
Cr6+对狼尾草种子萌发及幼苗生长的影响
2019-12-16 01:42葛元英陈新彤张鹏崔旭
江苏农业科学 2019年19期
葛元英 陈新彤 张鹏 崔旭

摘要:为了解狼尾草种子对重金属铬(Cr)污染的耐性,以狼尾草种子为试验材料,采用水培法,研究不同浓度的Cr6+胁迫对狼尾草种子萌发及幼苗生长的影响。结果表明,当Cr6+质量浓度较低时(≤25 mg/L),狼尾草种子的发芽率、发芽势、芽长、根长、发芽指数和活力指数等指标与对照组相比均无显著降低;当Cr6+质量浓度较高时(≥50 mg/L),狼尾草种子的发芽率、发芽势、芽长、根长、发芽指数和活力指数等指标均随Cr6+处理浓度的增加而降低。由研究结果可以看出,狼尾草种子对Cr6+有一定的耐受能力,可以应用于Cr6+轻度污染土壤的修复。
关键词:Cr6+;狼尾草;种子萌发;幼苗生长
中图分类号: S543+.9文献标志码: A
文章编号:1002-1302(2019)19-0126-03
收稿日期:2018-12-15
基金项目:山西省重点研发(指南)项目(编号:201603D21110-1-4)。
作者简介:葛元英(1977—),女,江苏泗阳人,硕士,实验师,主要从事污染土壤修复、实验室管理工作。E-mail:sxaugyy@126.com。
通信作者:崔 旭,博士,副教授,硕士生导师,主要从事污染土壤的治理工作。E-mail:sxaucx@126.com。
重金属铬(Cr)是地球地壳中常见的元素[1],存在于大气、水体和自然形成的土壤中,随着Cr在制革、染料、电镀和有机合成等行业的广泛应用,水体、土壤、生物均遭受了不同程度的污染[2]。近年来,常用的修复土壤重金属污染的方法包括物理、化学、生物修复等,其中生物修复中的植物修复与其他修复方法相比具有高效、经济和生态协调等优点,因而受到广泛应用[3]。但是,目前可应用的Cr超积累植物仍然不多,并且缺乏植物对重金属耐受能力的研究。种子萌发是植物生命的开始,萌发期是植物对环境最敏感的时期,它决定了幼苗的存活率、个体适合环境的程度以及植物生活史的表达等,进而影响到种群动态、植被分布和恢复等生态过程[4-7]。
狼尾草(Pennisetum alopecuroides)是一种在我国绝大部分地区常见的多年生禾本科草本植物,具有生命力旺盛、抗逆性强、根系发达、维护成本低廉、管理简单等优点[8-10]。目前,对狼尾草在Cr6+污染胁迫下种子萌发及其生长影响的研究较少[11]。因此,本研究选取狼尾草种子,通过在实验室水培条件下研究Cr6+对狼尾草种子萌发和生长的影响,旨在探索狼尾草对Cr的抗性机制及耐性表现,为选择具有高耐Cr能力同时能够富集Cr的植物物种提供一定的参考,也为后续利用植物修复Cr污染土壤的修复技术提供一定的科学依据。
1 材料与方法
1.1 材料
选择籽粒饱满完整、无病虫害、大小相近的狼尾草种子作为试验种子,所用的Cr6+试剂为K2Cr2O7(分析纯)。
1.2 种子萌发试验方法
试验于2018年4月在山西农业大学环境监测实验室进行。配制Cr6+浓度分别为0、5、10、25、50、100、200 mg/L的铬溶液,分别注入洗净、烘干、衬有滤纸的90 mm培养皿中,对照用去离子水。在每个培养皿中放入40粒狼尾草种子,每个浓度设3次重复。将加盖后的培养皿放入光照度为4 000 lx、光—暗周期为12 h—12 h、相对湿度为80%左右、25 ℃人工气候培养箱中进行恒温培养,每隔12h观察并记录发芽的种[HJ1.4mm]子数。发芽的第3天掀开培养皿盖,使其在透气状态下自然生长。每天定时加入一定量的去离子水,保持培养皿中的湿润状态,保证种子能够正常生长。当幼苗长出2张子叶时,测量种子的芽长、根长和鲜质量,计算发芽指数、活力指数等指标。
1.3 测定指标
本研究主要测定狼尾草种子的发芽势、 发芽率、鲜质量,以及芽长、根长、发芽指数等指标,每天记录发芽的种子数,以处理后第3天的数值计算种子的发芽势,以处理后第7天的数值计算种子的发芽率。并于第7天计算发芽指数,测定鲜质量,在每个培养皿中选取生长适中的10粒发芽种子,用直尺测量芽长、根长[12]。各项指标的计算方法如下[13]:
(1)发芽势。发芽势=(第3天发芽种子数/种子总数)×100%。
(2)发芽率。发芽率=(第7天发芽种子数/种子总数)×100%。
(3)鲜质量。从发芽的种子中挑出长势较好的3粒,晾干水分,用分析天平稱质量,求平均值,即为单株幼苗鲜质量。
(4)发芽指数。发芽指数=∑Gt/Dt。式中:Gt为td内的发芽数;Dt为相应的发芽时间,d。
(5)活力指数。活力指数=发芽指数×芽长度。
1.4 数据分析
试验数据用 Excel 2016软件进行分析。
2 结果与分析
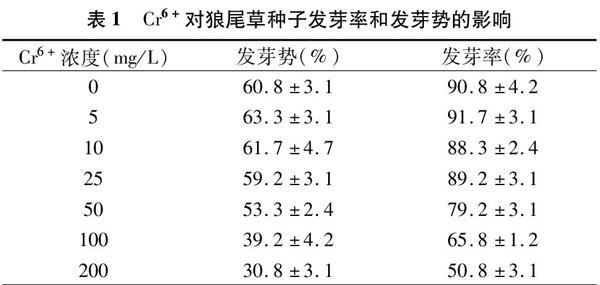
2.1 Cr6+对狼尾草种子萌发的影响
发芽势和发芽率是评价种子发芽速度和整齐度的重要指标[12],由表1可以看出,当Cr6+浓度≤25 mg/L时,狼尾草种子的发芽率和发芽势没有明显的变化,但是随着重金属Cr6+胁迫浓度的增大,发芽率和发芽势都表现出不同程度的下降。在5 mg/L Cr6+处理浓度下,狼尾草种子的发芽率、发芽势与对照相比,虽然上升幅度不是很高,但仍可表明,低浓度Cr6+处理对狼尾草种子的萌发有一定的刺激作用,促进了狼尾草种子的萌发;而当Cr6+处理浓度大于100 mg/L时,狼尾草种子的发芽率和发芽势都呈下降趋势,其中当Cr6+浓度为 200 mg/L 时,狼尾草种子发芽率、发芽势分别为对照的56%、51%。
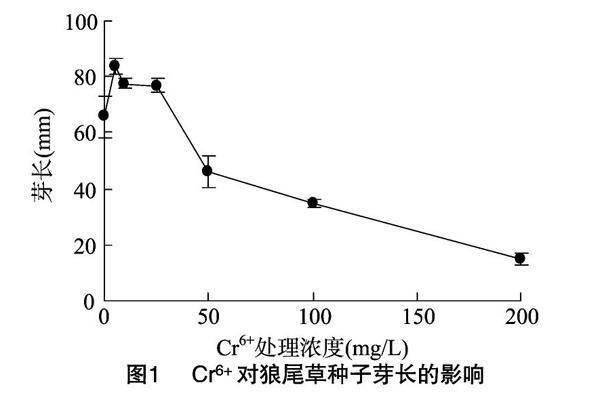
2.2 Cr6+对狼尾草芽长的影响
由图1可见,在低浓度(≤25 mg/L)Cr6+处理下,狼尾草种子的芽长均高于对照组;当Cr6+浓度为5 mg/L时,芽长最长。说明低浓度的Cr6+处理对狼尾草种子芽长有一定的促进作用,随着Cr6+浓度的增加,表现出一定的抑制作用,且随着胁迫浓度的增加,对狼尾草种子芽长的抑制作用也越强。狼尾草种子萌发的芽长变短以后,必然会影响狼尾草在随后生长期内的正常生长。
2.3 Cr6+对狼尾草根长的影响
由图2可知,在不同浓度的Cr6+处理下,狼尾草根长随Cr6+胁迫浓度增大,先略微上升,然后急剧下降,5~25 mg/L Cr6+处理对狼尾草种子根长有一定的促进作用,但Cr6+浓度≥50 mg/L 时,抑制作用就很明显。此外,由图1、图2可以看出,Cr6+对狼尾草种子根生长的抑制作用要大于对种子芽生长的抑制作用。
2.4 Cr6+对狼尾草种子发芽指数及活力指数的影响
发芽指数和活力指数也是能够反映种子萌发能力强弱的重要指标[12]。如图3所示,在低浓度(5~25 mg/L)Cr6+胁迫下,发芽指数随着Cr6+浓度的上升变化不大;当Cr6+胁迫浓度大于50 mg/L时,发芽指数表现出下降趋势,在200 mg/L Cr6+处理下的狼尾草种子发芽指数为对照的50.5%。活力指数是综合反映种子发芽速率和生长量的指标,是反映种子活力高低的一种可靠性指标[12]。由图4可以看出,狼尾草种子的活力指数变化规律与发芽指数的变化规律相似,在低浓度(5~25 mg/L)Cr6+胁迫下,狼尾草种子活力指数随着Cr6+浓度的上升而有所上升;当Cr6+胁迫浓度≥50 mg/L时,狼尾草种子活力指数呈下降趋势,在200 mg/L Cr6+处理下的狼尾草种子活力指数为对照的11.6%。这进一步说明,在低浓度Cr6+胁迫下,重金属对狼尾草种子萌发有促进作用,而高浓度的Cr6+对狼尾草种子萌发表现出一定的抑制作用。
3 讨论与结论
种子发芽率、发芽势、发芽指数和活力指数都是表示种子发芽程度及其生存能力大小的主要参数,根据这些参数在不同Cr6+质量浓度处理下的响应状况或被抑制程度,可以判断狼尾草种子在萌发期间耐受重金属Cr6+的状况[7,12]。不同浓度的Cr6+对狼尾草种子萌发及幼苗生长的影响一般表现为低浓度促进、高浓度抑制。在低浓度(≤25 mg/L)Cr6+处理下,Cr6+处理狼尾草种子的发芽率、发芽势、芽长、根长、发芽指数、活力指数与对照组间的差异不大,表明铬可能是植物生长过程中体内必需的微量元素之一。在高浓度(≥50 mg/L)Cr6+处理下,Cr6+处理对狼尾草种子的发芽率、发芽势、芽长、根长、发芽指数和活力指数等指标开始表现出抑制作用,且对各指标的影响相似。本研究结果与其他研究者对小麦种子、水稻种子、玉米种子和小香蒲种子的研究结果类似[14-16]。
Cr6+与植物作用时,首先接触到根部,因此重金属Cr6+进入植物体后,大部分会在根部蓄积,同时,根细胞壁中存在大量的交换位点,能够将重金属离子固定或吸收在这些位点上,从而阻止重金属离子向地上部分运移[17]。随着根部吸收或固定Cr6+离子的增加,加剧了细胞内染色体和核仁的破坏程度,这可能是抑制狼尾草种子根生长的主要原因。植物体中络合重金属最重要的部位就是根,因此根也是最易受重金属毒性影响的部位[7]。植物种子萌发所需要的能量和物质来源于贮存于种子中的物质氧化分解,这些物质的分解过程需要大量酶的参与,可能由于低质量浓度的Cr6+会与这些酶发生反应,产生物质刺激促进其萌发[18],而高浓度的Cr6+对植物种子萌发及幼苗生长表现为明显的抑制作用,可能由于高浓度的Cr6+增加了细胞膜透性,导致细胞内物质外渗[7,16]。
本研究结果表明,高浓度的Cr6+(≥50 mg/L)明显抑制了狼尾草种子的萌发和幼苗生长,且高浓度的Cr6+对狼尾草根生长的抑制作用大于对芽生长的抑制作用。但在低浓度(≤25 mg/L)Cr6+作用下,狼尾草表现出一定的耐受能力,这为选择具有高抗Cr能力,同时能够富集Cr的植物物种以及后续利用植物修复Cr污染的土壤净化治理提供了参考,但是对于Cr6+胁迫对狼尾草生长的影响机制,以及对于重金属Cr污染土壤的修复效率等还有待进一步的探索和系统的研究。
参考文献:
[1]Dahab O A,Al-Madfa H. Chromium distribution in waters and sediments of the eastern side of the Qatari Peninsula[J]. Science of the Total Environment,1997,196(1):1-11.
[2]Gao B,Gao L,Xu D Y,et al. Assessment of Cr pollution in tributary sediment cores in the Three Gorges Reservoir combining geochemical baseline and in situ DGT[J]. Science of the Total Environment,2018,628/629:241-248.
[3]孙约兵,周启星,郭观林. 植物修复重金属污染土壤的强化措施[J]. 环境工程学报,2007,1(3):103-110.
[4]王桔红,杜国祯,崔现亮,等. 青藏高原东缘61种常见木本植物种子萌发特性及其与生活史的关联[J]. 植物生态学报,2009,33(1):171-179.
[5]Donohue K,Dorn L,Griffith C,et al. The evolutionary ecology of seed germination of Arabidopsis thaliana:variable natural selection on germination timing[J]. Evolution,2005,59(4):758-770.
[6]陶 玲,任 珺,祝廣华,等. 重金属对植物种子萌发的影响研究进展[J]. 农业环境科学学报,2007,26(增刊1):52-57.
[7]Hou J,Liu G N,Xue W,et al. Seed germination,root elongation,root-tip mitosis,and micronucleus induction of five crop plants exposed to chromium in fluvo-aquic soil[J]. Environmental Toxicology and Chemistry,2014,33(3):671-676.
[8]朱秀红,侯国栋,茹广欣,等. 狼尾草对铬的积累及其抗氧化特性研究[J]. 河南农业大学学报,2017,51(3):330-334.
[9]陈 超,袁小环,滕文军,等. 狼尾草属植物生物学特性、生态适应性、观赏性和入侵风险关系的探讨[J]. 生态学杂志,2017,36(2):374-381.
[10]焦树英,李永强,沙依拉·沙尔合提,等. 干旱胁迫对3种狼尾草种子萌发和幼苗生长的影响[J]. 西北植物学报,2009,29(2):308-313.
[11]杨 阳,茹广欣,卢妍妍,等. 铬(Ⅵ)胁迫对狼尾草幼苗生长特性的影响[J]. 西部林业科学,2013,42(6):75-81.
[12]岑画梦,彭玲莉,杨 雪,等. Cd2+对狗牙根、假俭草种子萌發及幼苗生长的影响[J]. 草业学报,2015,24(5):100-107.
[13]王军涛,刘洪禄,吴文勇,等. 水培条件下重金属Cr(Ⅵ)对作物种子萌发影响的试验研究[J]. 农业工程学报,2008,24(6):222-225.
[14]吕笃康,李海冰,巴音山,等. 重金属铬及盐胁迫对小香蒲种子萌发的影响[J]. 种子,2013,32(2):72-75.
[15]康维钧,哈 婧,梁淑轩,等. 铬对萝卜种子发芽与根伸长抑制的生态毒性[J]. 河北科技大学学报,2005,26(4):322-325,329.
[16]卢志红,赵小敏,朱美英. 铬Cr6+对水稻种子萌发及幼苗生长的影响[J]. 中国土壤与肥料,2008(3):60-62.
[17]蒋光月,崔德杰. 重金属Cr对小白菜种子萌发及生长的影响[J]. 农业环境科学学报,2006,25(增刊1):85-88.
[18]张龙冲,张秋玲,赵 袁,等. 4种铬污染地植物的富集特征及其种子萌发对铬(Cr6+)处理的响应[J]. 河南农业大学学报,2016,50(4):550-557.
猜你喜欢
中国饲料(2022年5期)2022-04-26
牡丹(2021年11期)2021-07-20
现代农业科技(2016年20期)2016-12-20
广东饲料(2016年3期)2016-12-01
现代园艺(2016年17期)2016-10-17
作物研究(2014年6期)2014-03-01
