
钒在不同品种玉米幼苗中的亚细胞分布和动态变化
2019-12-25 07:13侯明杨心瀚霍岩罗萍
生态环境学报 2019年11期
侯明 ,杨心瀚 ,霍岩 ,罗萍
1. 桂林理工大学化学与生物工程学院,广西 桂林 541004;2. 广西高校食品安全与检测重点实验室,广西 桂林 541004
钒(V)是人和动物必需的微量元素,但超过一定限度有害人体健康。V 不是植物必须的微量元素,过量V 会影响大豆和水稻的正常生长(邹宝方等,1992;侯明等,2014)。V 显著影响鹰嘴豆植物中蛋白质含量,使植物生物量和根长等明显减少(Imtiaz et al.,2015)。有研究表明,V 对植物更具流动性和毒性(Panichev et al.,2006;Xiao et al.,2015;Larsson et al.,2013),因此,了解V 的生物学作用对评价V 对环境和生物系统的潜在风险很重要。近年来,V污染对土壤和生物的影响已引起人们关注,Saco et al.(2013)研究了V 胁迫下菜豆(Phaseolus vulgarisL. cv)根和叶的形貌和超微结构,表明当V 处理浓度增加,菜豆根和叶形貌发生变化,叶片中叶绿体数量减少,细胞类囊体解体,叶V 含量总是远低于根V 含量。Tian et al.(2014)研究发现,土壤生物活性会提高大白菜对V 的提取性,大白菜主要在根部积累V,V 从根部到地面部分的转运率较低。Teng et al.(2013)研究表明,土壤中加入V 质量浓度为0—400 mg·kg-1,V 主要富集在苜蓿根部,在地上部分略有积累。Yang et al.(2017)研究了土壤中V 对大豆不同生长阶段的毒性,高浓度V(Ⅴ)推迟了大豆萌发和生长,细胞壁V 浓度与根部总V 浓度比例随土壤中V(Ⅴ)浓度增加而增加。Imtiaz et al.(2018)探明,V 诱导使鹰嘴豆生物量、根系和幼苗长度显著降低,根系积累的V 大于幼苗;随着V 浓度增加,酶活性(SOD、CAT 和POD)和离子泄漏呈线性增加,蛋白质含量和耐受性指数显著下降,细胞活力受到严重损害。这些研究表明,低浓度V 处理促进植物生长,高浓度V 对植物产生毒性。然而,迄今为止,植物对V 的耐受机制仍然有限,对V 污染地区环境进行准确的风险评估仍有困难。由于采矿和冶炼活动增加,在中国西南地区有26.49%土壤被V 污染(Yang et al.,2017),因此,有必要进一步探明土壤-植物系统中V 的生物利用率以及V 的植物毒性和耐性。
研究植物在重金属胁迫下亚细胞组分的分布特征可以阐明植物的耐性机理。近年来,人们研究了玉米和水稻植株中Cd(Zhang et al.,2019;He et al.,2008;唐杰等,2016)和Y(Zheng et al.,2018),小麦中Pb 和Cu(杨素勤等,2015;苏忠亮等,2018),以及大豆幼苗中Mo(Xu et al.,2018)的亚细胞分布。这些研究结果表明,金属在植物细胞不同部位的分布大多数遵循:细胞壁>可溶性分数>细胞器的顺序,在细胞壁和可溶性组分的所占比例之和可达85%以上,说明植物为了抑制重金属对其的伤害,总会将金属分布在对植物生长相对不太重要的细胞器官。但金属在不同植物或同一物种不同品种中的细胞分布有显著差异(Liu et al.,2014;Xin et al.,2017)。因此,研究V 在植物体内的亚细胞分布,对阐明V 对植物的毒理和植物耐受性有重要意义。根据健康风险评估,广西也被标记为V 的优先控制区域(Yang et al.,2017),大量有色金属的开采、冶炼和加工,使土壤受到不同程度V 污染,将会影响到玉米作物的生长发育,可能导致玉米产量和质量降低,进而影响到人体健康。为了进一步阐明V在玉米作物的富集特性和毒性机理,笔者采用水培试验,研究了V 污染下不同品种甜玉米幼苗中V 的亚细胞分布及动态变化,旨在为探明不同品种玉米体内V 的富集规律和植物耐受机制提供科学依据。
1 材料与方法
1.1 试验材料
供试玉米(Zea maysL.)种子选取广西具有代表性且广泛种植的益甜608(广西浩凯生物科技有限公司)和美甜糯一号(大民种业股份有限公司)。
1.2 试验设计和处理
挑选籽粒饱满的玉米种子,置于质量分数为0.3% H2O2中消毒30 min,用纯水洗净后温水浸泡1h,于30 ℃烘箱中恒温催芽。培育玉米所用的容器为30 cm×24 cm×10 cm 的不透明塑料方盆,盆上盖有33 cm×26.5 cm 带孔(48 个直径2.3 cm 的孔洞)PVC 隔板。将已萌发小芽的玉米种子播种于隔板每个圆孔上的定植篮中,每盆定株48 株,24 h 供氧进行水培培养。待幼苗长至2 片叶时,用1/2 Hoagland 营养液培养,待玉米幼苗长至“两叶一心”时,用Hoagland 全营养液培养,待玉米幼苗长到第3 片叶时,挑选长势一致的幼苗用含V 的营养液培养。每个品种均设置4 个处理,V 胁迫质量浓度分别为0、5、15、30 mg·L-1(V 以NH4VO3形式添加),每种处理重复5 盆,两个品种共计40 盆,营养液每4 天更换一次(调节溶液pH 为5.5—6.5)。培养4、8、12 d 后分别采集每个处理80 株,10 株为一个试样,分根和茎叶用清水冲洗(根系于0.02 mol·L-1EDTA 溶液中浸泡10 min,以除去表面吸附的金属离子),再用纯水洗净。用滤纸吸干表面水分。将根系和茎叶分别剪碎,准确称取根系0.50 g和茎叶2.00 g,置于-20 ℃冰箱中备用。
1.3 试验方法
1.3.1 玉米组织的亚细胞组分分离方法
按杨居荣等(1993)方法改进。将适量冷藏的鲜样与预冷的亚细胞提取液(0.05 mol·L-1Tris-HCl,pH 7.5,0.25 mol·L-1蔗糖和0.001 mol·L-1二硫苏糖醇)以1:10(m/V)的比例混匀,在冰浴上迅速研磨成匀浆。使用差速离心技术分离植物组织。将匀浆液在冷冻离心机中以1 500 r·min-1离心20 min,沉淀部分为细胞壁(F1);上清液继续以2 500 r·min-1离心10min,沉淀部分为细胞核(F2),上清液再以10 000 r·min-1离心30 min,沉淀为线粒体和叶绿体(F3),上清液为可溶性溶液(F4),所有步骤在4 ℃下进行。
1.3.2 样品中V 含量的测定
将一定量植物鲜样磨成匀浆后,置于150 mL锥形瓶中(测定V 总量),同上述分离V 的亚细胞各组分一起,加入5:2(V/V)的HNO3和H2O2,置于数显控温电热板上充分消化至澄清,纯水定容后,利用 ICP-MS(1288090 iCAP Q Operating Manual,Rev.B)测定试样中V 含量。采用国家标准物质GBW 10015(GSB-6 菠菜)监控样品中V含量的测定结果。
1.4 数据处理
转运系数(translocation factor,TF)=茎叶V 质量分数(μg·g-1)/根系V 质量分数(μg·g-1)(Tiwari et al.,2011)。采用Microsoft Excel 2010 对数据进行统计分析,运用Duncan 多重比较法进行显著性差异分析(P<0.05)。结果以平均值±标准差(Mean±SD,n=3)表示。
2 结果与分析
2.1 不同品种玉米幼苗中V 的积累分布和转运
由表1 可知,在不同时间不同的V 浓度处理下,两个品种玉米根和茎叶中V 浓度均为:根远大于茎叶。益甜608 根部V 含量约占总量的63.75%—98.82%,美甜糯一号为73.91%—98.65%,可见,根系是玉米吸收和储存V 的主要部位,限制了V向地上部的运输。在不同时间,随着V 处理浓度增大,两个品种玉米根部和茎叶中富集的V 含量升高,与对照比较差异显著(P<0.05)。在V 胁迫4、8 和12 d,V 处理从5 mg·L-1增至30 mg·L-1时,与对照相比较,益甜608 玉米根部V 含量分别增加了35.7—178.5、25.5—75.5、67.1—195.0 倍,茎叶增加了0.8—5.3、1.4—32.8、2.4—13.4 倍;美甜糯一号根部分别增加了24.9—81.7、10.2—44.9、178.1—231.6 倍,茎叶部分别增加了2.8—10.4、4.6—21.3、6.9—20.4 倍。显然,两个玉米品种根部V 含量的增幅远大于茎叶部,胁迫12 d,美甜糯一号根部增幅大于益甜608。在相同V 浓度处理下,随着V 胁迫时间增加,玉米不同器官中V 含量增大后又降低,在胁迫8 d 时达最大。当V 处理为30 mg·L-1,胁迫8 d,益甜608 根部V 质量分数达305.74 μg·g-1,茎叶中V 质量分数为18.14 μg·g-1;美甜糯一号根部为319.80 μg·g-1,茎叶部为8.41 μg·g-1,与对照比较差异显著(P<0.05)。说明两个品种玉米对V吸收积累的特性差异在茎叶表现得更明显。
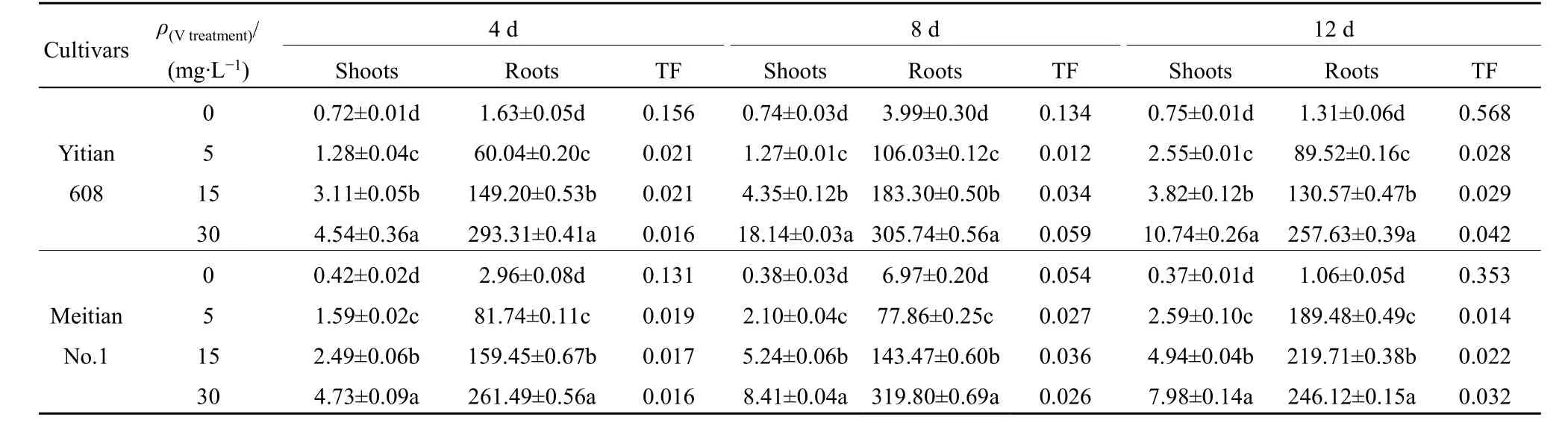
表1 两种玉米根和茎叶中V 质量分数 Table 1 Mass fraction of V in shoots and roots of two sweet maize varieties ω(Fresh)/(μg·g-1)
不同胁迫时间,玉米幼苗对V 的转运系数(TF值)变化不同。随着V 处理浓度增大,除对照外,在V 胁迫4 d,植物对V 的转运系数呈逐渐减小的趋势;在8 d 和12 d 的TF 值则有所增加,但始终小于对照(表1)。在V 处理浓度范围内,益甜608 的TF 值变化范围为0.568—0.016,美甜糯一号的TF 值在0.353—0.014。除对照外,随着胁迫时间增加,两个品种玉米幼苗的转运系数大多呈现升高后降低的变化趋势,在V 处理8 d,益甜608 和美甜糯一号的最大TF 值分别为0.059 和0.036,均小于0.1。两个品种玉米幼苗具有较低的TF 值,表明V 由植株根部向地上部转运的能力受限,植物的这种主动保护可使地上部组织免受过量V 的伤害。
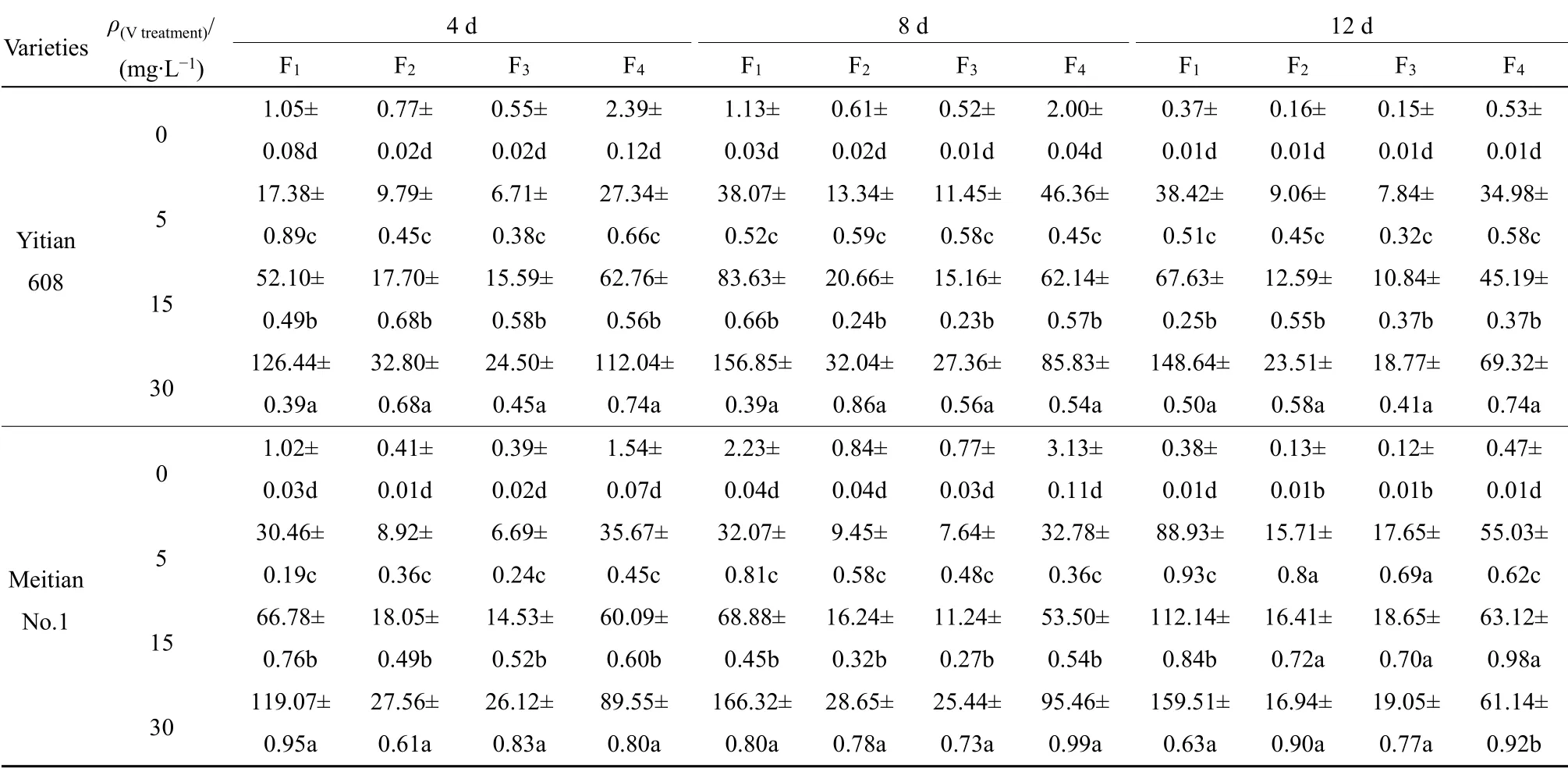
表2 两种玉米幼苗根中V 的亚细胞分布 Table 2 Subcellular distribution of V in roots of two sweet maize cultivars seedlings ω(Fresh)/(μg·g-1)
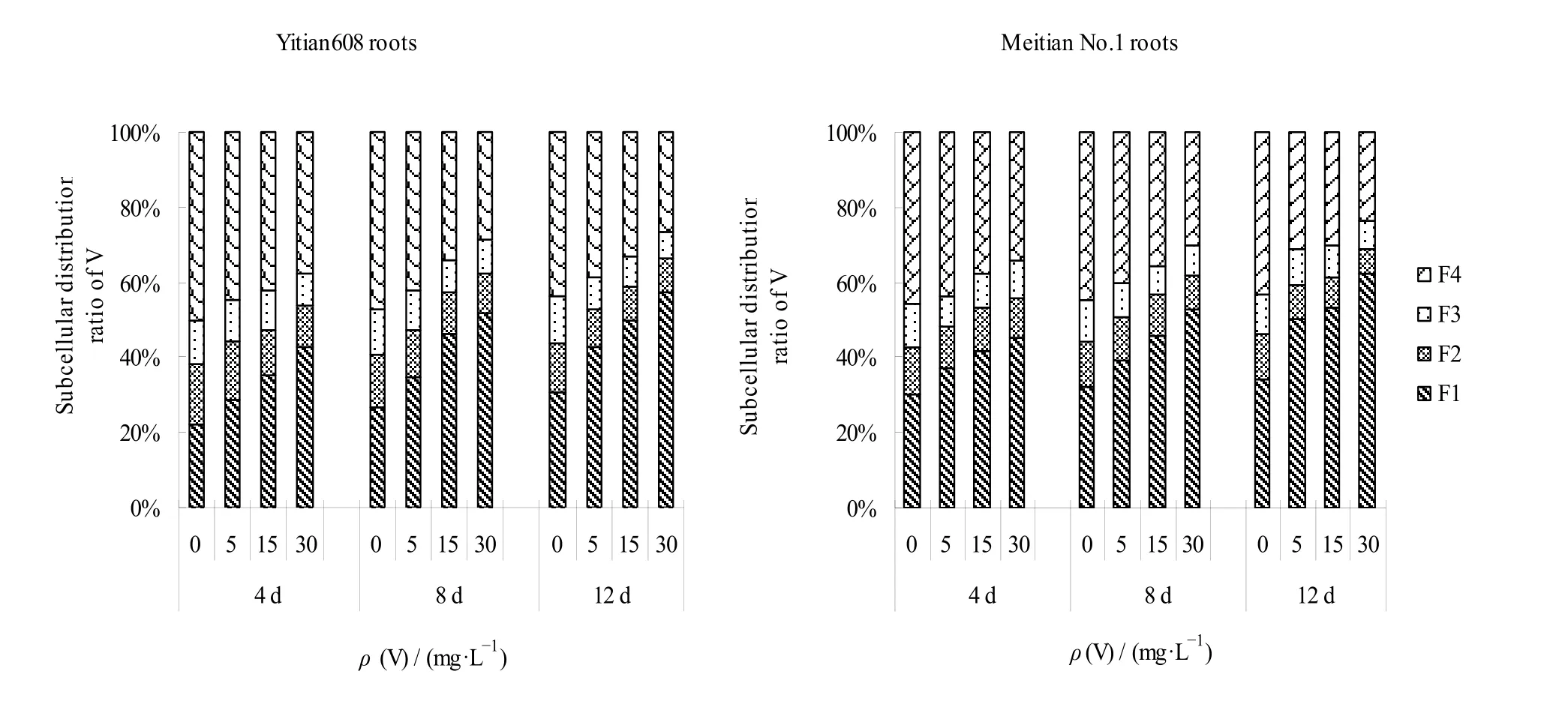
图1 两种玉米幼苗根中V 的亚细胞分配比例 Fig. 1 Subcellular distribution proportion of V in roots of two sweet maize varieties seedlings
2.2 不同品种玉米幼苗根中V 的亚细胞分布
亚细胞分析结果(表2,图1)可见,不同V浓度处理下,不同品种玉米根中V 的亚细胞分布比例顺序为:益甜608,细胞可溶性组分(F4)>细胞壁(F1)>细胞核(F2)>线粒体和叶绿体(F3),F4和F1二者比例之和为72.34%—83.75%;美甜糯一号,细胞壁(F1)>细胞可溶性组分(F4)>细胞核(F2)>线粒体和叶绿体(F3),F1和F4比例之和为76.24%—85.98%。两个品种玉米根系在细胞器(F2和F3)中分配较少(14.02%—27.66%),可见,在玉米根部,V 被固定在细胞壁和液泡区格化,进入细胞器的V 较少。随着V 处理浓度的增加,玉米根系亚细胞各组分V含量明显增加,与对照比较差异显著(P<0.05)(表2)。两个品种玉米根系亚细胞组分含量分配比变化相似,随着V 处理浓度增加,F1分配比例增大,而F4分配比例下降,益甜608 和美甜糯一号的F1分配比例分别从 22.04%和 30.31%增加到57.11%和62.15%,F4的从50.30%和45.94%降低到26.64%和23.82%,而F2和F3的分配比例变化不大(图1)。
2.3 不同品种玉米幼苗茎叶中V 的亚细胞分布
由表3 图2 可知,两个品种玉米茎叶的亚细胞各组分V 含量分配比例表现出不同规律,益甜608的细胞可溶性组分(F4)>细胞壁(F1)>细胞核(F2)=线粒体和叶绿体(F3),F4和F1二者比例之和为75.84%-82.34%;低质量浓度V 处理(<15 mg·L-1)时,美甜糯一号的细胞可溶性组分(F4)>细胞壁(F1)>细胞核(F2)=线粒体和叶绿体(F3),高质量浓度V 处理(≥15 mg·L-1)时,细胞壁(F1)>细胞可溶性组分(F4)>线粒体和叶绿体(F3)>细胞核(F2),F1和F4比例之和为71.94%-86.55%。随着V 处理浓度增大,美甜糯一号茎叶亚细胞各组分的V 含量差异均达显著水平(P<0.05),益甜608 除胁迫4 d 外,其余亚细胞组分V 含量在低浓度V 处理下差异均不显著(P<0.05)(表3)。与根部比较,不同品种玉米茎叶部亚细胞组分含量分配比变化有所不同,益甜608 的F1分配比下降(44.61%—25.88%),F4分配比升高(31.33%—55.29%);而美甜糯一号的F1分配比升高(23.52%—53.94%),F4的分配比下降(48.42%—32.62%),玉米茎叶的F2(12.55%—9.31%)和F3(11.71%—8.35%)的分配比变化较小(图2)。
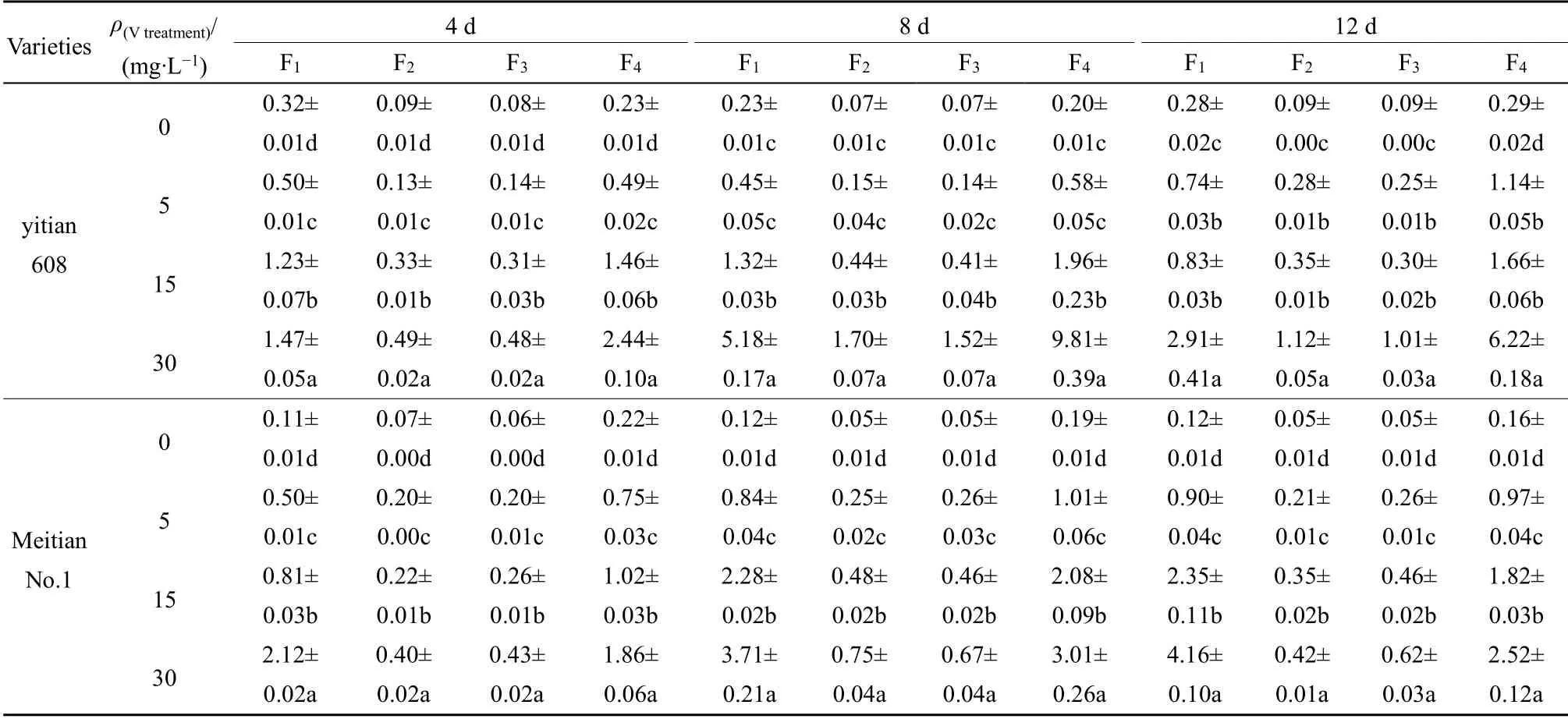
表3 两种玉米幼苗茎叶中V 的亚细胞分布 Table 3 Subcellular distribution of V in shoots of two sweet maize varieties seedlings ω(Fresh)/(μg·g-1)
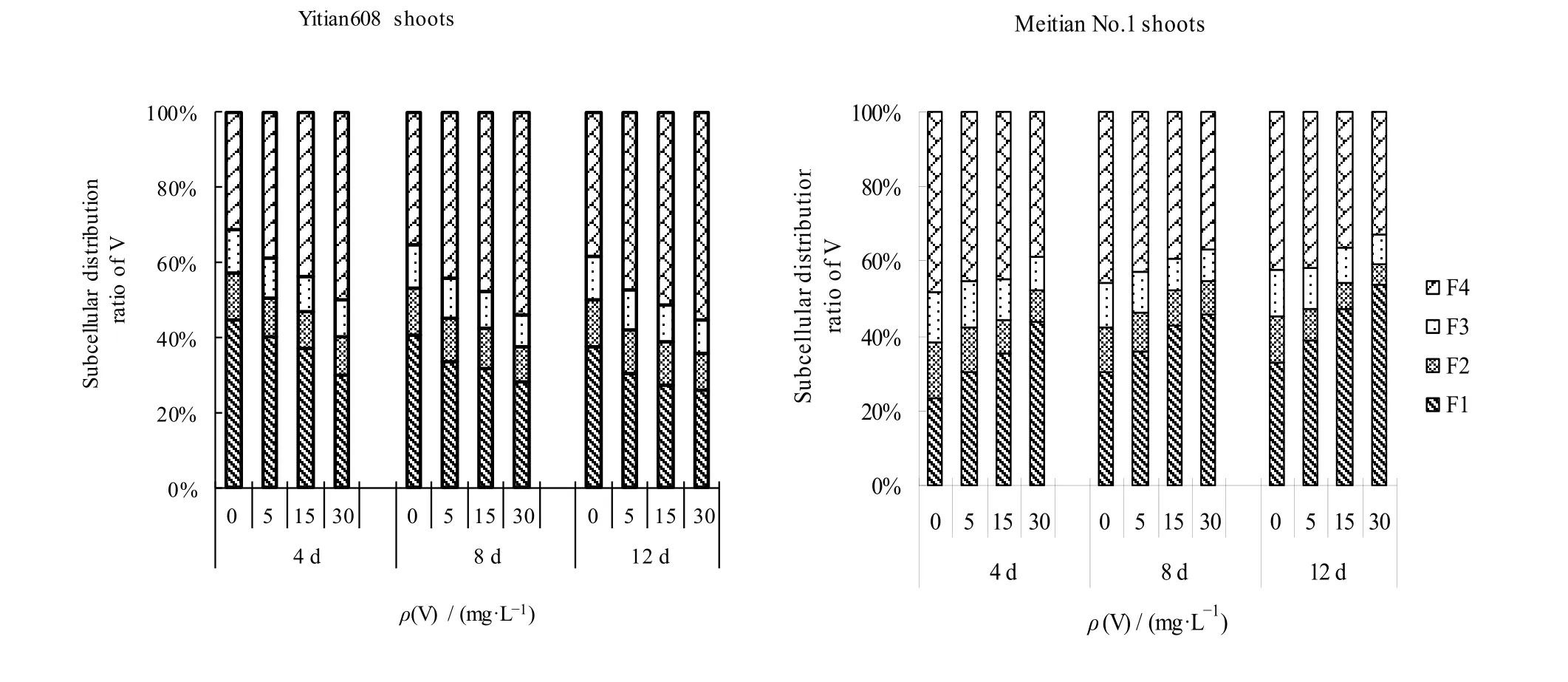
图2 两种玉米幼苗茎叶中V 的亚细胞分配比例 Fig. 2 Subcellular distribution proportion of V in shoots of two sweet maize varieties seedlings
2.4 不同胁迫时间对V 的亚细胞分布影响
在同一时间,随着V 浓度增加,玉米幼苗根和茎叶中亚细胞各组分V 含量均增加,尤其根部增加显著。在相同V 浓度处理下,随着时间增加,不同品种玉米幼苗V 的亚细胞组分分配变化不同,V 处理质量浓度为5 mg·L-1时,与4 d 比较,8 d 和12 d的益甜608 茎叶中,F1分配比分别下降了15.95%和24.13%,F4增加了14.11%和22.97%,根中F1比例增大了22.76%和49.81%,F4下降了4.96%和13.26%;美甜糯一号茎叶和根中,F1比例增加了18.19%和27.42%,5.01%和34.60%;F4比例下降了5.87%和7.96%,8.32%和28.88%。时间越长,植物根系中F1分配比增加越大,细胞壁对V 的沉积能力提高,成为V 的主要积累场所。
3 讨论
3.1 玉米幼苗对V 的吸收积累和耐性
一些研究结果(Teng et al.,2013;Qian et al.,2014;Chongkid et al.,2007)表明,植物首先在根部富集V,再向地上部转运,植物中V 浓度分布依次为:根>叶片>茎>子粒。司江英等(2008)研究也表明,在添加Cu 的土培玉米植物中,Cu 浓度分布以根>>叶>茎>子粒的顺序。本研究中,当用不同V 浓度处理,两个品种玉米幼苗根部对V 的积累均远大于茎叶部,根部分配比例高达约98%,这和前人的研究结果一致。显然,玉米植株根部是存储V的主要器官,玉米幼苗根部对V 的富集,可以降低V 在植物中的生物有效性,减少V 对地上部的毒害。从本研究时间动力学吸收特性来看,当V 胁迫时间较短(4 d),两个品种玉米幼苗生长较好,吸收积累V 量相差不大;随着时间增加,在不同V 浓度处理下,玉米幼苗根系和茎叶中V 含量均先增加后降低,这与李红婷等(2015)研究结果一致。可能是随着玉米植株培养时间增加,重金属在玉米根细胞内部和外部流速达到平衡,根系V 含量不再增加,使两种玉米幼苗对V 的吸收在第8 天时达最大值。在V 处理后期,玉米幼苗茎叶有枯萎症状,这也影响植物对V 的吸收,导致植物中V 含量降低。两个品种玉米各器官积累V 含量不同,表明不同品种玉米对V 的吸收机制不同。
植物的转运系数(TF)指植物将金属从根系到茎叶转运的能力,TF 值越大,植物对金属的转运能力越强。在胁迫时间较短(4 d)时,随着V 处理浓度增大,两个品种玉米幼苗TF 值减小,而胁迫时间延长,TF 值则有所增大,但除对照外TF 值均小于0.1,表明玉米幼苗对V 的迁移能力较低,这就有利于将V 固持在根系,从而避免V 对植物茎叶部组织的损伤。程海宽等(2015)研究结果表明,不同品种玉米对Pb 的耐受性不同,Muhammad et al.(2018)研究两种芥末基因型中,根系积累的V 都高于芽,而紫芥末对V 耐受性较高。同理,不同品种玉米对V 的吸收能力不同,其迁移能力和耐性也存在差异,当V 胁迫时间增加,美甜糯一号根系V含量比例增加达98.6%,而TF 值小于益甜608,表明对V 的耐性较好。可以推测玉米对V 的耐性机理为:外部适量V 不仅会诱导玉米幼苗体内谷胱甘肽(GSH)和植物螯合素(PCs)的合成,而且细胞质中的GSH 和PCs 能与V 生成稳定硫肽复合物,降低其对植物的危害,而玉米根系比茎叶合成更多的GSH 和PCs,抵御V 的毒害作用大于茎叶,使玉米的耐性增强(Hou et al.,2019)。玉米植物能将吸收的V 隔离在植物根部,阻止其向地上部分运输,不表现出明显的中毒症状,所以玉米植物为耐受非超富集型植物(Sekhar et al.,2011)。
3.2 V 在两种玉米幼苗中亚细胞分布
植物亚细胞组分中细胞壁的果胶、半纤维素和纤维素等大分子物质能与重金属结合将其固定(沈奕昕等,2018;Francine et al.,2014)。液泡是所有有毒物质在植物体内的终点站,转运至液泡的重金属与苹果酸、柠檬酸、草酸等结合以及与非蛋白巯基(NPTs)等鳌合,实现金属离子的区域化隔离,从而降低游离金属离子的毒害(Pittman,2005;Brunetti et al.,2015)。本研究结果表明,在外源V的胁迫下,两个品种玉米幼苗根系和茎叶中,细胞壁(F1)和细胞可溶性组分(F4)是植物V 积累的主要部位,二者比例之和根系达86%。茎叶部达87%,而进入细胞器的V 分配较少,表明玉米植株细胞壁V 的沉降和液泡区隔化作用是植物体内部抵御V 胁迫的两个重要过程。Fu et al.(2011)研究了美洲商陆对Cd 的耐性和解毒机制,在植物根和叶中Cd 主要位于可溶性部分和细胞壁中;Xu et al.(2018)研究发现,大豆幼苗根和叶中大约90%的Mo 在可溶性组分和细胞壁中积累,根富集的Mo 比叶片更高;杨素勤等(2015)研究结果表明,2 种小麦体内Pb 主要存在于细胞壁和细胞液中,其合量占小麦体内Pb 总量的72%—86%,本研究结果与前人相似。植物吸收的V 与细胞壁的多糖、蛋白质等结合,这就限制了金属的跨膜转运,以便维持植物细胞正常的生理活动。当细胞壁的活性基团与V 结合达到饱和后,多余的金属就会进入细胞内,此时植物细胞内的第二层保护机制被启动,细胞内分泌出的物质如高尔基体、内质网等可以将金属区隔化而固定在液泡中,防止金属伤害细胞器。玉米幼苗中V 还会与细胞质中的非蛋白巯基如谷胱甘肽或植物螯合素结合,使V 的毒性进一步降低(Hou et al.,2019)。美甜糯一号玉米根部F1和F4分配比之和大于益甜608,更有利于对V 的解毒作用。随着V 处理浓度的增加,两个品种根系细胞壁(F1)分配比例增加,可溶性组分(F4)比例下降,但茎叶中两组分比例变化不同,益甜608 的F1比例减小,F4比例增加,美甜糯一号的F1和F4变化相反。两个品种玉米幼苗细胞内细胞壁和可溶性组分分配比变化的此长彼消,是植物能够抵抗过量V 毒害的主要机制。随着V 胁迫时间延长,玉米根系F1的分配比例增加,表明植物根系细胞壁对V 沉积能力的提高,可以有效抑制V 向细胞器和地上部的转移,避免了过量V 进入玉米茎叶组织,大大降低其毒性。V 在玉米根中的大量积累可能与植物体内V 的亚细胞分布特征有很大关系。
唐杰等(2016)在研究不同水稻品种组织细胞中Cd 的转运分配中发现,低积累水稻品种根系F1、F4等含量显著高于高积累水稻品种;He et al.(2008)研究结果表明,在Cd 处理下,与野生型相比,突变型水稻植物生物量减少,可能是由于根系和叶片的细胞器分数中Cd 的分布较高,从而表现出更强的症状。这些结果说明同类植物品种不同表现出不同的耐性。益甜608 根细胞中的V 主要分配于细胞可溶部分(F4),而美甜糯一号则分布于细胞壁(F1),这可能与不同品种玉米幼苗对V 的耐性差异有关。要进一步阐明玉米对V 的解毒机制,还需结合超微结构的变化等深入研究。
4 结论
(1)两个品种玉米幼苗根系V 含量显著高于茎叶部,且具有较低的V 转运能力(TF<0.1),限制了V 在植物组织中的移动,避免了过量V 对玉米幼苗的伤害。玉米植物是V 的耐性植物。
(2)无论是在玉米幼苗根细胞或茎叶细胞中,V 主要富集在细胞壁(F1)和可溶性组分(F4),在细胞器中分配较少。随着V 处理浓度和胁迫时间增加,细胞壁固持和液泡区隔化的相互协调,是玉米植物应对V 胁迫的主要耐性机制。
(3)不同品种玉米对V 具有不同的解毒机制,美甜糯一号主要依靠细胞壁的沉积作用,而益甜608 则是可溶性组分的区隔化作用。甜糯一号玉米根部吸收的V 更多位于细胞非生理活动区,植物对V 的耐性更好。
猜你喜欢
现代园艺(2022年7期)2022-11-19
湖南电力(2021年4期)2021-11-05
四川劳动保障(2021年3期)2021-06-09
三农资讯半月报(2020年18期)2020-10-14
制造技术与机床(2019年9期)2019-09-10
数码世界(2018年1期)2018-12-23
浙江工业大学学报(2017年5期)2018-01-22
现代园艺(2017年13期)2018-01-19
广东农业科学(2017年5期)2017-08-29
腹腔镜外科杂志(2016年11期)2016-06-01
