
植物多酚对肉制品蛋白氧化的抑制机理及其延长货架期的应用
2019-12-03 01:08李文慧李应彪冉丽丹钟媛媛
食品科学 2019年21期
李文慧,刘 飞,李应彪,冉丽丹,侯 然,钟媛媛,董 娟,*
(1.石河子大学食品学院,新疆 石河子 832000;2.喀什大学生命与地理科学学院,新疆 喀什 844006)
目前,质量安全问题仍然是我国肉与肉制品加工和贮藏过程中面临的一个重要问题。其中微生物污染和蛋白氧化会导致肉与肉制品色泽变差、风味恶化、持水性及嫩度下降、消化率降低、货架期缩短等问题,这些不良变化不但给肉制品加工、生产、销售带来了难以估计的影响,而且对消费者的身体健康会产生不利的影响;因此,对肉与肉制品加工贮藏过程中蛋白氧化的有效控制及货架期延长具有重要的现实意义。以往研究者对肉制品脂质氧化的抑制机理方面已进行了大量研究,对肉制品中蛋白氧化控制方面研究较少,但近年来其成为了研究热点。蛋白质氧化修饰主要发生在氨基酸侧链上,包括巯基氧化、芳香族羟基化和羰基的形成[1-2]。氧化剂可以直接攻击蛋白质的骨架,引起蛋白质二级和三级结构的碎裂和构象变化。肉制品氧化诱导形成的二硫键、二酪氨酸和其他分子间的桥连作用导致蛋白质聚集,使肉与肉制品中蛋白质的营养价值下降[3-4]。还有研究表明,不适当的屠宰、操作和贮藏会导致肉中微生物繁殖和交叉污染,包括沙门氏菌和引起食源性疾病的单核细胞增生李斯特氏菌,造成严重的经济损失[5]。因此,寻找控制肉与肉制品加工与贮藏中蛋白质氧化和微生物污染的有效方法是非常必要的。
多年来,合成的抗氧化剂和抑菌剂能够作为自由基清除剂和金属螯合剂抑制肉中蛋白氧化和延长货架期[6]。有毒理学报告指出,合成的抗氧化剂和抑菌剂有潜在增加心血管疾病、肥胖症和癌症等疾病患病风险的可能[7-8]。随着人们生活水平的提高和健康意识的增强,绿色天然食品越来越受到消费者青睐,因此植物多酚取代合成抗氧化剂和抑菌剂已是大势所趋[9]。目前,富含酚类的植物提取物在肉制品中的应用已成为研究热点,如天然植物多酚在肉与肉制品中的抗氧化作用、植物多酚对微生物的抑制机理及其对肉制品品质的影响等研究具有重要的理论价值。
1 植物多酚分类、来源
天然多酚可以分为3大类,即黄酮类化合物、芪类和酚酸类[10]。黄酮类化合物是在植物和真菌中发现的一类次级代谢产物,在化学结构上,黄酮骨架由2 个苯环和1 个含氧杂环组成,具有15碳(C6-C3-C6主链)骨架结构。研究者们已经从各种植物中分离出5 000多种黄酮类化合物,大致分为黄烷醇、黄烷酮、黄酮、异黄酮、黄酮醇和花青素[11]。花青素是黄酮类化合物最重要的亚类,广泛分布于水果和蔬菜中[12-14]。茶叶和可可豆中发现的主要黄酮类是单体黄烷-3-醇(儿茶素)及其衍生物(表儿茶素、没食子儿茶素)[15]。木本植物、水果、啤酒和葡萄酒中的主要黄酮类组分是浓缩单宁和原花青素,这些聚合物可以沉淀蛋白质并具有抗菌活性。
芪类是14碳(C6-C2-C6)骨架,具有单体、低聚体和聚合物结构[16]。白藜芦醇属于芪类单体,具有多种生物活性,如抗氧化、抗癌和抗菌活性。在葡萄、浆果、花生、可可以及红葡萄酒中均发现了白藜芦醇[17]。
酚酸是植物界中广泛分布的次生代谢产物,可以在碱性介质或酶促反应中水解。酚酸是一类苯环上含有一个或多个羟基的化合物,按结构可分为苯甲酸和苯乙烯型(羟基肉桂酸)[18]。羟基苯甲酸包括没食子酸、对羟基苯甲酸、原儿茶酸、香草酸和丁香酸。最常见的羟基肉桂酸包括咖啡酸、阿魏酸、对香豆酸和芥子酸[19]。
2 植物多酚对肉制品蛋白氧化的抑制机理
蛋白氧化定义为通过活性氧(reactive oxygen species,ROS)直接诱导,或通过氧化应激的次级产物间接诱导蛋白质共价修饰,一般来说ROS可以介导蛋白质氧化,即自由基(羟自由基(•OH)、超氧阴离子自由基()、烷过氧基(ROO•)、烷氧基(RO•)、一氧化氮自由基(NO•)及活性物种、多不饱和脂肪酸遇高温形成的自由基)、氧的非自由基衍生物(H2O2、HOCl、过氧化物)或过渡金属离子(Fe2+、Cu2+)会促进蛋白氧化[20-21]。研究表明植物中的酚类化合物与肉和肉制品中蛋白质的共价结合可以抑制蛋白氧化,植物多酚的抗氧化能力与其含量相关[22]。植物多酚抑制蛋白质氧化机理可归纳为以下几点。
2.1 多酚作为自由基和活性物质的清除剂
蛋白氧化过程涉及到自由基的形成和反应,肉制品中的铁离子可与H2O2反应产生具有强氧化能力的•OH,NO带有自由基,化学性质非常活泼,研究发现NO为蛋白质氧化的前体物[23]。由于植物多酚中苯环上存在羟基结构,可通过氢原子转移机制将酚羟基基团上的氢原子向自由基转移而清除自由基(Ar—OH+R•→ArO•+RH)[24-25],di Meo等[26-27]提出了4 种可能导致这种反应的机制:1)氢原子转移和质子耦合电子转移(Ar—O··H→Ar—O•+H•;R•+H•→RH)。该机制是纯氢原子的直接转移,自由基清除活性完全由酚羟基基团上O—H键的均裂解离控制,多酚电子转移到自由基单独占据的分子轨道。2)电子转移 质子转移。根据自由基与酚羟基反应的氧化体系,这种机制作为次要机制发生,是由电子转移引起(ArOH+R•→ArOH•++R-),然后是质子释放(ArOH•++R-→ArO•+RH)。3)连续质子损失-电子转移。该机制由质子损失引起(ArO-H→ArO-+H+),然后多酚阴离子进行电子转移(ArO-+R•→ArO•+R-;R-+H+→RH)。4)在放射溶液中,加合物形成(ArO—H+R•→[ArOH-R]•;[ArOH-R]•→ArO•+RH),这种机制对•OH具有相对特异性,该基团可以在双键和芳香环上加成。
Xiong Youling L.[28]研究指出将植物酚类(儿茶素、咖啡酸、绿原酸、迷迭香)加入牛肉馅饼和炸丸子中,植物多酚会以清除自由基的方式来抑制蛋白质的氧化修饰。Soladoye等[3]研究发现,自由基与肉制品蛋白质多肽主链的特定位点发生反应,导致其碎裂和氨基酸侧链的修饰。Gülçin等[29]通过红外单线态氧(1O2)化学发光法证明,作为清除剂,单宁酸比合成抗氧化剂丁基羟基茴香醚、二丁基羟基甲苯和α-生育酚更有效,1O2是肉制品加工中常出现的一种自由基和高反应性分子,多酚(单宁酸)可作为电子和质子供体使单线态氧氧化程度大大降低。
植物多酚在肉与肉制品中抗氧化性与其清除自由基活性有关[30-32]。在肉与肉制品加工、贮藏和光照过程中会导致活性物质形成,从而破坏蛋白质结构、功能特性,并降低产品营养价值和感官可接受性,甚至产生有毒化合物。逯阳[33]认为,植物酶的活性非常强,其中超氧化物歧化酶可将ROS、活性氮转变为相对稳定的化合物,减少自由基的产生从而发挥抗氧化作用。Zhang Yumeng等[34]研究发现,肌原纤维蛋白在肉制品加工过程中很容易受到ROS的攻击,β-环糊精(β-cyclodextrins,β-CDs)与表没食子儿茶素没食子酸酯(epigallocatechin gallate,EGCG)相互作用,形成的β-CD-EGCG复合物可以减少ROS攻击并提高肉制品的凝胶强度。
2.2 多酚作为非自由基衍生物的清除剂
H2O2在肉制品中是一种具有强氧化作用的非自由基衍生物,在Fe2+存在的情况下,其可将H2O2还原产生•OH,由于酚类化合物是良好的电子和质子供体,植物多酚将•OH转化成水,从而清除非自由基,达到抑制肉和肉制品蛋白氧化的目的[16]。Fenton反应被认为是•OH的主要来源,其中Fe3+是肌肉蛋白氧化的起始因子。除了Fe3+外,其他过渡金属离子在H2O2存在的条件下也会引发蛋白质氧化[35]。几乎所有的过渡金属离子都有能力在不同的氧化态中发挥作用,在活性氧化还原态中,过渡金属离子可以作为许多生物分子自动氧化的催化剂。在大多数情况下,具有氧化还原活性的过渡金属离子与H2O2反应产生羟自由基进而引发氧化,称为Fenton反应[36]。
2.3 多酚作为过渡金属离子的螯合剂
Adjimani等[37]使用咖啡酸-铁螯合剂作为体外抗氧化剂的实验结果表明,咖啡酸-铁螯合剂可通过清除ROS和自由基而起到抗氧化剂的作用,减少由Fenton反应产生•OH的量,从而减少肉类蛋白质的氧化。Gülçin[38]认为,Fe3+是血红素和铁蛋白释放出来的最丰富的离子,是蛋白质氧化的重要催化剂,当Fe3+被植物多酚螯合时,它可能失去其助氧化剂性质。白藜芦醇的结构及其与金属离子螯合作用的结合位置见图1A,白藜芦醇可以利用其两个羟基螯合Fe2+。同时,Liu Yuanzhen等[39]利用电喷雾串联质谱研究了第一过渡金属离子(Fe3+、Co2+、Ni2+、Cu2+和Zn2+)对金属离子-类黄酮螯合物形成和稳定性的影响,通过全扫描质谱,发现Fe2+与槲皮素主要形成化学计量比为1∶2和2∶3的金属-类黄酮螯合物,而Cu2+和槲皮素主要形成化学计量比为1∶1和1∶2的金属-类黄酮螯合物(图1B);螯合分子的数量会随着金属离子半径的减小而减少,槲皮素通过其5-OH和4-羰基与二价金属离子形成不同的络合物。脂质氧化中引发自由基链的主要途径是通过H2O2介导血红素蛋白的降解,使肉制品释放游离铁,或者将血红素铁迅速地还原为铁氰化物自由基,肉制品中含血红素蛋白的铁在脂质过氧化的催化中起着关键作用[40]。

图 1 白藜芦醇(A)、槲皮素(B)与金属离子的螯合Fig. 1 Chelation of resveratrol (A) or quercetin (B) with metal ions
2.4 多酚作为高铁肌红蛋白的还原剂
氧合肌红蛋白(MbO2)是一种含有血红素Fe(II)的蛋白质,其使肉呈鲜红色。MbO2中Fe(II)被氧化成Fe(III),MbO2转化成高铁肌红蛋白(MetMb),MetMb呈褐色,降低了肉的市场价值。除了颜色变化之外,MbO2的氧化会产生超氧化物自由基,由此加速了肉的氧化变质,因此控制MbO2的氧化对于维持肉品质至关重要。Lorrain等[41]研究发现,当多酚(芥子酸、儿茶素、紫杉叶素、桑色素和阿魏酸)浓度为600 μmol/L时会降低肉制品中MetMb含量,可以抑制MbO2氧化、减少MetMb的产生,使肉与肉制品保持良好的色泽。Inai等[42]通过研究植物相关酚类化合物对MetMb还原的影响发现,酚羟基可通过捕获自由基来抑制肉制品的氧化,且单酚化合物的抗氧化活性不是很强,但邻位和对位二酚化合物在体外的抗氧化活性很强。但在还原MetMb过程中,酚羟基容易受空气氧化形成稳定的醌或半醌自由基,而醌或半醌自由基因含有α、β-烯酮具有亲电子活性,会促进MbO2的氧化,使其不稳定。Masuda[43]、Honda[44]等认为,植物多酚的羟基结构并不是抑制MbO2氧化的必要条件,在等量的半胱氨酸存在下,半胱氨酸中的巯基具有有效的亲核性和自由基反应性,会与氧化后的多酚反应,得到的产物将MetMb还原成MbO2。多酚结构中半胱氨酰被硫取代的数量影响着MetMb还原活性和MbO2的维持。具有两个半胱氨酸残基的羟基酪醇衍生物具有最高的MetMb还原活性并且对MbO2没有影响。以儿茶素为例,其还原MetMb的机理具体见图2。
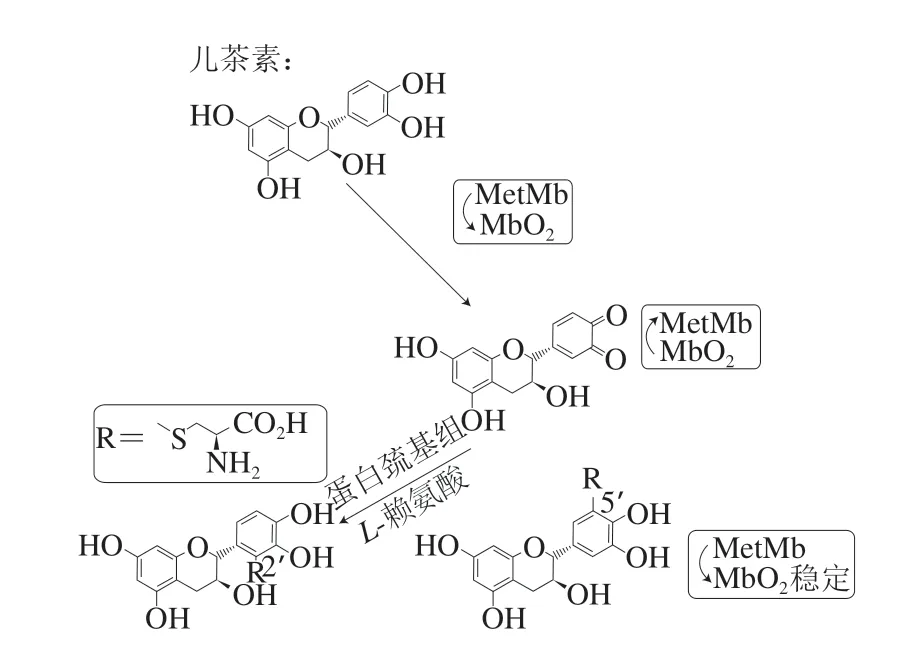
图 2 儿茶素还原MetMb的机理Fig. 2 Mechanism of action of catechin as a reducing agent for MetMb
3 植物多酚在延长肉制品货架期中的应用
近年来对植物多酚抑菌机理的探究比较热门,由于多酚结构多样,不可能预测每个类别结构特征多酚的抑菌活性以及细菌与多酚之间的作用机理[45]。因此,植物多酚的抑菌活性机理仍需进一步研究。植物多酚抗菌机理可归纳为以下几点。
3.1 与细菌细胞壁的相互作用
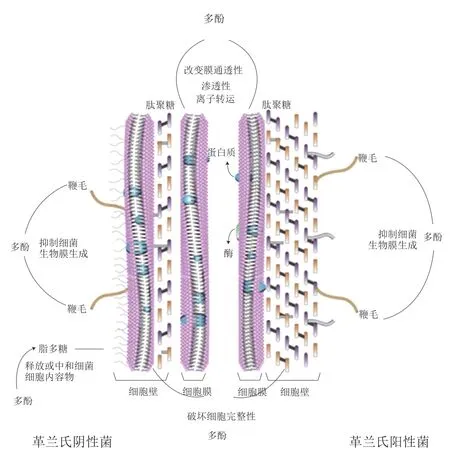
图 3 多酚对革兰氏阴性菌和革兰氏阳性菌的抑菌机理Fig. 3 Antibacterial mechanisms of polyphenols against Gram-negative and Gram-positive bacteria
革兰氏阴性菌和革兰氏阳性菌的细胞壁不同,革兰氏阴性菌细胞壁由肽聚糖和外层薄膜组成,革兰氏阳性菌由一层厚的肽聚糖和磷壁酸组成。细胞壁在保护细胞的渗透压中起重要作用;因此,对细胞壁的任何损害都会降低细胞对高离子强度和低渗透压的耐受性[46]。许多研究人员已经证明了多酚与细菌细胞壁相互作用的能力,由于细胞壁组成的差异,革兰氏阴性细菌对多酚的抵抗作用更强[47-48]。儿茶素、EGCG能够直接结合金黄色葡萄球菌的肽聚糖,影响细胞完整性,从而降低细胞对高离子强度和低渗透压的耐受性。Bukvički等[49]将精油加入猪肉后针对单核细胞增生李斯特氏菌进行研究,结果发现用精油处理的猪肉成功地抑制了猪肉中单核细胞增生李斯特氏菌的增殖,使猪肉的颜色和风味得到改善。植物多酚对革兰氏阳性菌和革兰氏阴性菌的抑菌机理见图3。
3.2 与细菌细胞膜的相互作用
董璐[50]、钱丽红[51]、王慧敏[52]等研究发现,茶多酚能增加大肠杆菌、金黄色葡萄球菌和铜绿假单胞菌、腐败希瓦氏菌的细胞膜通透性,影响膜流动性,阻碍细菌蛋白质的正常表达,破坏细菌细胞膜的脂质层,使细菌发生形态学改变,从而影响细菌细胞结构以及酶的催化活性,抑制呼吸作用和改变离子转运过程,最终导致细菌正常生理功能下降,产生良好的抑菌活性,为茶多酚应用在肉制品和水产品上延长其货架期提供了理论依据。Phan等[53]研究发现,黄酮类化合物和反式芪化合物通过两种完全相反的方式改变膜性质,由于没食子酸酯、没食子酰基和羟基的存在,类黄酮能够与细菌膜脂质双层的亲水头部基团形成氢键磷脂,形成的氢键磷脂主要影响脂质双层的亲水区域,介导磷脂的聚集,从而导致膜面积减小并使膜的刚性更强;具有开放苄基环结构的反式芪倾向于深入脂质双层的疏水内部并增加膜面积,使膜的流动性增加。
3.3 抑制生物膜的形成
生物膜是肉类在屠宰和处理过程中,腐败菌和病原微生物接触并附着在肉类表面时为适应生存环境由菌体分泌而形成的一种胞外物质[54]。群体感应是细菌细胞间通过化学信号交流信息的一种有效通讯方式,可以调节一些功能(如毒力、抗生素生成和生物膜形成),在孢子形成和分散过程中执行专门的任务,当浓度达到阈值时,这些自动诱导物与转录调节物相互作用,允许特定表达一组基因,以响应高浓度的该自动诱导物[55-57]。此外,大多数细菌已经发展出精密的机制来黏附肉类固体表面和协调内部的活动,从而建立复杂的菌落,生物膜能够保护细菌免受环境污染和攻击,改善其对许多不同宿主的依赖,有效地获取氧气、营养物质维护、拆卸结构化的多细胞群落关键取决于细胞之间的化学通讯[58]。常见的肉类腐败细菌(假单胞菌、乳酸杆菌)和食源性致病菌(沙门氏菌、李斯特菌和大肠杆菌)在肉制品表面易形成生物膜,导致严重的卫生问题[59]。肖洁等[60]研究发现,茶多酚具有很强的抑制大肠杆菌(O157:H7)生物膜的能力。Bostanghadiri[61]、Giaouris[62]等发现,反式白藜芦醇和精油对附着到肉表面的细菌生物膜具有显著的抑制作用,其应用可减少肉类细菌、食源性细菌数量和延长肉类货架期。细菌生物膜是微生物细胞彼此黏附在一起产生的,其具体结构和组成取决于物种和环境条件。生物膜黏附性使细菌难以从水管或食品加工设备表面完全清除,导致食源性疾病和食物腐败的蔓延。有研究表明,荧光假单胞菌使用群体感应来控制生物膜形成和运动,肉桂醛可以通过与荧光假单胞菌的LuxR型蛋白相互作用的方式来抑制群体感应,对荧光假单胞菌生物膜形成明显的抑制[63]。鞭毛是细菌重要的毒力因子,除了提供细菌运动相关功能外,还能在细菌生物膜形成初期及菌落生长过程中加快细菌黏附速度,当细菌黏附于固体表面后,细菌产生的胞外多糖对生物膜的形成和稳定结构起着重要作用,因此鞭毛在细菌生物膜形成过程中具有重要作用[64]。
3.4 抑制生物大分子的合成
抑制蛋白质合成是多酚抑菌作用的主要机理。多酚抑制细菌细胞的蛋白质、DNA和RNA等生物大分子的合成,使细菌的遗传表达受到影响,细胞形态发生改变[65]。Gradisar等[66]研究表明,DNA旋转酶是一种依赖于ATPI型的DNA拓扑异构酶,EGCG可通过抑制DNA旋转酶的活性,使其失活而导致细菌死亡。大蒜素可以快速抑制鼠伤寒沙门氏菌RNA合成,并且可以部分抑制DNA合成,抑制微生物中的许多细菌酶活性,从而降低其毒力,因此大蒜素可用作抑菌剂或杀菌剂化合物[67]。
3.5 抑制细菌酶的活性
环二腺苷酸(cyclic di-adenosine monophosphate,c-di-AMP)是细菌核苷酸的第二信使分子,其作用包括控制不同细菌中的细胞壁动态平衡、细菌生长代谢、生物被膜形成、细菌毒力因子等。在枯草芽孢杆菌、金黄色葡萄球菌、肺炎链球菌以及单核细胞增生李斯特菌等常见致病菌中均发现了c-di-AMP的存在[68]。Opoku-Temrng等[69]研究指出, DisA是细菌中最为普遍存在的c-di-AMP合成酶,c-di-AMP合成酶可利用ADP合成c-di-AMP;14 种植物多酚对枯草芽孢杆菌的DisA活性具有抑制作用,可通过抑制c-di-AMP的合成发挥抑菌作用。
4 结 语
植物多酚可作为反应性物质清除剂、非自由基衍生物清除剂、过渡金属离子螯合剂和高铁肌红蛋白还原剂抑制蛋白氧化,同时多酚物质还可以通过与细菌细胞壁和细胞膜相互作用、抑制生物膜的形成等形式抑制肉制品中细菌物质的生长繁殖,来减缓肉与肉制品中蛋白质的氧化,从而延长肉与肉制品货架期[70]。植物多酚为植物体内复杂酚类的次生代谢物,具有多元酚结构,主要存在于植物的皮、根、叶、果实中,在植物中的含量仅次于纤维素、半纤维素和木质素。植物多酚具有抗氧化、抗菌、抗炎症、抗过敏等生理功效,近年来成为人们关注和研究的热点。随着人们对纯天然食品的重视,植物多酚或者富含多酚的植物提取物已经被越来越多地应用于肉制品中,用来抑制蛋白质和脂肪氧化、提高肉制品品质及营养价值,延长货架期。植物多酚抑菌机理仍存在许多争议,因其结构不同,抑菌机制应进行更深入系统的研究,其未来的研究前景仍十分广阔。
猜你喜欢
现代临床医学(2021年5期)2021-11-02
昆明医科大学学报(2021年4期)2021-07-23
生物技术通报(2021年2期)2021-04-26
今日农业(2020年23期)2020-12-31
食品安全导刊(2020年33期)2020-12-04
食品安全导刊(2020年36期)2020-12-02
中成药(2018年2期)2018-05-09
中国塑料(2016年8期)2016-06-27
中国塑料(2016年12期)2016-06-15
人间(2015年11期)2016-01-09
