
花生子仁长宽及单仁重的遗传分析
2019-12-09 01:52张晓霞杨会张秀荣骆璐吕玉英张昆刘风珍万勇善
山东农业科学 2019年9期
张晓霞 杨会 张秀荣 骆璐 吕玉英 张昆 刘风珍 万勇善

摘要:本研究以栽培种花生品系05D677与品种中花12号为亲本材料,正反交构建2个F2分离群体,根据主基因+多基因分离分析方法,进行子仁性状遗传分析。结果表明:2个F2群体中花生子仁的仁长、仁宽及单仁重均存在广泛变异,表现出超亲遗传现象,且子仁性状频次均呈正态分布,具有数量性状特征,符合主基因+多基因遗传特点。仁长在2个F2群体中均符合3对主基因控制的加性-上位性遗传模型,其遗传率分别为80.0%和76.8%;仁宽符合1对具有加性效应的主基因+多基因混合遗传模型或2对具有显性上位效应的主基因+多基因混合遗传模型,主基因遗传率分别为2.0%、15.6%;单仁重符合具有完全等加性效应的主基因遗传模型或3对具有加性-上位性效应主基因遗传模型,主基因遗传率分别为52.0%、92.6%。
关键词:栽培种花生;子仁性状;主基因+多基因遗传分析;遗传率
中图分类号:S565.203.2文献标识号:A文章编号:1001-4942(2019)09-0073-07
Genetic Analysis of Kernel Length, Width and Single Kernel Weight in Peanut (Arachis hypogaea L.)
Zhang Xiaoxia, Yang Hui, Zhang Xiurong, Luo Lu, Lü Yuying, Zhang Kun, Liu Fengzhen, Wan Yongshan
(College of Agronomy, Shandong Agricultural University/State Key Laboratory of Crop Biology, Taian 271018, China)
Abstract In this study, the cultivated peanut line 05D677 and cultivar Zhonghua 12 were used as parents to construct two F2 isolation populations by reciprocal crosses, and the inheritance of kernel traits was analyzed according to the main gene+polygene separation analysis method. The results showed that there were wide variations in kernel length (KL), kernel width (KW) and single kernel weight (SKW) in the two F2 population showing super-parental inheritance, and the frequency of kernel traits were normal distribution with the property of quantitative traits, so they accorded with the characteristics of main gene + polygene inheritance. KL conformed to the additive-epistatic genetic model controlled by three major genes in the two F2 population with heritability of 80.0% and 76.8%, respectively. KW accorded with one pair of additive major gene + polygene mixed inheritance model or two pairs of dominant epistasis major gene + polygene mixed inheritance model. The heritability of major gene was 2.0% and 15.6%, respectively. The heritability of SKW was 52.0% and 92.6% respectively, which accorded with the main gene genetic model with complete equal additive effect or three pairs of main gene genetic model with additive-epistatic effect.
Keywords Cultivated peanut; Kernel trait; Major gene + polygene genetic analysis; Heritability
花生屬于豆科蝶形花亚科,起源于南美洲,由哥伦布发现新大陆后向全世界传播,是热带和亚热带区域各个国家广泛种植的经济和油料作物[1]。据USDA数据显示,2018年全世界花生年产量4 195万吨,中国、印度、美国为主要生产国。我国是世界上最大的花生生产国,且主要用于国内消费。花生仁在中国主要用于榨油消费、直接食用消费[2]。花生仁含有丰富的脂肪和蛋白质,是集营养、保健和防病功能于一身的健康食品[3]。随着经济水平和人民生活质量的提高,花生将会受到越来越多重视。
被子植物中,成熟的种子由胚、胚乳和种皮组成。这三种结构协调生长和发育,从而确定种子的大小和形态[4]。并且种子大小与基因[4]、转录因子[5]、激素[6]等多种因素相关。花生作为双子叶植物,其胚的发育类型为紫苑型,而胚乳只是一个短暂的结构,在成熟时只留下一层胚乳细胞,营养物质主要储存在子叶里面。胚乳虽然是成熟时的非储存组织,但在滋养发育胚胎方面起着重要作用,并且间接影响子仁大小[7]。前人利用主基因+多基因混合遗传模型主要是对花生进行荚果性状[8,9]、品质性状[10,11]、株高性状[12,13]及产量性状[14,15]等方面的遗传分析研究,而对子仁长宽的研究鲜有报道。
子仁大小是决定花生产量和品质的关键因素,解析子仁长、宽和仁重的遗传机制,挖掘关键基因对花生高产优质育种具有重要意义。本研究以子仁皱缩变异品系05D677以及子仁饱满的品种中花12号为材料,配制正交、反交组合,构建2个F2群体,进行子仁长宽、单仁重等性状的遗传分析,旨在研究花生子仁主要性状的遗传规律,为进一步的分子育种奠定基础。
1 材料与方法
1.1 试验材料
本试验选取山东农业大学选育的子仁皱缩变异品系05D677,以及中国农业科学院油料作物研究所培育的子仁饱满品种中花12号为材料。配制杂交组合05D677×中花12号(正交组合)、中花12号×05D677(反交组合),获得2个杂交组合的F1代,F1自交获得F2群体。利用2个F2群体进行子仁性状遗传分析。
1.2 试验设计
试验在山东农业大学农学实验站进行。2015年杂交,2016年大田种植F1代杂交种,2018年起垄覆膜种植F2群体。垄宽1.1 m,每垄2行,穴距0.2 m,双粒播种。出苗后间苗留单株,05D677×中花12号和中花12号×05D677群体单株数分别为586株和1 004株。
1.3 性状测定
将成熟后的花生按单株收获,悬挂晒干,剥壳后留子仁,对子仁的仁长、仁宽、单仁重等性状进行测定。依据《花生种质资源描述规范和数据标准》[16],用万深SC-G自动考种分析仪测定仁长、仁宽及单仁重。每个单株考察5个典型的子仁,计算平均值代表单株性状值。
1.4 数据处理
采用Microsoft Excel 2010进行数据处理。利用SPSS 20.0软件进行子仁数据的频次统计作图和相关性、差异显著性分析及遗传变异分析。根据植物数量性状主基因+多基因分离分析方法,进行子仁性状遗传模型分析[17-19]。利用G3DH.exe软件对P1、P2及F2群体进行联合分析,得到仁长、仁宽及单仁重3个性状的AIC值,再根据AIC值最小原则选出相应的遗传模型,并对备选模型进行5种适合性检验,包括均匀性检验(U21、U22、U23)、Smimov检验(nW2)和Kolmorov检验(Dn)。
2 结果与分析
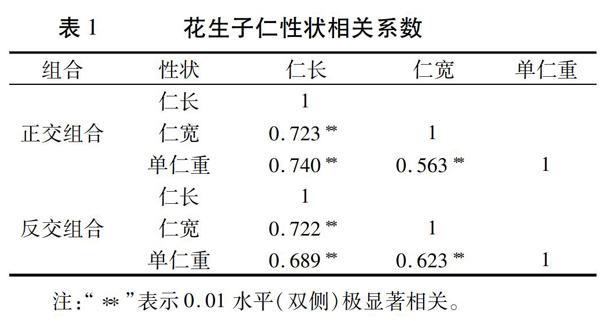
2.1 子仁性状相关性分析
由表1可知,正反交组合的2个F2群体中,仁长与仁宽均呈极显著正相关,相关系数分别为0.723和0.722;单仁重与仁长均呈极显著正相关,相关系数分别为0.740和0.689;单仁重与仁宽也均呈极显著正相关,相关系数为0.563和0.623。由此可以得出,单仁重与仁长的相关性更大。
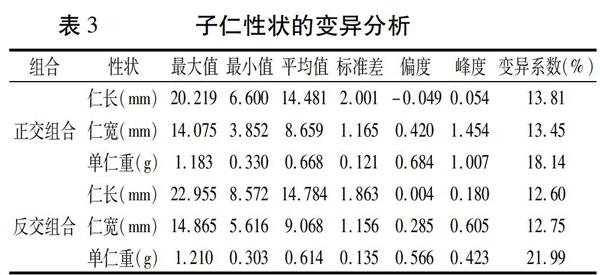
2个亲本的统计分析表明,05D677的仁长、仁宽及单仁重均低于中花12号,亲本间差异极显著(表2)。
在正交组合中,仁长、仁宽及单仁重3个性状均存在超亲遗传现象,变异广泛,三者的变异系数分别为13.81%、13.45%、18.14%。在反交组合中,仁长、仁宽及单仁重3个性状同样均存在超亲遗传现象,且仁长、仁宽及单仁重变异系数也均在10%以上,分别为12.60%、12.75%、21.99%(表3)。
由子仁性状的频次分布图(图1、图2)可以看出,在2个F2群体中仁长、仁宽及单仁重均呈正态分布,符合数量性状分布特征,说明该性状有可能是主基因和多基因控制,可以作进一步的相关遗传研究。结合表3可以看出,在正反交的2个F2群体中,仁长、仁宽及单仁重3个子仁性状的变异情况及频次分布均相似,表明其遗传受核基因控制。
2.3 子仁性狀最适遗传模型
利用G3DH.exe软件对3个子仁性状的表型数据进行分析,得到不同遗传模型包括AIC值在内的一系列参数,并根据AIC值最小原则选出相应的备选遗传模型,并对其进行模型5种适合性检验。
正反交组合中,仁长均为F-1模型的AIC值最小,说明仁长性状遗传符合3对主基因控制的加性-上位性遗传模型(表4)。
正交组合中,仁宽D-1模型的AIC值最小,说明仁宽性状遗传符合1对加性效应主基因+多基因混合遗传模型;在反交组合中,仁宽的适合模型为E-1-4,说明仁宽性状遗传符合2对显性上位效应主基因+多基因混合遗传模型(表5)。
正交组合中,单仁重F-3模型的AIC值最小,说明单仁重性状遗传符合3对完全等加性效应的主基因遗传模型;反交组合中,单仁重F-1模型的AIC值最小,说明单仁重性状遗传符合3对主基因控制的加性-上位性遗传模型(表6)。
2.4 子仁性状最适遗传模型的遗传参数
根据选定的3个性状的最适模型,对2个F2分离群体仁长、仁宽及单仁重3个子仁性状进行遗传参数估计(表7)。在正反交组合中仁长符合F-1模型,F2分离群体受3对具有加性-上位性的主基因控制,正交组合中3个主基因的加性效应值分别为-1.723、0.541、-1.16 主基因方差为3.204,主基因遗传率为80.00%;反交组合中加性效应值分别为2.237、-0.673、1.226,主基因方差为2.667,主基因遗传率为76.82%。
仁宽在正交组合中符合D-1模型,F2分离群体受1对具有加性效应的主基因+多基因控制,加性效应值为8.169;主基因方差为0.002,主基因遗传率为2.00%;仁宽在反交组合中符合E-1-4模型,F2分离群体受2对具有显性上位效应的主基因+多基因控制,加性效应值为2.147、6.342,主基因方差为0.209,主基因遗传率为15.60%。
单仁重在正交组合中符合F-3模型,F2分离群体受3对具有完全等加性效应的主基因控制,加性效应值为-0.076、-0.076、-0.076;主基因方差为0.008,主基因遗传率为52.00%;单仁重在反交组合中符合F-1模型,F2分离群体受3对具有加性-上位性的主基因控制,加性效应值为0.097、-0.020、0.044,主基因方差为0.018,主基因遗传率为92.60%。
3 讨论
本研究利用主基因+多基因分析法对花生2个F2群体的3个子仁性状进行遗传分析,结果表明,仁长在正反交组合中均符合F-1模型,即3对主基因控制的加性-上位性遗传模型,其遗传率分别为80.00%和76.82%,遗传率较高并较为接近,表明该性状可以稳定遗传。仁宽在正交组合中符合D-1模型,即1对具有加性效应的主基因+多基因混合遗传模型,主基因遗传率仅为2.00%;在反交组合中,3个备选遗传模型的AIC值相近,并且适合性检验的结果相同,但E-1-4模型的AIC值最小,即符合2对具有显性上位效应的主基因+多基因混合遗传模型,主基因遗传率为15.60%。仁宽性状在两群体中的遗传模型不一致,说明仁宽的遗传规律较复杂,两组合中其主基因遗传率均较低,表明仁宽性状受环境影响较大。
查阅花生相关文献发现,对于花生仁长及仁宽的主基因+多基因遗传分析鲜有报道。黄招德[20]在水稻中进行了粒长粒宽的研究,结果表明,粒长受一个主效显性基因控制,同时还有其它微效多基因修饰,而粒宽可能受多个主基因+多基因共同作用。樊庆琦等[21]在玉米中的研究表明,玉米粒长同时受主基因和多基因的影响。杨兴圣[22]在小麦中的研究表明,粒长符合加性-显性-上位性多基因模型,无主基因,粒宽符合2对加性-显性-上位性主基因+多基因遗传模型。
单仁重在2个组合中有一定的差异,正交组合中符合3对具有完全等加性效应的主基因遗传模型,反交组合中符合3对具有加性-上位性的主基因遗传模型。两组合中主基因的遗传率均较高,表明该性状可以稳定遗传,受环境影响较小。该结果与李兰周等[8]的研究结果一致;与刘佳琪等[9]的研究结果不一致,其研究表明在RIL群体中单仁重在3个环境中受1对或2对主基因控制。
综上所述,花生子仁长宽及单仁重的遗传机制比较复杂,不同群体、不同环境中的表现不一致,在花生子仁性状的改良中应继续深入研究。
4 结论
由栽培种花生品系05D677与品种中花12号构建的2个F2代分离群体中子仁的长宽及单仁重均表现为连续变异,有超亲遗传现象,且呈正态分布,说明其为数量性状,符合主基因+多基因遗传特点。
本研究初步确定了子仁3个主要性状的遗传模型:仁长在两群体中的最适模型均为3对主基因控制的加性-上位性遗传模型,其遗传率分别为80.00%和76.82%;仁宽的最适模型在两群体中分别为1对具有加性效应的主基因+多基因混合遗传模型和2对具有显性上位效应的主基因+多基因混合遗传模型,主基因遗传率分别為2.00%和15.60%;单仁重的最适模型在两群体中分别为3对具有完全等加性效应的主基因遗传模型和3对具有加性-上位性效应的主基因遗传模型,主基因遗传率分别为52.00%和92.60%。
参 考 文 献:
[1]王宝卿. 明清以来山东种植结构变迁及其影响研究[D]. 南京:南京农业大学,2006.
[2] 张怡. 中国花生生产布局变化研究[D]. 北京:中国农业大学,2015.
[3] 吕巨智,梁和,张智猛,等. 花生仁的营养成分及保健价值[J]. 中国食物与营养,2009(2):50-52.
[4] Li N,Li Y H. Signaling pathways of seed size control in plants[J]. Current Opinion in Plant Biology,2016,33:23-32.
[5] Luo M,Dennis E S,Berger F,et al. MINISEED3 (MINI3),a WRKY family gene,and HAIKU2 (IKU2),a leucine-rich repeat (LRR) KINASE gene,are regulators of seed size in Arabidopsis[J]. Proc. Natl. Acad. Sci. U.S.A.,2005,102(48):17531-17536.
[6] Li J,Nie X,Tan J L,et al. Integration of epigenetic and genetic controls of seed size by cytokinin in Arabidopsis[J]. Proc. Natl. Acad. Sci. U. S. A., 2013,110(38):15479-15484.
[7] Hehenberger E,Kradolfer D,Claudia K.Endosperm cellularization defines an important developmental transition for embryo development[J]. Development,2012,139(11):2031-2039.
[8] 李兰周,刘风珍,万勇善,等. 花生荚果和籽仁相关性状的主基因+多基因混合遗传模型分析[J].华北农学报,2013,28(5):116-123.
[9] 刘佳琪,李英杰,杨会,等. 栽培种花生RIL群体荚果及籽仁性状遗传变异分析[J]. 山东农业科学,2017,49(11):13-19.
[10]刘华. 栽培花生产量和品质相关性状遗传分析与QTL定位研究[D]. 郑州:河南农业大学,2011.
[11]张胜忠,焦坤,胡晓辉,等. 花生百仁质量和含油量的遗传分析[J]. 花生学报,2018,47(4):7-12.
[12]刘华,张新友,韩锁义,等. 花生主茎高、侧枝长的遗传分析及QTL检测[J]. 中国油料作物学报,2013,35(5):508-514.
[13]江建华,王嵩,汪清,等. 花生4个主要农艺性状的遗传分析[J]. 花生学报,2015,44(3):7-13.
[14]江建华,王嵩,王晓帅,等. 2个环境条件下花生产量性状的遗传分析[J]. 北方农业学报,2016,44(3):48-5 63.
[15]张新友. 栽培花生产量、品质和抗病性的遗传分析与QTL定位研究[D]. 杭州:浙江大学,2011.
[16]姜慧芳. 花生种质资源描述规范和数据标准[M]. 北京:中国农业出版社,2006.
[17]王建康,盖钧镒. 利用杂种F2世代鉴定数量性状主基因-多基因混合遗传模型并估计其遗传效应[J]. 遗传学报,1997(5):432-440.
[18]盖钧镒,王建康. 利用回交或F2∶[KG-*2/3]3家系世代鉴定数量性状主基因-多基因混合遗传模型[J]. 作物学报,1998,24(4):402-409.
[19]章元明,盖钧镒. 数量性状分离分析中分布参数估计的IECM算法[J]. 作物学报,2000,26(6):699-706.
[20]黄招德. 水稻籽粒性状的遗传分析及粒长基因GL-2的定位[D]. 福州:福建师范大学,2008.
[21]樊庆琦,杨克诚,乔善宝. 普通玉米3个籽粒性状的遗传分析[J]. 西南农业学报,2005,18(4):378-381.
[22]杨兴圣. 普通小麦籽粒特性的遗传分析[D]. 杨凌:西北农林科技大学,2013.
收稿日期:2019-05-09
基金项目:国家自然科学基金项目(31571711);山东省现代农业产业技术体系花生创新团队建设项目(SDAIT-04-03);山东省花生良种产业化工程项目(2017LZN033);国家现代农业产业技术体系建设专项(CARS-14);山东省重点研发计划项目(2019GNC106002)
作者简介:张曉霞(1993—),女,山东临朐人,在读硕士研究生,从事花生遗传改良与生物技术研究。E-mail:2274967222@qq.com
通讯作者:刘风珍(1966—),女,山东东阿人,博士生导师,教授,从事花生遗传育种研究。E-mail:liufz@sdau.edu.cn
万勇善(1960—),男,山东临沭人,博士生导师,教授,从事花生育种和栽培研究。E-mail:yswan@sdau.edu.cn
