
银叶树群落优势树种分布与土壤环境、群落竞争的关系研究
2019-11-30 06:36:06郭乐东黄芳芳张卫强甘先华
生态环境学报 2019年10期
郭乐东,黄芳芳,张卫强,甘先华
广东省森林培育与保护利用重点实验室/广东省林业科学研究院,广东 广州 510520
植物群落中物种生长和种群更新受到多个因素影响。其中,土壤是森林生态系统中的重要组成部分,为物种生长提供必要的养分和水分,对植物生长和更新有着重要作用;同时,土壤还可通过影响群落物种竞争关系、群落构成及演替格局,影响群落发展动态(Paoli et al.,2006;Toriyama et al.,2007)。群落中不同物种对土壤环境的需求存在差异(Porazinska et al.,2003),良好的土壤环境,如充足的养分和有利的土壤微生物群落,有助于群落物种稳定发展,而恶劣的土壤环境则可能影响植物生长,降低群落物种多样性(Isermann,2005;Lai et al.,2018)。不同的土壤环境往往支持不同类型群落的发展,反映出物种对土壤环境响应的差异性(陈林等,2019)。
种间关系是影响群落物种生长和更新的重要生物因素(Martorell et al.,2014)。由于植物营固着生长,且对资源往往表现为刚性需求,物种间竞争是种间关系的重要形式,并显著影响物种在群落中的分布格局(Ding et al.,2019)。竞争和土壤环境存在相互作用(Lekberg et al.,2018),即不同土壤环境下群落竞争程度不一。良好的土壤环境可缓解竞争,而恶劣的土壤环境则可能加剧竞争。此外,除了通过优先占领及竞争性状表达等直接形式,竞争也可能通过间接形式,如改变土壤生物和非生物环境来影响物种生长(van de Voorde et al.,2011)。
古树群落对研究群落演替格局和物种更新具有重要的研究价值。但是,持续的人为干扰对现存古树群落产生了严重影响,加强对古树群落的保护和恢复显得尤为迫切。银叶树(Heritiera littoralis)隶属于梧桐科银叶树属,是热带、亚热带海岸半红树植物,既能生长于潮间带,也可生长于陆地上,不具有气生根、胎萌及耐盐高渗压等典型红树植物特征。银叶树通常散布于红树林内缘,以优势种成林则非常少见(简曙光等,2004;陈晓霞等,2015)。近 30年来,海岸经济快速发展导致银叶树生境的大量丧失,目前中国现存成年个体数在 20株以上的野生银叶树种群仅见于广东深圳市盐灶和海丰县香坑、广西防城港和海南清澜港(简曙光等,2004)。其中,深圳盐灶种群是目前全国乃至全世界发现分布最完整、树龄最长的银叶树群落,保有多株树龄过百的古树。由于经济发展、旅游开发等多种原因影响,该群落目前面积仅为0.8 hm2左右,并已表现出部分濒危特征,如分布区域减少、更新困难等(陈晓霞等,2015)。
现有研究对银叶树群落的群落特征(陈晓霞等,2015)、生态学特性(简曙光等,2005;孙红斌等,2018)、化学成分(Christopher et al.,2014;Ge et al.,2016)及抗性(高敏等,2015)等已有探讨,但尚未见文献对群落优势树种生长与土壤环境的关系进行研究。本文以深圳盐灶银叶树群落为研究对象,探讨优势物种的生长和更新对土壤环境与种间关系的响应特点,以期为银叶树群落的保育与管理提供科学依据。
1 研究方法
1.1 研究地点
本研究位于广东省深圳市大鹏半岛北端盐灶(22°37′—22°39′N,114°30—114°32′E),为亚热带季风气候,年均温为 22.1 ℃,年平均相对湿度为79%,降水量为1800.4 mm。该银叶树群落多样性较高,林下草本、蕨类植物和幼苗分布广泛。群落生境主要为内海湾沼泽(海生环境)、邻近的风水林山坡(陆生环境)及风水林邻海海岸(海生环境)等3种生境(简曙光等,2005)。树龄超过500 a的银叶树1棵,树龄200 a以上的约32棵,以及许多树龄过百的古树(陈晓霞等,2015)。此外,群落内还包括多种红树植物,其中真红树有桐花树(Aegiceras corniculatum)、木榄(Bruguiera gymnorhiza)、秋茄(Kandeliacandel)、卤蕨(Acrostichumaureum)、老鼠簕(Acanthus ilicifolius)、海漆(Excoecariaagallocha)和白骨壤(Avicenniamarina)等,半红树有海杧果(Cerbera manghas)、黄槿(Hibiscustiliaceus)、苦郎树(Clerodendruminerme)等。
1.2 群落样地调查
在银叶树群落中设置20个20 m×20 m样地,每个样地划分为4个10 m×10 m样方(图1)。在每个样方内,胸径大于1 cm的个体视为乔木,对乔木层采取每木调查,记录树木生长状况(胸径、树高、冠幅、分枝数)及坐标位置;设置5个5 m×5 m小样方,对灌木层进行调查,包括灌木株数、平均高、基径及冠幅。群落调查于2017年12月完成。本次调查乔木层共发现77种植物,隶属于34科61属;灌木层共发现47种植物,隶属于29科42属。各样方情况见表1。

图1 深圳盐灶银叶树群落样方示意图Fig. 1 Plots in Heritiera littoralis community in Yanzao, Shenzhen

表1 银叶树群落20个样方物种丰富度及主要优势树种重要值Table 1 Species richness and important values of major dominant species for 20 plots in Heritiera littoralis community

续表1 银叶树群落20个样方物种丰富度及主要优势树种重要值Continued table 1 Species richness and important values of major dominant species for 20 plots in Heritiera littoralis community
1.3 土壤采样及指标测定
群落调查同时,于每个样地内采集3处0-20 cm表层土样品,采集后在室内通风处风干后,除去植物根系、碎石等杂物,磨碎后过筛,测定其理化性质。土壤指标测定方法如下:土壤pH值采用土水质量比 1:2.5电位法测定;土壤有机质采用重铬酸钾外热氧化法测定;全氮采用凯氏定氮法测定;全磷采用 NaOH-熔融-钼锑抗比色法测定;全钾采用NaOH-火焰光度法测定;有效氮采用碱解扩散法测定;有效磷采用双酸浸提后钼锑抗比色法测定;有效钾采用中性乙酸铵提取后火焰光度计法测定;土壤盐度采用水提烘干法测定。具体方法见鲍士旦(2000)。20个样方土壤状况见表2。
1.4 数据统计分析
计算群落中乔木层和灌木层各物种的相对多度、相对频度和相对显著度,并计算各物种的重要值,以区分群落中的主要优势树种。计算公式如下:

表2 银叶树群落20个样方土壤状况Table 2 Soil condition of 20 plots in Heritiera littoralis community

其中,Ir为重要值,Dr为相对密度,Cr为相对显著度,Fr为相对频度;S为物种总数;Hi为对应物种i的密度,Ci为对应物种显著度,Fi为对应物种频度。
此外,计算各样地物种重要值及多样性相关指数,以衡量群落物种多样性,包括物种丰富度(S)、Shannon-Weiner指数(H′)、Simpson 指数(D)、Pielou均匀度指数(E),计算公式如下:


其中Pi为对应物种个体数占总个体数比例,S为物种数。
为研究银叶树群落组成与结构及优势树种与土壤、群落多样性的关系,分别对乔木层和灌木层采用非度量多维尺度分析(Nonmetric multidimensional scaling,NMDS)进行排序分析,然后对NMDS分析结果与土壤、群落多样性及优势树种重要值进行相关分析。对银叶树群落分析发现,乔木层主要优势树种为银叶树、阴香(Cinnamomum burmanni)、假苹婆(Sterculia lanceolata)、海杧果及多毛茜草树(Aidia pycnantha),重要值分别为0.24、0.08、0.07、0.05及0.05;灌木层主要树种为阴香、九节(Psychotria rubra)、银叶树、罗伞树(Ardisia quinquegona)及假苹婆,重要值分别为 0.12、0.12、0.12、0.10及0.07。因此,在NMDS分析中采用以上优势树种重要值进行分析。
此外,采用Pearson相关分析研究不同群落中优势树种重要值与群落多样性及土壤的关系,以探讨乔木层和灌木层的优势树种生长对群落组成及土壤状况的响应。
由于灌木层个体分布会受到乔木层个体分布影响,因此,灌木层优势树种重要值与环境的相关关系结果很可能受此影响,无法得到灌木层优势树种生长对环境的真实响应状况。因此,对乔木层和灌木层均占优的3种主要优势树种,即银叶树、阴香和假苹婆进行进一步分析:以灌木层重要值对乔木层重要值作线性相关分析,利用灌木层重要值残差计算去趋势化值(Lankau,2013),再利用该去趋势化值与多样性及土壤进行Pearson相关分析。
2 结果与分析
2.1 乔木层群落结构与优势树种分布趋势
NMDS分析表明,乔木层群落结构与土壤 pH及盐度显著相关,与土壤营养状况相关性不显著(表 3)。排序图进一步显示,土壤 pH及盐度与群落多样性呈现负相关趋势(图2),随着pH和盐度的增加,群落物种及多样性降低。多样性相关指数在排序图中指向较为一致,表明随着群落中物种多样性的增加,均匀度也随之增加,群落结构趋于稳定。阴香、假苹婆、海杧果和多毛茜草树的重要值均随着群落多样性的增加呈现不同程度的增加,而银叶树重要值则与群落多样性指数呈负相关,表明在多样性越高的群落中,银叶树重要值越低。此外,银叶树重要值与土壤pH及盐度有较一致的指向,说明随着土壤pH及盐度的增加,群落中银叶树优势程度增加。
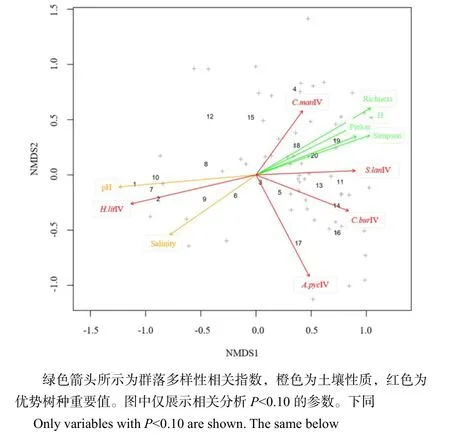
图2 乔木层NMDS排序及与群落多样性相关指数、土壤性质、优势树种重要值相关分析Fig. 2 NMDS plot and post-hoc correlation with variables describing community diversity (green arrows), soil properties (orange) and important values of dominant species (red) in tree layer
优势树种重要值与土壤、群落多样性相关指标的相关分析结果表明,除土壤pH及盐度外,各优势树种的生长和分布还与土壤营养状况尤其是磷和钾有关,但与氮元素含量相关性不显著(表4)。就树种而言,银叶树和海杧果重要值与钾含量相关,假苹婆和多毛茜草树重要值与磷含量相关,阴香重要值与所测定的土壤主要元素均不呈显著相关性。与NMDS分析结果类似,银叶树与所测定的大部分土壤营养元素含量呈负相关,其他树种则与土壤营养元素含量呈正相关。这说明在乔木层,银叶树随着土壤营养元素和群落多样性的分布趋势异于其他树种。

表3 乔木层和灌木层NMDS分析结果Table 3 Results of post hoc ordination of NMDS scores for tree and shrub layers

表4 乔木层优势树种重要值与土壤性质及群落多样性指数的相关分析结果Table 4 Correlation results of dominant species important values with soil properties and community diversity indices for tree layer
2.2 灌木层群落结构与优势树种幼树分布趋势

图3 灌木层NMDS排序及与群落多样性相关指数、土壤性质、优势树种重要值相关分析Fig. 3 NMDS plot and post-hoc correlation with variables describing community diversity, soil properties and important values of dominant species in shrub layer
灌木层群落结构与土壤 pH、全磷及有效磷含量显著相关。除阴香以外,其他优势树种重要值在群落中的变化趋势均与群落结构显著相关(表3)。排序分析进一步显示(图3),土壤pH与群落多样性呈负相关关系,磷及有效磷含量与多样性呈正相关关系。与乔木层多样性趋势不同的是,灌木层多样性相关指数变化趋势并不集中,丰富度指数与其他多样性指数在第二轴上指向不同方向。灌木层银叶树重要值变化趋势与乔木层相似,均与土壤 pH值指向相近,与群落多样性相关指数指向相反,说明灌木层银叶树的分布趋势异于其他优势树种。
灌木层优势物种重要值与土壤、群落多样性相关分析表明,除阴香以外,其他优势物种的重要值与土壤pH具有较高的相关性(表5)。银叶树重要值还与有效磷、有效钾及盐度相关,假苹婆重要值还与全钾及有效磷含量具有较高相关性。所有物种重要值与土壤氮含量相关性均不显著(表5),这与乔木层趋势类似,表明在该群落中磷和钾可能是限制物种生长和更新的主要元素。此外,除阴香以外的其他优势物种均在不同程度上与群落多样性相关,其中以银叶树相关性较高,罗伞树相关性较低。与乔木层类似,银叶树随着土壤和群落物种多样性的变化趋势与其他树种相反。

表5 灌木层优势树种重要值与土壤性质及群落多样性指数的相关分析结果Table 5 Correlation results of dominant species important values with soil properties and community diversity indices for shrub layer
为排除乔木层树种分布对灌木层物种对环境响应的潜在影响,利用银叶树、阴香和假苹婆的去趋势化数据再次进行相关分析。结果显示,去除乔木层树种分布的影响后,群落中银叶树和阴香的重要值与土壤状况相关性较低,假苹婆则显示出与各营养元素较高的相关性,表明假苹婆幼树对土壤元素状况的响应可能较其他两种优势树种幼树更为敏感。此外,去趋势化的银叶树幼树重要值与群落Simpson和Pielou多样性指数呈较显著负相关,其他两个树种重要值则没有与群落多样性指数呈现显著相关性,表明银叶树幼树的生长和分布受群落物种组成和结构的影响较大。
3 讨论
本研究探讨了深圳盐灶银叶树群落优势树种分布与土壤及群落竞争的关系。研究发现,土壤环境和群落竞争对优势树种生长和分布影响显著。土壤pH值、盐分、磷元素和钾元素含量影响群落树种生长的主要土壤因子;乔木层优势树种的分布和生长与群落多样性格局的相关性显著,且乔木层的分布格局对幼树分布和生长产生较大影响;不同优势树种对多样性与土壤的响应差异明显,银叶树的响应趋势与其他优势树种相反;银叶树幼树生长显著受群落多样性的影响。
银叶树群落多样性表现为从邻海海岸向内海沼泽递减(图2和图3),该结果与以往研究结论一致(陈晓霞等,2015)。该多样性趋势可能与土壤养分逐渐减少有关,邻海生境主要为沙壤,积累了大量有机质,山坡生境则土层较深,凋落物输入较多,这两类生境充足的养分和适宜的土壤化学性质为树种生长提供了良好条件;相反,沼泽生境中土壤缺氧,有机物分解缓慢,养分含量相对较低。此外,由于海水涨退潮,沼泽生境土壤高度盐渍化,盐分和pH值较高,可能不利于物种建群繁殖。以往研究也发现,土壤盐分和pH值是群落物种分布的关键环境因子,过高的盐分和pH值可能会降低群落物种多样性(马宗文等,2012;尹德洁等,2018)。
除盐分及pH值的影响外,银叶树群落物种生长与磷元素的相关性较高。磷元素是热带亚热带地区森林的主要限制元素(Wardle et al.,2004;Vitousek et al.,2010),以往研究发现,该银叶树群落土壤磷含量较低(孙红斌等,2018),表明该群落生长可能受到一定程度的磷限制。此外,乔木层和灌木层的结果均显示物种分布与氮元素相关关系不显著,充足的氮供给可能进一步加剧磷元素对该群落物种生长的限制。
不同优势树种对群落竞争和土壤因子的响应存在差异。在乔木层,各优势树种均表现出与土壤因子和多样性指数的显著相关性;在灌木层,除阴香外的其他树种与土壤和多样性指数显著相关。具体而言,银叶树幼树的分布受群落竞争格局影响,当群落多样性和均匀度增加时,其幼树的重要值下降,这表明银叶树幼树的种间竞争力可能较其他物种弱;在排除乔木层的潜在影响后,3种优势树种中只有银叶树表现出与群落多样性指数的显著负相关关系,结合银叶树乔木重要值最高、幼树重要值低于阴香和九节的调查结果可推测,相比于土壤因素,银叶树幼树的生长可能更明显地受到群落物种竞争的影响。阴香幼树重要值的变化与群落多样性及土壤因子均无显著相关性,且阴香乔木和幼树的重要值在群落中均较高,表明阴香可能具有较高的环境适应性。假苹婆乔木和幼树的重要值均与磷元素有较高的相关性,表明磷对假苹婆生长可能有着重要作用。
银叶树随着多样性和土壤的分布格局不同于其他优势树种。首先,银叶树重要值随着群落多样性的增加而减少,其个体多分布在沼泽生境,这一方面有可能是其他树种对低养分的盐渍化土壤无法适应,导致其无法在此区域建群,为银叶树提供了低种间竞争的生长空间;另外,由于部分古树在该生境发现,因此不排除这些银叶树是人为栽种而非自然扩散至该生境的可能性。但由于银叶树不是典型红树植物,不具备高耐盐性,因此推测该生境的土壤状况可能并非最适条件,银叶树在此生境优势群落的形成很可能是对土壤和群落竞争权衡的结果。其次,银叶树幼树的分布格局反映了群落物种竞争对银叶树更新的影响。乔木层和灌木层的银叶树个体分布趋势均表现出对群落竞争的规避,即在低群落多样性的沼泽区域分布个体更多,这表明,尽管银叶树在群落中重要值较高,但其本身可能并非高竞争力物种,从长期来看,在土壤条件较好的山坡及邻海海岸生境中,与其他树种的竞争可能会对银叶树幼树的生长产生不利影响,进而影响银叶树种群在群落中的更新和维持。
4 结论
深圳盐灶银叶树群落优势树种的生长受到土壤环境和群落物种竞争的影响。土壤pH值、盐分、磷元素和钾元素含量影响群落树种生长的主要土壤因子,乔木层优势树种的分布与群落多样性显著相关,且乔木层的分布格局对幼树分布存在较大影响。不同优势树种的生长响应差异明显,随着群落多样性的增加和土壤盐分及pH值的降低,除银叶树外的其他优势树种重要值均呈上升趋势,银叶树重要值则呈下降趋势。银叶树幼树生长显著受群落多样性的影响。
综上所述,本研究推测,对于群落中主要优势树种,如阴香、假苹婆、海杧果、多毛茜草树、九节和罗伞树等,土壤养分尤其是磷和钾元素含量的提高及群落正向演替可能促进这些树种的生长更新。银叶树则更多地分布于沼泽等土壤环境较差的生境中,其幼树对群落竞争较为敏感,并很可能影响其在山坡、邻海海岸等土壤环境较好的生境中的更新和维持。因此,对银叶树种群的保育措施应考虑改善沼泽土壤环境,以缓解土壤pH值和盐分过高对银叶树的潜在胁迫,同时应持续监测银叶树幼苗幼树个体在山坡和邻海海岸生境中的生长状况,以跟踪银叶树在这些生境中的种群动态变化。此外,本研究仅利用调查手段分析了银叶树群落主要优势树种的分布格局,今后需结合控制实验手段研究银叶树对竞争和土壤环境的响应,为银叶树种群的保育措施提供更具体的科学指导。
猜你喜欢
建筑与预算(2022年6期)2022-07-08 13:59:18
河北果树(2020年1期)2020-02-09 12:31:18
资源节约与环保(2018年1期)2018-02-08 02:18:33
资源节约与环保(2018年1期)2018-02-08 02:17:49
中国资源综合利用(2017年3期)2018-01-22 02:45:52
现代园艺(2017年11期)2017-06-28 11:32:57
土壤与作物(2015年3期)2015-12-08 00:46:58
中国质量与标准导报(2015年2期)2015-02-28 22:27:14
建筑与预算(2014年5期)2014-04-11 07:29:28
河南科技(2014年16期)2014-02-27 14:13:33
