
贵州分布长肢林蛙种组物种一新记录
——徂徕林蛙
2019-11-22 07:07肖宁罗庆华罗涛魏芳曾亚军
四川动物 2019年6期
肖宁,罗庆华,罗涛,魏芳,曾亚军
(1.贵阳护理职业学院,贵阳550081;2.贵州师范大学生命科学学院,贵阳550001; 3.荔波县世界自然遗产管理局,贵州荔波558400;4.贵州省林业科学研究院,贵阳550004)
徂徕林蛙Ranaculaiensis隶属于蛙科Ranidae蛙属Rana,是Li等(2008)依据山东泰安徂徕山的标本而描述,目前已知分布于山东、河南、湖南和广西等地(费梁等,2012;AmphibiaWeb,2019;Frost,2019;中国两栖类,2019)。杨宝田等(2013)依据线粒体16S rRNA支持徂徕林蛙的有效性,并认为其与镇海林蛙Ranazhenhaiensis为姐妹种,两者处于物种分化过程中的并系阶段,其物种分化还未完成。大多数研究均支持该物种的有效性(Yanetal.,2011;杨宝田等,2013;Yuanetal.,2016;Wangetal.,2017;Zhaoetal.,2017)。贵州省现记录分布的蛙属物种有威宁蛙R.weiningensis、昭觉林蛙R.chaochiaoensis、峨眉林蛙R.omeimontis、寒露林蛙R.hanluica和镇海林蛙R.zhenhaiensis5种(伍律等,1986;费梁等,2009;张雷等,2010;陈继军等,2017)。魏刚等(2017)基于分子形态数据将贵州梵净山地区的林蛙种类记录为镇海林蛙(样本号:lshz40)和寒露林蛙(样本号:lshz41),但是梵净山种群(样本号:lshz40)与徂徕林蛙模式产地种群(山东徂徕,标本号:SYNU07050129,GenBank登录号:KF204620)的遗传距离为0。
2018年5月在贵州省黄平县朱家山县级自然保护区进行本底资源调查时采集到林蛙标本10号,经过形态学比较和分子系统发育分析,鉴定为徂徕林蛙,为贵州省新记录。本研究对徂徕林蛙贵州种群进行了详细的形态学描述,补充了其分子遗传学数据,并与长肢林蛙R.longicrus种组物种进行了系统发生关系分析,探讨它们之间的遗传分化和系统演化关系。
1 材料与方法
1.1 材料
徂徕林蛙成体标本10号(4雄6雌),采自贵州省黄平县朱家山县级自然保护区(107°39′44.49″E,26°54′35.23″N,海拔833 m;图1)。取腿部肌肉保存于90%乙醇中;使用10%福尔马林浸泡标本,保存于贵州师范大学生命科学学院动物生态学标本室(GZNU)。

图1 徂徕林蛙采样点Fig.1 Sampling points of Rana culaiensis
1.2 形态学数据测量
测量标准依据费梁等(2009),使用游标卡尺(Serial No.14133868,德国)测量标本的12个形态特征(精确度为0.01 mm),包括头体长(SVL)、头长(HL)、头宽(HW)、吻长(SL)、眼径(ED)、鼻间距(INS)、眼间距(IOS)、前臂及手长(LAHL)、手长(HL)、后肢全长(HLL)、胫长(TL)和足长(FL)。
1.3 DNA提取、扩增和测序
总DNA提取自保存于90%乙醇中的标本腿部肌肉,使用DNeasy Blood &Tissue Kit(QIAGEN Catalog 69504)进行提取。选择线粒体基因16SrRNA、cytb和ND2作为分子标记,扩增引物和反应体系参考Yuan等(2016)进行PCR扩增。扩增产物由成都擎科梓熙生物技术有限公司双向测序。
1.4 系统发育分析
选择长肢林蛙种组9个物种的16SrRNA、cytb和ND2基因序列用于系统发育分析(表1),以威宁蛙、中国林蛙R.chensinensis为外群,分子数据主要来自于Yuan等(2016)。使用MEGA 7.0(Kumaretal.,2016)中MUSCLE进行多重序列比对,并使用DAMBE(Xia &Xie,2001)进行序列饱和性检测。
使用贝叶斯(BI)法和最大似然(ML)法进行系统发育树的构建。最适模型基于赤池信息准则(AIC)在PartitionFinder 2(Lanfearetal.,2012)中进行选择。在MrBayes 3.2(Ronquistetal.,2012)中进行BI分析,设置4条链独立运行107代,每隔100代对树进行1次采样,前25%被舍弃,使用贝叶斯后验概率(BPP)来检测节点的支持率。在Tracer 1.6(Rambautetal.,2013)中查看相关运行参数的收敛性和有效样本量(ESS)。在IQ-TREE(Nguyenetal.,2014)中运行1 000次Bootstrap重复抽样分析,以检验各节点置信度,使用自举支持值(BSP)检验节点处的支持率。在MEGA 7.0中以物种为组,采用分组的方法计算了长肢林蛙种组物种间的遗传距离。
2 结果
2.1 系统发育分析
将采自贵州省黄平县朱家山县级自然保护区的4号林蛙标本的16SrRNA、cytb和ND2基因序列上传到GenBank,标本编号对应的检索号见表1。对长肢林蛙种组9个物种共计26个标本的16SrRNA、cytb和ND2基因序列进行多重序列比对拼接后的碱基长度为2 336 bp。基于使用3个线粒体基因联合序列计算的长肢林蛙种组物种间的遗传距离为0.77%~2.58%;采自朱家山县级自然保护区的徂徕林蛙与该物种模式产地种群之间的遗传距离为0.77%。
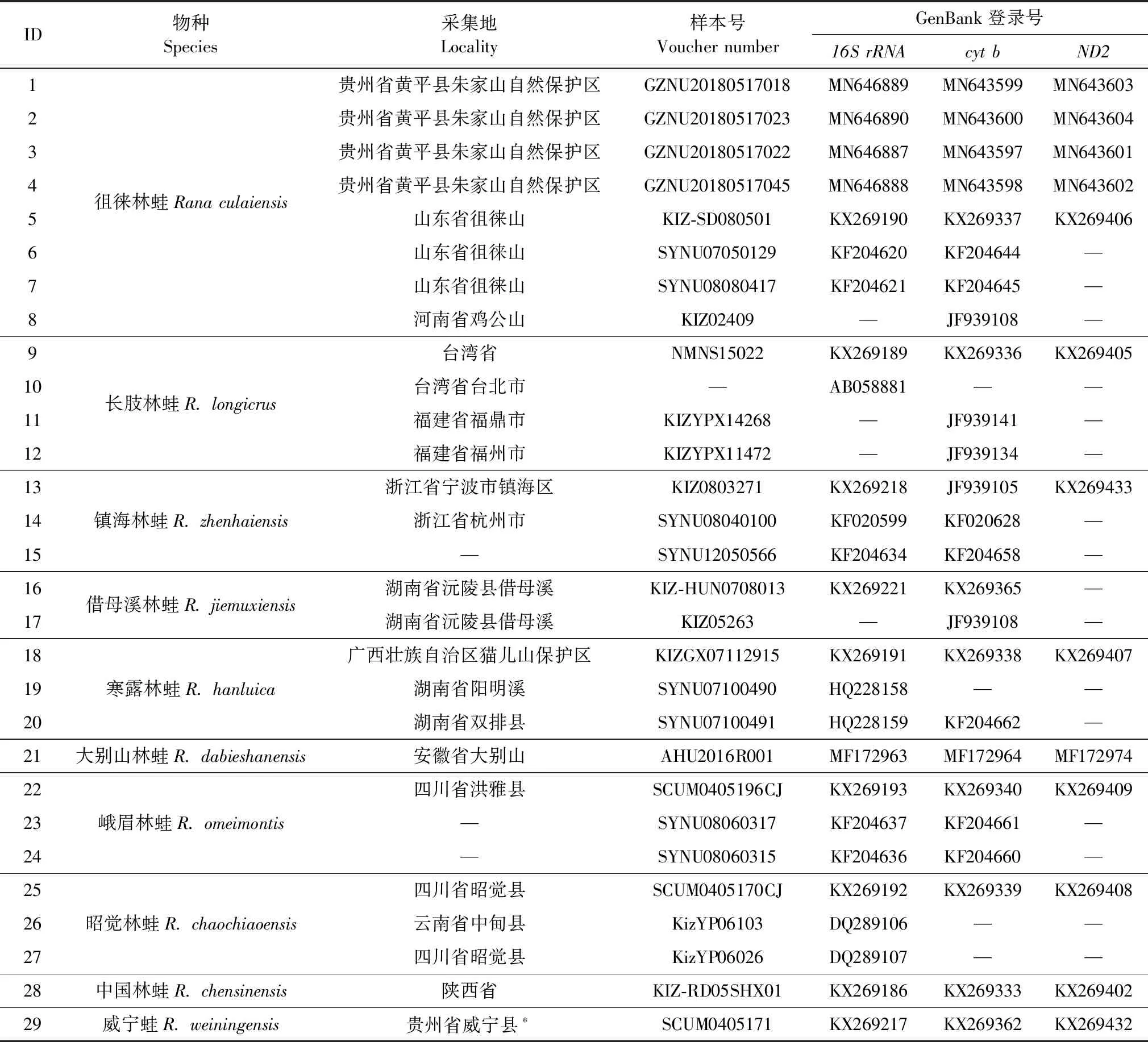
表1 由GenBank中下载用于构建系统发育树的相关序列Table 1 Information of gene sequences used for building phylogenetic trees
注:*Yuan等(2016)将此标本错录入为四川威宁,现修订为贵州威宁
Note:*Yuanetal.(2016)recorded "Sichuan:Weining" as the sampling site,it is now revised as "Guizhou:Weining"
基于BI法和ML法构建的系统发育树生成了相同的的拓扑结构(图2),仅在末端分支节点上存在一些差异。本研究样本与模式产地的徂徕林蛙聚为一支(BPP=0.99,BSP=91),与镇海林蛙为姐妹种,这与之前的研究有着相似的树形结构(Yanetal.,2011;杨宝田等,2013;Yuanetal.,2016;Wangetal.,2017)。因此,基于系统发育分析和遗传距离的结果表明,本次采集的林蛙物种为徂徕林蛙。
2.2 形态描述
雄性头体长41.61~51.09 mm,雌性头体长48.46~56.60 mm(表2);头长大于头宽(雄),头长略小于头宽(雌);吻端略钝圆,突出于下唇;吻棱清晰;鼻孔位于吻棱下方,在眼和吻端之间;眼间距略等于鼻间距;瞳孔呈横椭圆形;颞褶弱;鼓膜近圆形,约为眼径的2/3;犁骨齿列呈两短斜行;舌卵圆形,其后端缺刻深。
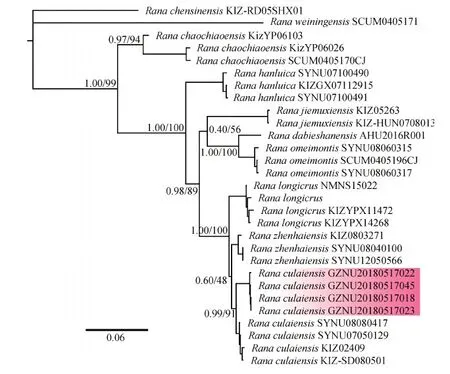
图2 基于线粒体联合序列构建的长肢林蛙种组物种系统发育树Fig.2 Phylogenetic tree of the Rana longicrus species group constructed using mitochondrial joint sequence红色部分表示来自贵州省黄平县的徂徕林蛙样品Red indicates the R.culaiensis samples were from Huangping county,Guizhou province
四肢较为细长,前臂及手长为头体长的59.29%;指细长,末端钝圆;指侧具较细的缘膜;第二指与第四指约等长;指长顺序为Ⅲ>Ⅱ≈Ⅳ>Ⅰ;关节下瘤发达,内掌突较大呈卵圆形;后肢全长为体全长的189.76%;前伸贴体时胫跗关节超过眼前角;左右跟部略重叠;胫长为头体长的60.32%;胫长略大于足长;趾细长,趾端略膨大呈圆形而无沟;关节下瘤大而明显;第三趾长于第五趾,第三趾超过第四趾第二关节下瘤,第五趾达第二关节下瘤,趾长顺序为Ⅳ>Ⅲ>Ⅴ>Ⅱ>Ⅰ;趾间蹼较发达;雌蛙趾间蹼均不达趾端,第四趾两侧蹼其凹陷处位于第四趾第一关节与第二关节约 1/2处,两侧蹼以缘膜达第二关节下瘤;内蹠突呈椭圆形、明显,外蹠突不显著(图3)。
生活史颜色变异较大,为棕色或橄榄棕色,雌蛙多棕色;皮肤较光滑,背部及体侧有少数小圆疣;体腹面光滑;体背面多为红褐色或棕灰色而无深色斑,背侧褶细窄从眼后上方直达胯部,在颞部上方略向外侧弯曲,不与颞褶相连;颔腺细窄从口角延续至肩部上方;颞部有黑色三角斑,眼间有深色横斑或横斑颜色浅;两眼间的横斑、颞部的三角形斑以及背部的“八”形斑或短杆状斑均为黑褐色,个别个体体侧有褐色斑或小点;四肢背面黑褐色横纹,较为细窄长,排列较为紧密,前臂有2~3条,股部有4~7条;胫部有5~6条;腹面乳黄色(图3)。
第二性征:雄性无声囊;雄性第一指具大而密的婚刺。
2.3 生境描述
贵州朱家山县级自然保护区位于107°35′~107°42′E,26°53′~27°06′N,为武陵山脉向西南的延伸部分,是舞阳河主流和清水江支流的主要发源地。该区具明显的亚热带季风气候特征。采自朱家山县级自然保护区的徂徕林蛙生境较多样,包括农田和山间较湿润的林区。5月调查时已发现其幼体完全变态营陆地生活,因此推测该蛙的繁殖时间为3—4月,这与Li等(2008)记录的一致。
3 讨论
本研究结合形态学数据和3个线粒体基因从分子生物学的角度证实贵州省黄平县朱家山县级自然保护区分布林蛙为徂徕林蛙。从贵州黄平和山东徂徕山采集的徂徕林蛙在繁殖时间(Lietal.,2008)和遗传上的差异较小(表3),两地种群遗传距离为0.77%,小于徂徕林蛙与镇海林蛙之间的遗传距离(1.80%),因此本研究采纳了杨宝田等(2013)的观点,将贵州黄平分布的林蛙记录为徂徕林蛙。但贵州黄平和山东徂徕山的样本在身体各部位的形态性状和量度上仍存在一定的差异:1)前者的第四趾两侧蹼凹陷处位于第四趾第一关节与第二关约1/2处;而后者的趾间蹼缺刻深,其凹陷处位于第四趾第二关节处;2)前者眼间有深色横斑或色斑颜色较浅,背面有倒“V”形斑;而后者眼间无深色横斑,无倒“V”形斑;3)在前臂及手长占头体长的比例上,前者(雌性为64.07%,雄性为59.29%)大于后者(雌性为45.54%,雄性为44.26%);4)在体型上,前者(平均值:雄性44.73 mm,雌性52.29 mm)小于后者(平均值:雄性53.57 mm,雌性62.04 mm)。由于本研究采集的徂徕林蛙标本数量有限,上述差异是否为适应不同的地理环境而导致的形态变异,还有待采集符合统计学的标本数据来论证。
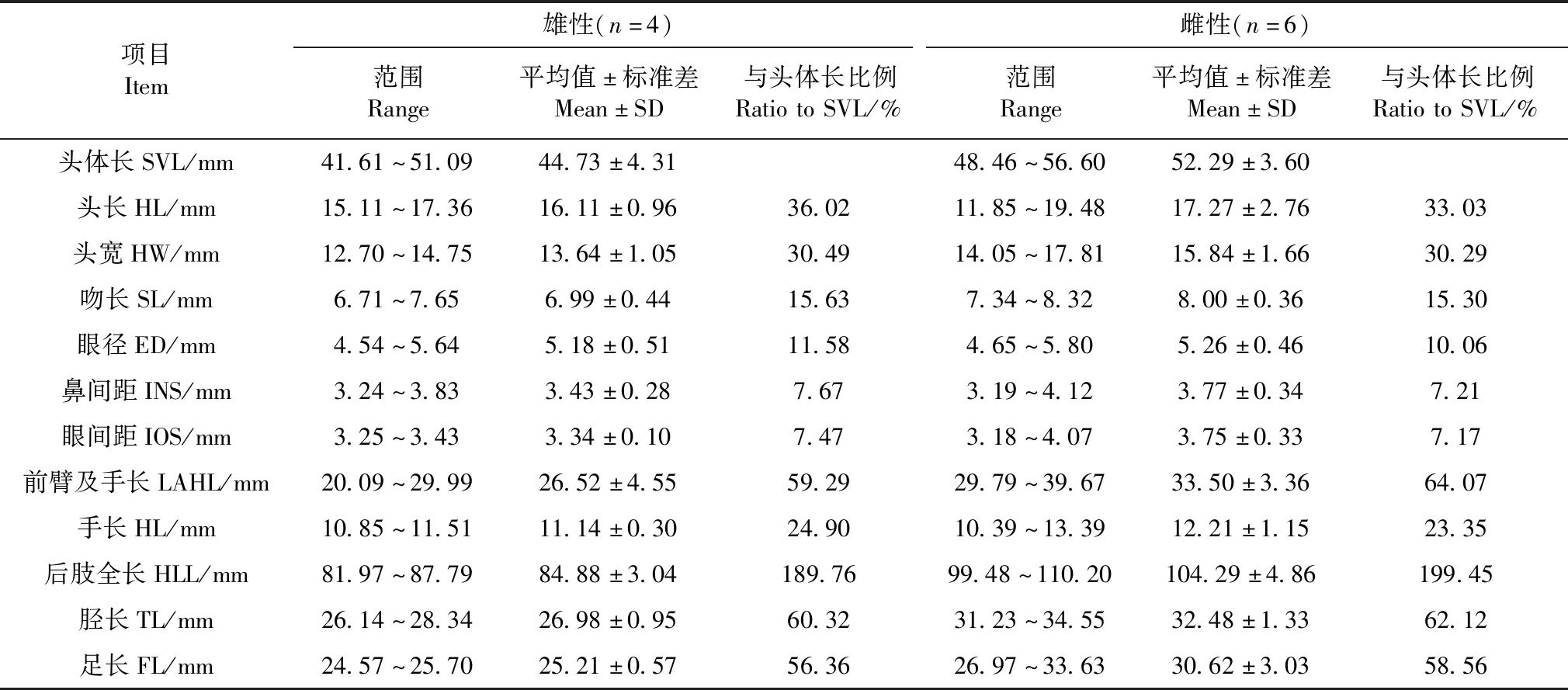
表2 贵州黄平朱家山县级自然保护区徂徕林蛙的量度Table 2 Morphological measure of Rana culaiensis in the Zhujiashan Nature Reserve,Huangping,Guizhou

图3 贵州省黄平县朱家山县级自然保护区分布的徂徕林蛙生态照Fig.3 Ecological images of Rana culaiensis distributed in the Zhujiashan Nature Reserve,Huangping county,Guizhou provinceA.背面观,B.腹面观,C.侧面观,D.手腹面观,E.足腹面观,F.头部侧面观A.dorsal view,B.ventral view,C.side view,D.ventral view of hand,E.ventral view of foot,F.side view of head

表3 基于线粒体联合序列计算的长肢林蛙种组物种间的遗传距离(P-distance,%)Table 3 Genetic distances among the species of Rana longicrus species group based on mitochondrial joint sequences (P-distance,%)
贵州地区最早分布的林蛙记录为日本林蛙指名亚种R.japonicajaponica和日本林蛙昭觉亚种R.japonicachaochiaoensis,后修订为峨眉林蛙和昭觉林蛙(Liuetal.,1946;费梁等,1990;叶昌媛等,1993)。
这2种林蛙曾被作为广泛种记录在贵州省的各个区县(伍律等,1986;贵州省黔东南苗族侗族自治州人民政府,1989;喻理飞,2004;贵州省林业厅,2006;费梁等,2009;贵州省林业科学研究院,贵州省盘县林业局,2013;苟光前等,2017)。但近年来贵州省东部地区的蛙属物种分布尚存争议(张雷等,2010;陈继军等,2017)。魏刚等(2017)对梵净山地区的蛙属进行了调查和研究,基于线粒体16SrRNA的分析表明,该地区分布有镇海林蛙和寒露林蛙,而未发现所记录的日本林蛙指名亚种(贵州省林业厅,梵净山国家级自然保护区,1990),即峨眉林蛙。然而,在魏刚等(2017)基于线粒体16SrRNA的研究中,其镇海林蛙样本(样本号:lshz40)与来自模式产地的徂徕林蛙的遗传距离为0,加上黄平朱家山与梵净山地区同属于武陵山系,对物种的阻隔较小(Zhanetal.,2009),因此,魏刚等(2017)记录于梵净山地区分布的镇海林蛙与朱家山地区分布的林蛙应为同一物种,是徂徕林蛙的错误鉴定。截止目前,贵州地区是否分布有峨眉林蛙,尚有待进一步调查。
蛙属物种的形态保守性成为依靠形态特征鉴别的困难,因此在今后有必要结合分子生物学开展系统地理学和分类学研究。贵州作为第一阶梯和第二阶梯的过渡区域,已有越来越多的研究证明了该地区是一些生物的起源和扩散的中心(Yanetal.,2013;Yuanetal.,2016;姚明灿等,2018)。因此,对于在青藏高原以外的地区进行相关的生物地理学研究和厘清贵州高原地区的物种组成,以期进一步理解高原运动对物种形成和对保护物种多样性作出科学的保护决策有着重要的作用。
致谢:感谢贵州省林业科学研究院、黄平县朱家山县级自然保护区工作人员在调查过程中提供的帮助。
猜你喜欢
Chinese Physics B(2022年11期)2022-11-21
Plasma Science and Technology(2022年2期)2022-03-10
妇女(2021年12期)2021-12-25
吉林蔬菜(2020年1期)2020-04-21
红蜻蜓·低年级(2017年11期)2018-03-09
现代营销·经营版(2016年12期)2016-12-21
浙江林业(2016年6期)2016-07-05
绿色中国(2015年12期)2016-01-05
阅读与作文(英语初中版)(2013年3期)2013-04-10
三月三(2009年10期)2009-10-07
