽率最低,为76%。当亲代在高养分条件下生长时,其子代在高养分条件下有较高的发芽势(图1B),为68%,而在低养分条件下的发芽势为54%。当亲代在低养分条件下生长时,其子代在低养分条件下有较高的发芽势,为56%,而在高养分条件下的发芽势仅为48%。二者的发芽指数没有显著区别(图1C)。
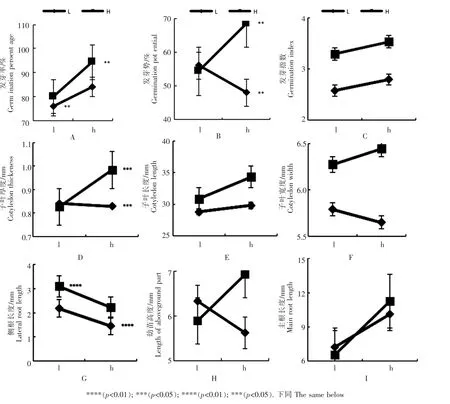
图1 子代种子在不同养分条件下萌发特性与幼苗生长状况Figure 1 Germination characteristics and seedling growth of offspring seeds under different nutrient conditions
当亲代在高养分条件下生长时,其子代在高养分条件下有较大的子叶厚度(图1D),为0.98mm(p<0.05),在低养分条件下的子叶厚度为0.82mm(p<0.05),当亲代在低养分条件下生长时,其子代在低养分条件下有较大的子叶厚度,为0.84mm(p<0.05),在高养分条件下的子叶厚度为0.82mm(p<0.05)。而在子叶长度与子叶宽度方面没有明显区别(图1E和图1F)。
当亲代在高养分条件下生长时,其子代在低养分条件下侧根长度较大(图1G),为3.1cm(p<0.01),在高养分条件下为2.2cm(p<0.01),当亲代在低养分条件下生长时,其子代在低养分条件下侧根长度较大,为2.2cm(p<0.01),在高养分条件下为1.4cm(p<0.01)。而在幼苗高度与侧根长度方面没在明显区别(图1H和图1I)。
2.2 瘤突苍耳不同养分条件下亲代产生的子代种子与其亲代相比在不同养分条件萌发期和幼苗期表型变化率
通过试验发现,不同养分瘤突苍耳亲代产生的子代种子与其亲代相比在不同养分条件萌发期和幼苗期表型存在一定变化。从发芽率来看,低养分条件下生长的亲代,其子代在高养分条件下发芽率比原种高2.43%(p<0.01),在低养分条件下发芽率比原种低17.39%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下发芽率比原种高14.98%(p<0.01),在低养分条件下发芽率比原种低13.04%(p<0.01)(图2A)。从发芽势来看,低养分条件下生长的亲代,其子代在高养分条件下发芽势比原种低25%(p<0.01),在低养分条件下发芽势比原种低26.31%(p<0.01);高养分条件下生长的亲代,其子代高养分条件下发芽势比原种高7.14%(p<0.01),在低养分条件下发芽势比原种低28.57%(p<0.01)(图2B)。对于发芽指数,低养分条件下生长的亲代,其子代在高养分条件下发芽指数比原种低15.02%(p<0.01),在低养分条件下发芽指数比原种低21.7%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下发芽指数比原种高7.47%(p<0.01),在低养分条件下发芽指数比原种高0.12%(p<0.01)(图2C)。对于子叶长度,低养分条件下生长的亲代,其子代在高养分条件下子叶长度比原种低16.55%(p<0.01),在低养分条件下子叶长度比原种低21%;高养分条件下生长的亲代,其子代在高养分条件下的子叶长度比原种低4.02%(p<0.01),在低养分条件下的子叶长度比原种低15.39%(p<0.01)(图2D)。对于子叶宽度,低养分条件下生长的亲代,其子代在高养分条件下子叶宽度比原种低23.44%(p<0.01),在低养分条件下子叶宽度比原种低27.58%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下的子叶宽度比原种低12.71%(p<0.01),在低养分条件下的子叶宽度比原种低21.52%(p<0.01)(图2E)。对于子叶厚度,低养分条件下生长的亲代,其子代在低养分条件下的子叶厚度比原种低12.26%(p<0.01),在高养分条件下的子叶厚度比原种低16%(p<0.01);高养分条件下生长的亲代,其子代在高养分的子叶厚度比原种高0.76%(p<0.01),在低养分条件下的子叶厚度比原种低16.7%(p<0.01)(图2F)。
对于幼苗高度,低养分条件下生长的亲代,其子代在高养分条件下生长时幼苗高度比原种高5.65%(p<0.01),在低养分条件下生长时幼苗高度比原种高26%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下生长时幼苗高度比原种高30.13%(p<0.01),子代在低养分条件下生长时幼苗高度比原种高17.24%(p<0.01)(图2G)。对于主根长度,低养分条件下生长的亲代,其子代在高养分条件下生长时主根长度比原种高55.38%,低养分条件下生长时主根长度比原种低32.54%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下主根长度比原种高73.08%(p<0.01),低养分条件下主根长度比原种低38.79%(p<0.01)(图2H)。对于侧根长度,低养分条件下生长的亲代,其子代在高养分条件下侧根长度比原种低39.14%(p<0.01),在低养分条件下侧根长度比原种低32.37%(p<0.01);高养分条件下生长的亲代,其子代在高养分条件下侧根长度比原种低6.47%(p<0.01),在低养分条件下侧根长度比原种低 3.66%(p<0.01)(图 2I)。
可见,当子代在高养分环境条件下生长时,亲代也在高养分条件下生长的子代的各种指标均大于亲代在低养分条件下生长的子代。当子代在低养分条件下生长时,发芽势、子叶厚度和主根长度三项指标在亲代也在低养分环境条件下生长时较大,其余各项指标小于亲代在高养分环境条件下生长时的各项指标。不同养分瘤突苍耳亲代产生的子代种子与其原种相比在不同养分条件萌发期和幼苗期表型变化均存在极显著差异(p<0.01)
2.3 瘤突苍耳相同养分条件下亲代的子代在高养分与低养分条件下9种表型指标值的差异
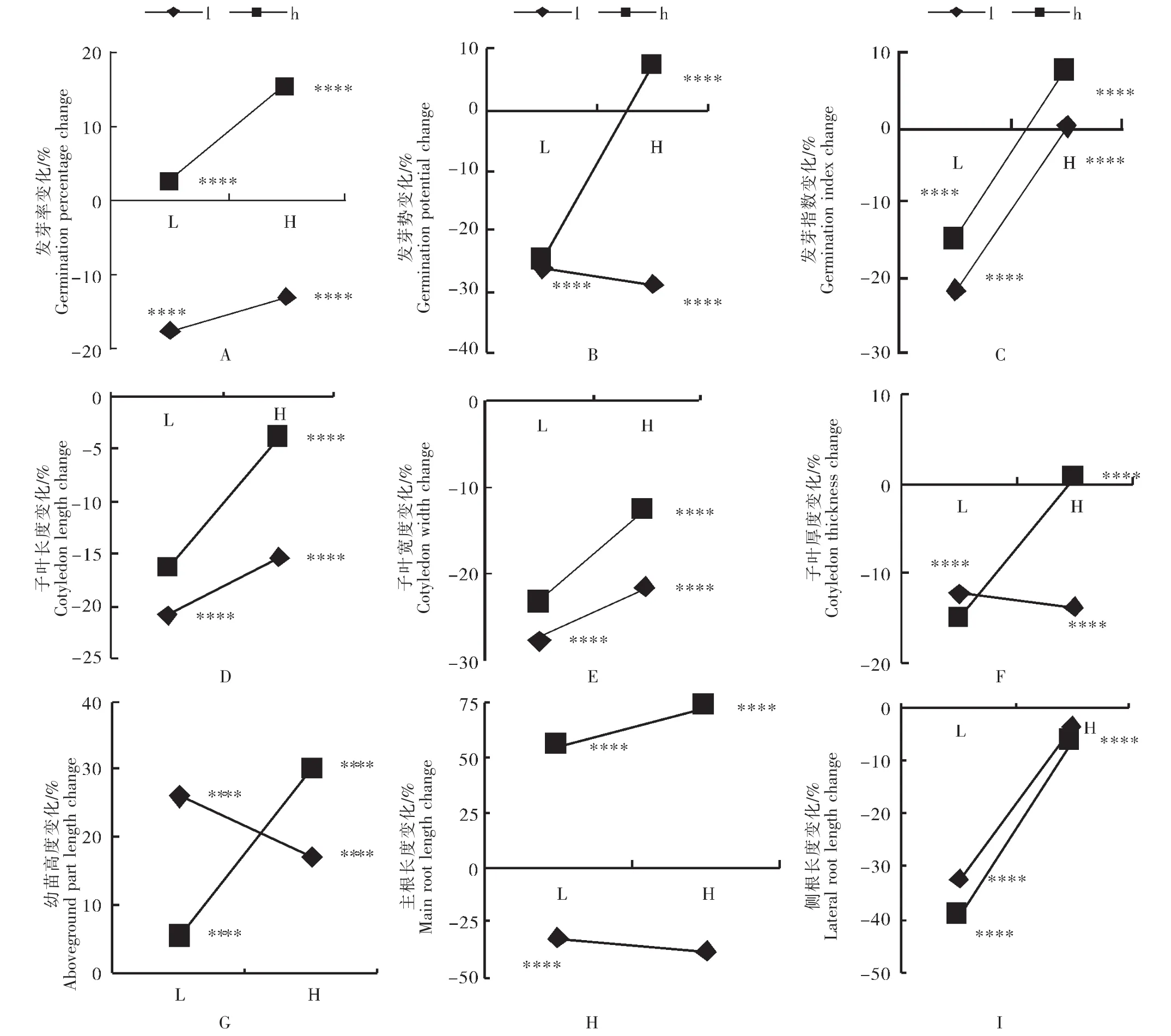
图2 不同处理亲代的子代在不同养分条件下与其亲代相比萌发特征与幼苗生长状况的变化Figure 2 Changes of germination characteristics and seedling growth of offspring of parents with different treatments under different nutrient conditions compared with their mothers
对于发芽率,低养分条件下生长的亲代,其子代在高养分条件下的发芽率比低养分条件下高10.52%,高养分条件下生长的亲代,其子代在高养分条件下生长的发芽率比低养分条件下高19.86%(图3A)。对于发芽势,低养分条件下生长的亲代,其子代在高养分条件下的发芽势比低养分条件下低14.29%,高养分条件下生长的亲代,其子代在高养分条件下生长的发芽势比低养分条件下高26.32%(图3B)。对于发芽指数,低养分条件下生长的亲代,其子代在高养分条件下的发芽指数比低养分条件下高8.41%,高养分条件下生长的亲代,其子代在高养分条件下的发芽指数比低养分条件下高7.24%(图3C)。对于子叶长度,低养分条件下生长的亲代,其子代在高养分条件下的子叶长度比低养分条件下高3.46%,高养分条件下生长的亲代,其子代在高养分条件下生长的子叶长度比低养分条件下高11.12%(图3D)。对于子叶宽度,低养分条件下生长的亲代,其子代在高养分条件下的子叶宽度比低养分条件下低2.40%,高养分条件下生长的亲代,其子代在高养分条件下生长的子叶宽度比低养分条件下高2.70%(图3E)。对于子叶厚度,低养分条件下生长的亲代,其子代在高养分条件下的子叶厚度比低养分条件下低1.42%(p<0.15),高养分条件下生长的亲代,其子代在高养分条件下生长的子叶厚度比低养分条件下高18.94%(p<0.15)(图3F)。对于幼苗高度,低养分条件下生长的亲代,其子代在高养分条件下的幼苗高度比低养分条件下低11.18%,高养分条件下生长的亲代,其子代在高养分条件下生长的幼苗高度比低养分条件下高28.21%(图3G)。对于主根长度,低养分条件下生长的亲代,其子代在高养分条件下的主根长度比低养分条件下高39.63%,高养分条件下生长的亲代,其子代在高养分条件下生长的主根长度比低养分条件下高71.41%(图3H)。对于侧根长度,低养分条件下生长的亲代,其子代在高养分条件下的侧根长度比低养分条件下低33.45%,高养分条件下生长的亲代,其子代在高养分条件下生长的侧根长度比低养分条件下低28.21%(图 3I)。
可见,当亲代在高养分条件下生长时,其子代除侧根长度外,其他指标均为高养分条件下表现更好,表明子代在高养分条件下生长更好;当亲代在低养分条件下生长,子代在低养分条件下的发芽势、子叶宽度、子叶厚度、幼苗高度四个指标较高养分条件下数值大,表明子代在低养分环境条件下生长更好。但统计结果表明,当亲代在同一养分条件下生长,子代在高养分条件下与低养分条件下9种表型指标值的差异不显著,仅在子叶厚度上存在一定差异(p<0.15)。
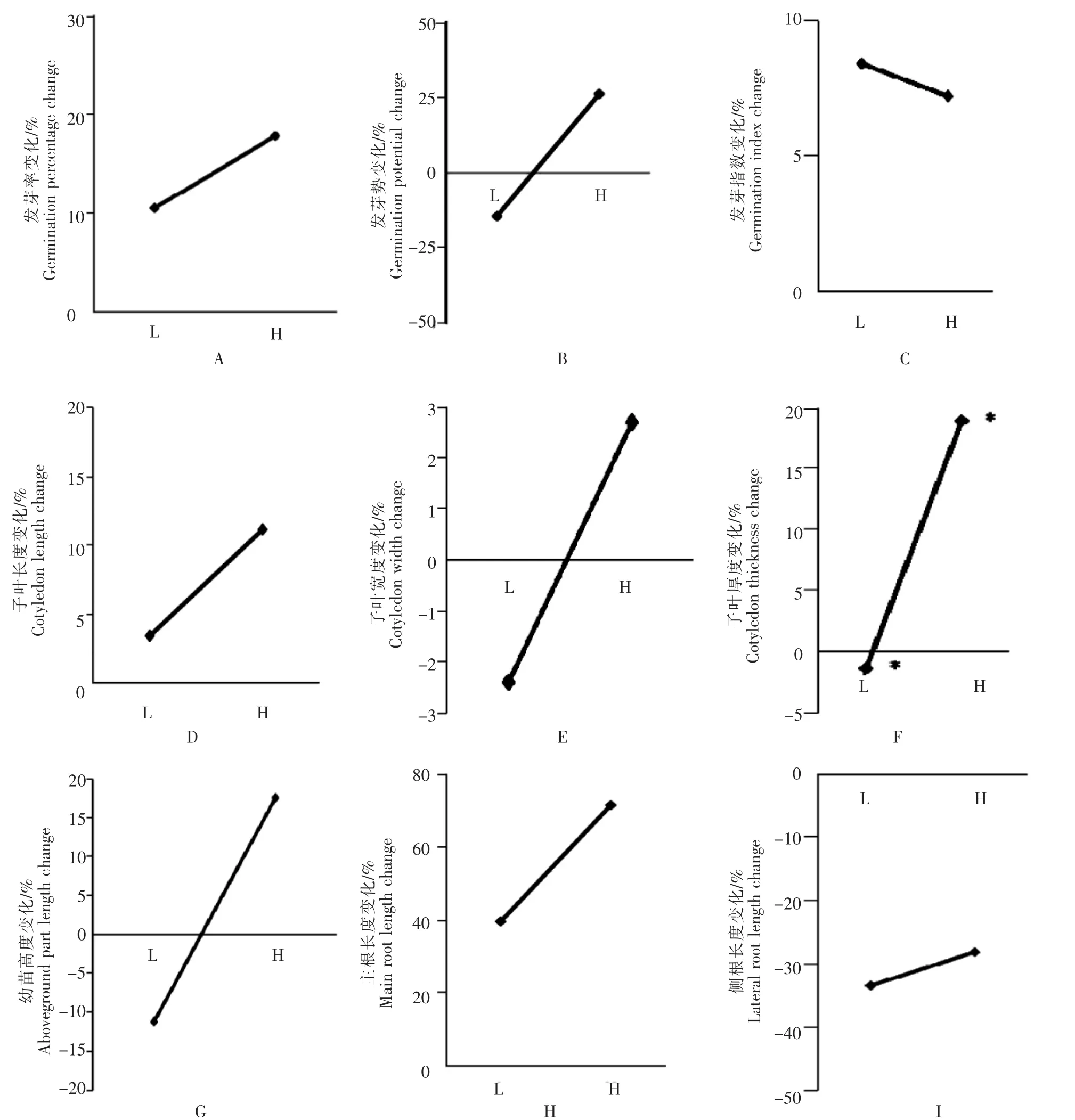
图3 相同处理亲代的子代在不同养分条件下种子萌发特征与幼苗生长状况变化Figure 3 Seed germination characteristics and seedling growth of offspring of parents with the same treatment under different nutrient conditions
3 讨论与结论
本试验通过研究我国入侵植物瘤突苍耳种子萌发过程发现,低养分条件下生长的亲代,其子代发芽率、发芽指数、子叶长度、子叶宽度、子叶厚度和侧根长度这些性状为高养分条件表现更好,而发芽势、主根长度和幼苗高度为低养分条件下表现更好。AGRAWAL等[23]提出母体对土壤条件的反应能以更高的光合效率的形式赋予幼苗更大的抗逆境能力,与本研究结果相近。
亲代在高养分条件下生长时,其子代在不同环境中的表型差异较大。对于发芽势、子叶宽度、子叶厚度和幼苗高度这4个表型,生长在低养分条件下子代表型变化高于生长在高养分时表型变化,这说明,当亲代在低养分条件下生长时,其子代在低养分环境条件下的可塑性变化低于高养分环境条件下的可塑性变化,验证了表型可塑性可以通过某种线索在代间传递,且传递的结果有利于子代适应环境。但统计分析表明,仅子叶厚度可塑性变化在0.05水平表现出显著性差异。SANDNER等[25]也认为母性环境可能影响后代的性状和表现,但这种跨代可塑性不一定具有适应性。
低养分条件下生长的亲代其子代生长在不同养分条件下,幼苗期侧根长度可塑性变化大于来自高养分高养分条件下生长的亲代,高养分条件下生长的亲代其子代在低养分条件下可塑性变化最大,说明高养分条件下生长的亲代,其子代的可塑性最强,而低养分条件下的亲代其子代在低养分条件下可塑性性变化次之,则说明低养分条件下生长的亲代,其子代依然在低养分条件下生长时,可塑性性变化低于高养分条件下亲代的子代,表现出一定的适应性。但是来自低养分条件下生长的亲代其子代在高养分条件下的表型与高养分条件下生长的亲代在高养分条件下的表型相比前者更能够体现可塑性在代间传递的效应。当子代在高养分条件下生长时,可能不再需要过多的侧根吸收营养,但是由于受亲代传递的某种信号的影响,子代依然生长了过多的侧根,这可能是生长的冗余部分。
高养分条件下生长的亲代其子代除侧根长度之外所有的性状均在高养分条件下表现更好,而低养分条件下生长的亲代其子代发芽率、发芽指数、子叶长度、主根长度4个性状在低养分环境条件下表现更好,这说明当子代与亲代生长环境条件相同时,亲代可以预测子代即将遇到的环境条件,并预制出适应性策略传递给子代,而发芽势、子叶宽度、子叶厚度、幼苗高度、侧根长度依然为高养分环境条件下表现更好,这说明并非所有的亲代表型都可以通过种子传递给子代。高养分亲代的子代在高低养分条件下的表型差异大于低养分亲代的子代在高低养分条件下的表性差异,这表明在高养分条件下生长的亲代,其子代对环境养分变化的响应更为明显。
当子代在高养分环境条件下生长时,亲代也在高养分条件下生长的子代的各种指标均大于亲代在低养分条件下生长的子代。当子代在低养分条件下生长时,发芽势、子叶厚度、幼苗高度和主根长度4项指标为亲代也在低养分环境条件下生长时较大,其余各项指标小于亲代在高养分环境条件下生长时的各项指标。本研究结果与GALLOWAY[27]对Campanulastrum americanum的研究结果相同,当子代与亲代环境条件相同时,子代对环境的适应性更高。本研究发现在资源匮乏时,子代的表型代间可塑性较低,与ANNAMÁRIA等[28]研究结论相同。
综上,植物的某些表型可在代间传递,亲代能把对环境的一部分记忆通过种子传递给子代,即子代能“记住”亲代的环境特征,对不良环境(资源匮乏)的记忆可能更深刻,因此低养分条件下生长的亲代其子代在低养分条件时可塑性变化最低。瘤突苍耳在种子萌发过程中,低养分条件下生长的亲代其子代的可塑性变化低于高养分条件下生长的亲代的可塑性变化,这可能瘤突苍耳成功入侵各种生境的原因之一,需要进一步深入研究,特别是与本地近缘种进行同质园试验进行比较。
猜你喜欢
四川蚕业(2022年2期)2022-11-19 02:10:24
四川蚕业(2022年2期)2022-11-19 02:09:52
昆明医科大学学报(2022年1期)2022-02-28 07:43:34
今古文创(2021年46期)2021-12-21 07:22:06
昆明医科大学学报(2021年8期)2021-08-13 08:59:20
昆明医科大学学报(2021年6期)2021-07-31 07:40:08
四川蚕业(2020年4期)2020-02-10 06:14:46
新传奇(2018年17期)2018-05-14 23:33:16
西南医科大学学报(2015年1期)2015-08-22 13:01:48
教育教学论坛(2014年43期)2014-10-22 15:37:10
