
麋鹿的分类地位与遗传多样性研究概述
2019-11-19 08:26:14张树苗白加德李夷平梁兵宽李俊芳李俊生
野生动物学报 2019年4期
张树苗 白加德 李夷平 陈 颀 梁兵宽 李俊芳 李俊生 常 江*
(1.北京麋鹿生态实验中心,北京,100076;2.国家林业和草原局调查规划设计院,北京,100714;3.国家环境保护区域生态过程与功能评估重点实验室,中国环境科学研究院,北京,100012)
麋鹿(Elaphurusdavidianus)是世界自然保护联盟(IUCN)红皮书野外灭绝物种,是我国I级重点保护野生动物,是国内外物种迁地保护的经典案例,是世界物种重引进项目的成功范例。野生麋鹿种群在经历商周的鼎盛时期之后走向衰落,至十九世纪末,麋鹿在我国本土灭绝[1]。1893—1895年,英国第十一世贝福特公爵收集了18只散落在欧洲各地的麋鹿,集中饲养在其乌邦寺庄园里,这些麋鹿成了全球现存麋鹿的祖先“奠基者”[2]。第一次世界大战后,乌邦寺的麋鹿种群日渐扩大。为挽救濒危物种,保护生态平衡,实现麋鹿回归故乡,中国于1985—1987年共从英国引入麋鹿77只,分别饲养于北京南海子麋鹿苑和江苏大丰麋鹿国家级自然保护区,建立了麋鹿半散放种群,拉开了麋鹿在我国迁地保护的序幕[3]。经过30多年的努力,目前,中国麋鹿种群数量已超过7 000只,麋鹿栖息地较为稳定,暂未发现有明显的生存力和繁殖力下降的情况,是中国乃至全球珍稀物种保护领域中的成功范例。但是,麋鹿在近代经历了4次遗传瓶颈事件,使其群体遗传变异严重丢失。第一次是英国乌邦寺建群时,第二次是英国各动物园建群时,第三次是北京南海子麋鹿苑和江苏大丰麋鹿国家级自然保护区引入建群时,第四次是从南海子和大丰引入到湖北天鹅洲建群时[4]。目前,基于分子生物学的研究显示,麋鹿中性分子标记遗传多样性较低,且高度近亲繁殖,这个遗传困境备受学术界、野生动物保护工作者与管理部门的关注与重视。虽然我国的麋鹿种群发展稳定,且还会继续增长,在保护生物学领域创造了物种重引进的奇迹。但不能忽视,麋鹿的近交系数极高,奠基者数量较少,仍然面临严峻的遗传困境,在种群管理方面需要避免高度近交和遗传多样性减少与丧失。Clutton-Brock等[5]指出大型草食动物的种群管理应考虑实施人工种群调控措施,Foose和 Frankham等[6-7]指出濒危物种的保育要实施遗传管理措施。本文介绍麋鹿分类地位和麋鹿遗传多样性的有关研究,为中国麋鹿遗传资源保护及种群的健康可持续发展和管理提供参考。
1 麋鹿分类地位
麋鹿是中国特有的动物种类,麋鹿属曾被认为有5种,其中双叉麋鹿(E.bifurcatus)、晋南麋鹿(E.chinanensis)、蓝田麋鹿(E.lantianensis)和台湾麋鹿(E.formosanus)4种已灭绝,现仅存达氏麋鹿(E.davidianus),也即大卫鹿[1,8-9]。
脊索动物门Chordata
脊椎动物亚门Vertebrata
哺乳纲Mammalia
偶蹄目Artiodactyla
反刍亚目Ruminantia
鹿上科Cervoidea
鹿科Cervidae
鹿亚科Cervinae
麋鹿属
Elaphurus
达氏种
E
.
davidianus
双叉种
E
.
bifurcatus
晋南种
E
.
chinanensis
蓝田种
E
.
lantianensis
台湾种
E
.
formosanus
图1麋鹿的分类地位(引自白加德,2014)[8]
Fig.1 The classification of Père David’s deer(Cited from Bai Jiade,2014)[8]
达氏种是1866年由爱德华(Milne-Edwards A.)根据法国传教士大卫从北京南苑带到法国巴黎博物馆的标本研究建立的。其属名Elaphurus为长尾之意,指出麋鹿尾长这一鲜明特征。种名“davidianus”是纪念第一个发现者大卫的,所以西方人习惯地称麋鹿为“大卫鹿”(David’s deer)。在中国,很少这样称呼。麋鹿是中国学术界沿用甲骨文的称谓,老百姓称麋鹿为“四不像”,近三四十年来,麋鹿一词已被普及[9]。
双叉种(E.bifurcatus)是1930年,德日进和皮韦托[10]在研究中国河北泥河湾拉方期哺乳动物群时,发现一类鹿角化石,分叉性质与现生麋鹿相类似,建立的一个新种。
1933年,Sowerby[11]根据河南安阳殷墟的发现,在真鹿属(Cervus)的名下记述了一个新种,称为真鹿属梅氏种(Cervus(Rucervus)menziesianus),后被修改划入麋鹿属中,叫麋鹿属梅氏种(E.menziesianus)[10],与现生种麋鹿相并列。1955年杨钟健[12]、1956年裴文中[13]指出梅氏种是达氏种的同物异名,梅氏种不能成立。于是,根据国际动物命名法规优先权法则,梅氏种已经统一于1866年爱德华对现生种的命名。
晋南种(E.chinanensis),是1974年贾兰坡根据山西早更新世的考古材料确定的古麋鹿新种[14]。
蓝田种(E.lantianensis),是1975年计宏祥根据陕西早更新世的材料确定的另一古麋鹿新种[15]。
1978年,日本人大裕之等修定台湾早更新世的狍属台湾种(Capreolusformosanus)为麋鹿属台湾种(E.formosanus)。
2018年,Dong等[16]对产自山西天镇的鹿角标本进行了详细研究,发现了麋鹿的两个种,其中一个是双叉麋鹿,另外一个由于形态的不同,建立了麋鹿种之下的一个亚种——原达氏鹿Elaphurusdavidianuspredavidianus。他们认为鹿科麋鹿属历史曾共有4个种8个亚种,其中达氏麋鹿有4个亚种,双叉种有2个亚种,晋南种有2个亚种,丰富了我们对麋鹿属物种多样性的认识。
关于现存达氏麋鹿的分类地位,学术界存在不同意见。盛和林[17]和Simpson[18]认为,麋鹿属于鹿科(Cervidae)鹿亚科(Cervinae)麋鹿属(Elaphurus),与Grubb,Geist和白加德等认为麋鹿是独立的属观点相一致[8,19-20]。

反刍亚目Ruminantia 鼷鹿上科Tragulodea 鼷鹿科Tragulidae 鹿上科Cervoidea 麝科Moschidae 麝属Moschus 鹿科Cervidae 獐亚科Hydropotinae 獐属Hydropotes 鹿亚科Cervinae 麂族Muntiacini 毛冠鹿属Elaphodus 麂属Muntiacus 鹿族Cervini 斑鹿属Axis 鹿属Cervus 麋鹿属Elaphurus反刍亚目Ruminantia 鼷鹿次目Tragulina 异鼷鹿科Hypertragulidae 鼷鹿科Tragulidae 有角次目Pecora 鹿科Cervidae 麝亚科Moschinae 麝属Moschus 麂亚科Muntiacinae 麂属Muntiacus 毛冠鹿属Elaphodus 鹿亚科Cervinae 斑鹿属Axis 黇鹿属Dama 鹿属Cervus 麋鹿属Elaphurus
图2反刍亚目下的分类系统(引自盛和林,1992;Simpson,1945)[17-18]
Fig.2 The classification of Ruminantia(Cited from Sheng Helin,1992;Simpson,1945)[17-18]
但有学者认为麋鹿应该并入鹿属(Cervus)。Randi等[21]基于线粒体D-loop全序列分析,探讨了鹿科、鹿亚科25个种和亚种的系统发生关系,首次提出将麋鹿并入鹿属。Gilbert等[22]认为鹿亚科分为4个属,鹿属、斑鹿属(Axis) 、黇鹿属(Dama)和沼鹿属(Rucervus),麋鹿归入鹿属,Cronin等[23-24]和Randi等[25]把鹿亚科分为 3 个属,认为麋鹿应划入鹿属。刘向华等[26]根据线粒体细胞色素b基因(Cytb)序列及部分片段构建的系统发育树探讨鹿亚科属间属内进化关系,分析结果也认为麋鹿应并入鹿属,涂剑锋等[27]研究鹿亚科动物线粒体DNA控制区全序列,构建的系统发育树结果表明,麋鹿属的麋鹿、斑鹿属的豚鹿(Axisporcinus)以及黇鹿属的黇鹿(Damadama)与鹿属的分化处于属间差异,支持将其并入鹿属,匡叶叶[28]在“建立AFLP分子标记检测体系”的基础上,通过对北京南海子245只麋鹿的AFLP检测,基于遗传相似系数矩阵,结果表明,麋鹿应属于鹿属、Emerson[29]也认为麋鹿与鹿属进化关系较近,应并入鹿属。
2 麋鹿与鹿亚科动物间的进化关系
物种间的进化关系反映了物种的系统演变历程及亲缘关系,对物种的保护遗传学研究有重要的意义。鹿亚科是大、中型的鹿类,按照盛和林[17]对鹿亚科的分类方式,鹿亚科分成:黇鹿属(Dama)、斑鹿属(Axis)、麋鹿属(Elaphurus)和鹿属(Cervus)。中国的鹿亚科动物包括:鹿属的梅花鹿(Cervusnippon)、马鹿(C.elaphus)、坡鹿(C.eldii)、水鹿(C.unicolor)和白唇鹿(C.albirostris),斑鹿属的豚鹿和麋鹿属的麋鹿(Elaphurusdavidianus)7个物种,其中麋鹿与白唇鹿是我国的特有物种(表1)。古生物化石研究表明,自中新世早期鹿亚科从鹿科总进化枝分化出来后,麋鹿属是最早从鹿亚科分离出来的进化枝[30],现存麋鹿是这一属中高度特化的物种。
表1 中国鹿亚科动物起源、保护等级及国内分布
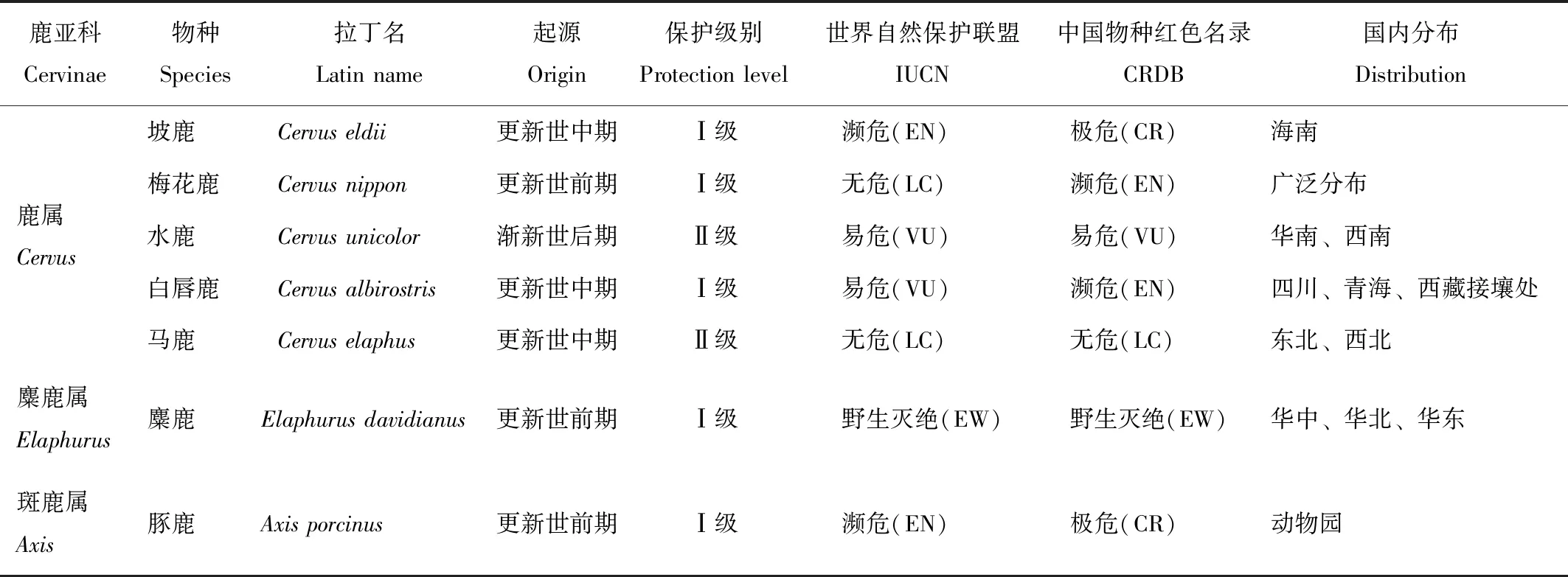
Tab.1 The origin,protection level and distribution of Cervinae animals in China
关于麋鹿与鹿亚科其他鹿的进化关系,学术界有多派观点。较多数学者认为,相比鹿亚科其他属,麋鹿属与鹿属有较近的进化关系。Grove等[31]对鹿科动物的共源性状进行了研究,结果显示麋鹿属和鹿属之间有较近的进化关系,这与对鹿科动物染色体核型研究的结果一致;国外学者利用分子生物学分析揭示,麋鹿更接近于鹿属Cervus[32-34]。Emerson和Tate[29,32]对鹿亚科4个属10个种和亚种的进化关系进行了蛋白电泳分析,发现麋鹿、马鹿和梅花鹿的遗传关系较近,结果亦支持这一观点,并认为麋鹿和其他鹿类角的形态差异并非长时间进化分歧造成的。
部分学者认为麋鹿与马鹿进化关系近。Maqbool[35]等研究了麋鹿和马鹿杂交后代的遗传连锁图谱以及其子代的数量性状座位(QTL),从遗传学角度反映二者具有较近的进化关系。
也有学者认为麋鹿与坡鹿、泽鹿(Rucervusduvaucelii)进化关系近。刘向华等[26]对鹿亚科 4 个属的马鹿东北亚种(C.elaphusxanthopygus)、甘肃亚种(C.elaphuskansuensis)、阿尔泰亚种(C.elaphusasiaticus)、西藏亚种(C.elaphuswallichii)、四川亚种(C.elaphusmacneilli)、天山亚种(C.elaphussongaricus)、四川梅花鹿(C.nipponsichuanicus)、白唇鹿、水鹿、坡鹿、豚鹿、印度斑鹿(Axisaxis)、麋鹿及鼷鹿科(Tragulidae)大鼷鹿(Tragulusnapu)共 15 个种和亚种的Cytb基因部分序列做了分析比较,分析结果认为麋鹿与坡鹿形成姊妹关系(PP=97;BP=89),具有较近的亲缘关系;盛桂莲等[36]对麋鹿、黇鹿、豚鹿、马鹿、水鹿、坡鹿、白唇鹿、梅花鹿、大鼷鹿的Cytb部分同源序列进行的系统发育分析来看,坡鹿与麋鹿聚成姊妹群,反映了二者具有较近的进化关系,这与Emerson基于蛋白质的研究结论一致。孟浩等[37]对鹿亚科的麋鹿、马鹿、坡鹿、泽鹿的mtDNA控制区序列进行比较,构建NJ分子系统发育树,发现麋鹿与坡鹿、泽鹿同属于一个分支,亲缘关系最近;涂剑锋等[27]对13种鹿亚科动物东北马鹿、甘肃马鹿、蒙古马鹿(C.elaphusalashanicus)、阿尔泰马鹿、天山马鹿、四川马鹿、塔里木马鹿(C.elaphusyarkandensis)、欧洲马鹿、东北梅花鹿(C.nipponhortulorum)、台湾梅花鹿(C.nippontaiouanus)、越南梅花鹿(C.nipponpseudaxis)、海南水鹿(C.unicolorhainana)、印度水鹿(C.unicolorniger)、台湾水鹿(C.unicolorswinhoii)、海南坡鹿(C.eldiihainanus)、泰国坡鹿(C.eldiisiamensis)、印度坡鹿(C.eldiieldii)、白唇鹿、麋鹿、豚鹿、黇鹿、毛冠鹿(Elaphoduscephalophus)的D-loop区全序列,并结合GenBank 检索到的12种鹿亚科动物同源序列开展分析,构建的系统发育树,表明麋鹿属的麋鹿、斑鹿属的豚鹿以及黇鹿属的黇鹿与鹿属的分化处于属间差异,麋鹿与鹿属的坡鹿互为姐妹枝,亲缘关系最近。
此外,匡叶叶[28]以麋鹿、林麝(Moschusberezovskii)、黑麂(Muntiacuscrinifrons)和豚鹿为研究对象,通过AFLP分子标记,检测遗传多样性的AFLP,基于遗传相似系数矩阵,构建聚类分析,发现麋鹿与豚鹿、梅花鹿的亲缘关系最近。
还有学者认为麋鹿与黇鹿进化关系近。李文斌等[38]利用15个微卫星标记对麋鹿、黇鹿、梅花鹿、赤麂(Muntiacusmuntjak)、马鹿5种鹿类动物群体DNA多态性进行测定,结果发现黇鹿和麋鹿为一类或各自成为一类。
3 麋鹿的遗传多样性研究进展
遗传多样性是生物多样性的核心问题,与生物多样性的形成、消失和发展休戚相关[39]。遗传多样性是评价物种进化潜能与健康状况,以及物种抵御环境变化、生存压力能力强弱的一个重要指标[7,40-41]。遗传多样性的研究工作是生物多样性就地保护的基础,更是迁地保护计划的关键与支撑[42-43]。
从保护遗传学角度看来,物种的长期生存和发展依赖于其遗传多样性和进化潜力的保持,遗传多样性的降低及丧失势必对物种的生存带来不利影响,导致种群生存能力下降和种群退化,甚至灭绝。种内遗传多样性或变异性愈丰富,物种对环境变化的适应能力愈大,其进化的潜力也就愈大。一般来说,濒危物种相比非濒危的广布种,具有较低的遗传多样性。生物种群遗传变异水平和群体遗传结构是其进化历史、分布范围、生活型、繁育方式、扩散机制等各种不同因素综合作用的结果,与其适应性和进化潜力密切相关。特别对于小种群而言,物种遗传多样性主要受到遗传漂变、有效种群大小、瓶颈效应和奠基者效应等因素影响。因此,最大限度地保护种群现有的遗传多样性就显得尤为重要。
我国麋鹿遗传多样性的研究是随着分子标记的发展而逐步深入的,但过去一致的结论是,麋鹿种源的遗传多样性极低[44]。乌邦寺庄园最初的18只麋鹿,性比记载为8雄10雌,至少2只雌性没有参加繁殖[45-46]。根据于长青[44]的推断,当时乌邦寺参与交配繁殖的雄性麋鹿应为2只左右,即现存麋鹿应是最初约 10 只麋鹿的后裔。也有学者认为,在英国乌邦寺出生的子一代麋鹿可能都是同一只雄性麋鹿的后代[44],它们的遗传变异非常单一。1945年,乌邦寺的麋鹿开始向其他地区扩散,Foose等[6]认为此期间麋鹿的近交系数为0.16—0.26,若假定乌邦寺初始麋鹿的近交系数为0,Foose等推算1977年麋鹿的平均近交系数为0.116。麋鹿重引进中国后,于长青[44]对中国麋鹿种群进行有效种群数量分析,结果显示我国麋鹿近交现象极其明显,平均近交系数已高于0.2,这与Sternicki等[47]利用国际物种信息系统(International Species Information System,ISIS)对世界各地动物园登记的1947—2002年间出生的2 042头麋鹿进行研究的结果相符,该研究显示麋鹿近交系数0.242 2—0.281 2,高于兄妹交配的近交系数。麋鹿作为高度近交的种群,在经历了十九世纪末极其严重的瓶颈效应以及其后乌邦寺庄园对外输出的多次间歇性瓶颈效应之后,遗传多样性较低。同时,受到环境条件的制约,使麋鹿的生长受到一定程度的制约,最终导致遗传质量较低[2]。关于中国的麋鹿遗传多样性,学者们进行了一系列的研究。蛋白质电泳分析表明,现在麋鹿种群没有发现明显多态性,遗传多样性严重降低,近乎纯合种群。
到了二十一世纪初期,中国学者通过线粒体、基因片段开展遗传多样性研究。Zeng等[48]利用线粒体DNA控制区及微卫星位点对来自中国的3个已发展稳定的麋鹿种群:北京麋鹿苑、江苏大丰麋鹿国家级保护区、湖北石首麋鹿国家级保护区研究发现,3个种群共享1个线粒体单倍型,目前中国麋鹿在母系遗传上非常单一,没有发生分化;84对跨物种微卫星引物只筛选出5个微卫星位点,且检测结果得出的平均期望杂合度为0.46—0.54,表明我国麋鹿种群的遗传多样性比较贫乏。吴海龙等[49]筛选出8个微卫星位点,构建了麋鹿血液cDNA文库,以用于麋鹿MHC基因及表达基因的研究。张林源等采用25个微卫星位点对中国麋鹿进行群体检测研究,结果显示北京麋鹿生态实验中心的麋鹿种群及其浙江临安的麋鹿亚种群的遗传多样性较低[50]。孟浩等[37]通过分析麋鹿线粒体DNA 控制区序列发现,在所测得的麋鹿样本序列中,同源性达99.81%,单倍型多样度(Hd)与核苷酸多样度(π)都较低,也表明了麋鹿群体多态性不高。匡叶叶[28]通过研究麋鹿个体间遗传距离,发现北京麋鹿苑麋鹿群体具有极高的遗传异质性,种群遗传多样性极低,为一个较为纯合的种群。
目前,关于麋鹿遗传多样性的研究已经进入到基因组水平。Zhang等[51]通过第二代高通量基因组测序技术,首次构建了麋鹿基因组草图,经与马、牛、羊对比,发现麋鹿具有物种特有的69个基因家族、167个基因。Zhu等[52]利用种群基因组测序和比较基因组分析,对麋鹿遗传多样性进行了研究,结果显示,与濒危的朱鹮(Nipponianippon,建群个体数仅为7只)[53]、大熊猫(Ailuropodamelanoleuca,较高的遗传多样性)[54]以及北极熊(Ursusmaritimus)的基因组[55]相比,朱鹮的近交程度最高,麋鹿次之。并认为麋鹿并没有存在很高的近交程度[52](Froh值越大,表示近交的程度越大)(图3),与多年来麋鹿群体的幼体存活率的结果比较一致,推测中国麋鹿种群的77头奠基者可能比朱鹮的7个奠基者具有更多遗传多样性。比较基因组分析显示,麋鹿群体尽管经历过严重的种群瓶颈,但是其仍存留较高的遗传多样性,推测近亲繁殖历史可能有助于清除有害的隐性等位基因。同时,检测到一批受到正选择的基因,涉及对高盐食物的潜在适应、血压调节、心血管发育、胆固醇调节、血糖控制和甲状腺激素合成,推测这些适应性遗传特征有助于缓解与高盐饮食相关的不利影响。

图3a 麋鹿、朱鹮、大熊猫与北极熊种群ROH基因组长度比例Fig.3a Box plot of Froh for Père David’s deer,crested ibis,panda,and polar bear populations.Froh denotes the proportion of total ROH length

图3b 麋鹿、朱鹮、大熊猫与北极熊个体ROH基因组长度比例(引自Zhu等,2018)[52]Fig.3b Box plot of length of ROH in each individual from Père David’s deer,crested ibis,panda,and polar bear
4 小结
麋鹿是中国重点保护的小种群代表动物之一,现存的麋鹿均为达氏种。2018年的研究结果,建立了麋鹿种之下的一个亚种——原达氏鹿[16],相比较与现生的麋鹿,新发现的这类麋鹿的特点是鹿角前枝比后枝更纤细短小,其前枝的分支点距离前枝与后枝的分支点较近。通过对麋鹿化石种分类的讨论,将麋鹿属分为4个种和8个亚种,对研究麋鹿属在中国的演化提供了重要证据。
根据考古研究,鹿亚科在中新世早期从鹿科总进化枝分化出来,而鹿属在上新世末期(Terminal Pliocene)至更新世初期起源并逐步分化。麋鹿出现于更新世早期(Early Pleistocene),与鹿属其他物种的分歧时间大约在 270 万年[26]。坡鹿是中国鹿属中最原始的种,与鹿属其他种的分歧时间在270万—320万年。水鹿与白唇鹿、梅花鹿和马鹿的分歧时间在 200万—240 万年。梅花鹿与马鹿的分歧时间在 140万—160 万年。大多数学者认为麋鹿与鹿属的亲缘关系近,尤其是与坡鹿的亲缘关系最近。
麋鹿在经历了十九世纪末极其严重的瓶颈效应以及其后乌邦寺庄园对外输出的多次间歇性瓶颈效应之后,中性遗传多样性基本丧失,种群内个体之间的遗传相似系数极高,理论上极度缺乏维持其可持续发展的生存潜力,麋鹿存在着近交衰退而走向灭绝的威胁。但是,从乌邦寺建群到现今遍布世界的多个小种群来看,由于中国政府的大力支持与保护,加上麋鹿对环境的潜在适应,麋鹿的数量一直在逐步增长,且未发现有明显的生存力和繁殖力下降的情况,说明麋鹿群体复壮取得初步成功。
目前,严重的近交似乎对麋鹿的种群繁衍未产生明显的不良影响。根据北京麋鹿苑的记载,近20年来幼体存活率一直维持在80%以上,成体体重以及雄性个体鹿角明显增大,子代发生畸形和母兽难产现象出现的概率都非常低。理论上,近交系数每增加10%,种群繁殖能力将下降25%左右,一方面,近亲繁殖似乎并没有对麋鹿的繁殖力和生存力产生明显的影响。有学者认为,现存麋鹿在建群初期,种群内部发生了积极的遗传净化作用,结合麋鹿种群迅速增长和繁殖过程中的有效竞争选择机制等,导致种群内部的稀有有害等位基因被淘汰,从而使种群内优良基因得以保存,耐受近交效应的能力增强[4,52]。另一方面,遗传多样性水平的降低也可能使麋鹿抵抗随机风险的能力随之降低,易受到自然灾害及疫病的严重威胁,湖北石首麋鹿保护区2008年受到雪灾的影响,造成大批麋鹿死亡。2010年北京南海子麋鹿苑和湖北石首麋鹿国家级自然保护区由于受魏氏梭菌(Clostridiumwelchii)的影响,麋鹿种群猝死率分别为10.87%和23.35%[56]。麋鹿是高度特化的物种,如果不加以人为打扰,一旦面临剧烈的条件改变,极可能由于缺乏自身的适应调节机制,遭受物种绝灭危险.这也是尽管全世界各地目前的麋鹿数量有了较大增长,但是并不足以将其从濒危物种名单中删除的原因[36]。
尽管麋鹿线粒体DNA多样性研究显示其具有极低的遗传多样性,但从其基因组水平多样性显示,中国麋鹿种群的77只奠基者比朱鹮的7只奠基者[57]具有更多遗传多样性[51-52]。考虑到朱鹮和麋鹿现存种群的复壮成果,建议进一步开展核基因水平和基因组水平多样性方面的研究工作,以促进我国麋鹿种群的健康可持续繁衍。
猜你喜欢
初中生学习指导·中考版(2020年12期)2020-09-10 07:22:44
武夷科学(2019年2期)2019-12-20 08:39:16
小哥白尼(野生动物)(2018年1期)2018-05-26 09:28:03
小哥白尼·野生动物画报(2018年1期)2018-02-05 16:49:12
武夷科学(2017年0期)2017-03-10 10:03:31
新疆大学学报(自然科学版)(中英文)(2016年4期)2016-05-16 05:56:12
中国畜牧兽医文摘(2014年5期)2014-08-15 00:50:09
幼儿智力世界(2006年5期)2006-05-11 03:06:20
幼儿智力世界(2006年5期)2006-05-11 03:06:20
幼儿智力世界(2006年5期)2006-05-11 03:06:20
