
山药根际土壤微生物16S rRNA多样性及影响因素*
2019-11-18 02:23:54张红霞张舒雅张玉涛张天缘
土壤学报 2019年5期
张红霞 张舒雅 张玉涛 张天缘
(1 安顺学院化学化工学院,贵州安顺 561000)
(2 安顺学院/贵州省科技厅土壤污染控制与修复工程技术中心,贵州安顺 561000)
(3 武汉贝纳科技服务有限公司,湖北武汉 430070)
山药(Dioscorea opposita)为薯蓣科多年生草质藤本植物,其地下根茎为主要利用器官,是我国传统的药食同源植物。现代药理研究表明,山药具有抗衰老、调节免疫、抗肿瘤、降血糖等作用,其主要化学成分有多糖、尿囊素、皂甙等[1]。我国栽培山药历史悠久,由于栽培的地域环境条件不同和人工选择的原因,形成了各具特色的地方品种。贵州安顺山药因口感软糯,富含蛋白质、多糖等成为大众喜爱的品种之一。土壤生态环境因子、养分状况是影响植物生长及品质形成的主要因素,随着人们对土壤微生物生态因子重要性认识的不断深入,将土壤微生物学特性作为评价土壤质量和健康程度的相应指标已广受关注,其结果可用于指导生产实践[2]。陆地生态系统中的土壤微生物群落多样性受到土壤养分的显著影响[3]。研究表明,山药连作会破坏土壤耕层微生物种群结构,抑制土壤酶活性,可能导致山药连作障碍[4]。随着山药种植规模的扩大,为保证其优良品质,推行规范化种植,开展了大量栽培管理措施、施肥及土传病害防治方面的研究,然而结合土壤微生物群落结构及其多样性,分析其与土壤化学性质、根茎营养成分关系方面的研究鲜有报道。高通量测序技术具有可全面、准确获得微生物群落结构信息的特点,已被广泛应用于微生物研究中[5-7]。本研究采用高通量测序技术,通过提取不同环境土壤样品的DNA,选择合适的通用引物扩增16S的目标区域,检测目标区域的序列变异和丰度,分析环境中细菌群落结构及其多样性,结合根际土壤养分状况及部分酶活性的检测,研究山药根际土壤中细菌群落特征与土壤养分、根茎糖类含量的关系,阐明影响山药根茎中营养物质累积的重要土壤生态因子,对于研究山药种植专用生物肥料及其种植土壤环境评价提供数据参考。
1 材料与方法
1.1 供试材料
前期对安顺4个山药种植区山药营养成分的分析发现,不同种植区的山药根茎在灰分、粗纤维和蛋白质的质量分数上有显著差异[8],因此本研究选取安顺市辖区内7个集中种植山药的村寨(表1)采集样本,每个村寨选取连续种植山药2年的有代表性的5个地块作为样本(5个重复)。于2017年3月10日至30日期间整地并施基肥后,用薯蓣科薯蓣属植物山药(经安顺学院农学院沈昱翔副教授鉴定)芦头种植。于2017年10月3日至15日期间采集土壤样品及山药样品,每个地块按5点取样法,将山药根茎采挖出后,抖落根茎上附着的土壤作为根际土壤,混匀后储存于无菌自封袋中,按表1编号后低温运输,-20℃冰箱保存。土壤均为黄棕壤。

表1 研究样本基本情况Table 1 Basic information of the experiment plots of the study
1.2 16S rRNA基因扩增、文库构建及测序
采用土壤DNA提取试剂盒(北京索莱宝科技有限公司)提取土壤样本总DNA,经紫外分光光度计(上海元析仪器设备有限公司)测定纯度,1%琼脂糖凝胶电泳检测后,用DNA回收试剂盒回收,于-20℃保存备用。
以提取的总DNA为模板,采用16SrDNA V3-V4区引物(上海生工生物工程技术服务有限公司):338F:(5′-ACTCCTACGGGAGGCAGCAG-3′),806R:(5′-GGACTACHVGGGTWTCTAAT-3′)进行PCR扩增(上海精宏实验设备有限公司)。PCR反应条件:94℃预变性4 min,94℃变性30 s,55℃退火30 s,72℃延伸45 s,35个循环,最后72℃延伸10 min。PCR产物采用琼脂糖凝胶DNA回收试剂盒(TIANGEN)回收目的条带,使用Qubit荧光定量系统对回收产物进行检测定量,等量混合不同样品PCR 回收产物,构建高通量测序文库,IlluminaHiseq 2500上机测序。
1.3 根际土壤生态指标及根茎糖类指标检测
土壤化学性质采用常规分析方法[9],总有机碳采用高温外热重铬酸钾氧化容量法,碱解氮采用碱解扩散法,有效磷采用0.5mol∙L-1NaHCO3浸提—钼蓝比色法,速效钾采用1mol∙L-1NH4OAc浸提—火焰分光光度法,pH用电位法(酸度计)。
土壤酶活性测定[10]:过氧化氢酶活性测定采用容量法(以KMnO4计,mL∙g-1∙30min-1),蔗糖酶活性测定采用3,5-二硝基水杨酸比色法,脲酶活性测定采用靛酚蓝比色法,磷酸酶活性测定采用磷酸苯二钠(4-氨基安替比林)比色法。
山药根茎糖类指标检测:可溶性糖、淀粉含量测定[11]采用硫酸蒽酮法和高氯酸法;粗多糖测定[12]采用85%乙醇除去低聚糖、单糖、双糖等,用95℃水浴水提后硫酸蒽酮显色,于620 nm波长下比色法测定;粗蛋白含量测定采用GB/T5009.5-2010中的分光光度法。
1.4 数据处理
样品经IlluminaHiseq 2500测序进行PE250双端测序后,对获得的原始测序序列进行过滤和质控,通过质量分值衡量测序序列的质量和拼接效果,选取长度大于100bp,平均质量分值>20,拼接序列重叠区域错配率小于20%的序列作为有效序列,进行下游分析。以97%水平的SILVA数据库为参考,计算OTUs的丰度。
采用QIIME软件进行α多样性分析,包括香农指数、Chao1指数、文库覆盖率。采用QIIME分析流程,使用UniFrace方法进行β多样性分析。
采用Vegan软件进行典型相关分析(Canonical Correlation Analysis,CCA)。使用R软件做环境因子与物种丰度的相关性分析。
2 结 果
2.1 根际土壤生态特性与根茎营养成分含量
从表2可见,7个样本土壤偏酸性,且其养分含量及酶活性均有显著差异。表3可见,7个样本的山药根茎中4种营养成分含量也呈现一定差异。对根际土壤的化学性质与山药根茎中营养成分相关性的分析(表略)可知,碱解氮与有机质呈极显著正相关(相关系数为0.877),过氧化氢酶与有效磷呈显著正相关(相关系数为0.811),山药根茎中粗多糖与根际土壤的有机质呈显著正相关(相关系数为0.770),可溶性糖与土壤有机质、全氮及碱解氮含量呈极正显著相关或显著正相关(相关系数分别为0.942、0.810、0.790),上述结果说明土壤碳、氮养分能够促进山药根茎中糖的积累。
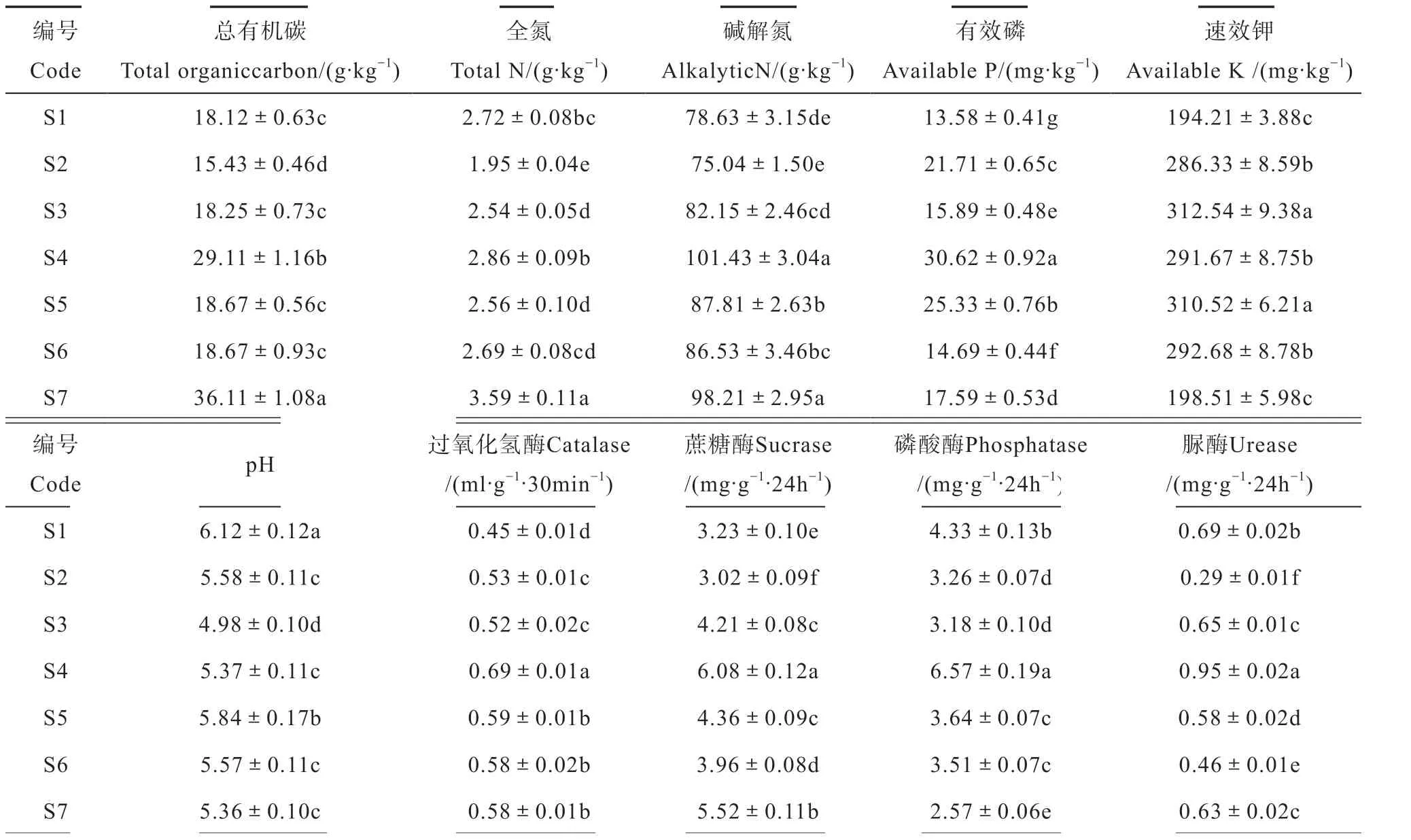
表2 山药根际土壤生态特性Table 2 Ecological characteristic of the rhizosphere soil of Chinese yam
2.2 物种相对丰度统计
高通量测序数据经过滤、质控后,对7个样本随机抽取的有效序列所对应的稀释曲线分析,结果表明深度测序已接近饱和,可用于后续分析。在97%相似水平下的OTU进行生物信息统计分析。图1显示了“门”水平的物种相对丰度。从统计结果可知,相对丰度大于1%的菌群有9个,分别为变形菌门(31.0%)、绿弯菌门(22.8%)、酸杆菌门(22.7%)、放线菌门(8.6%)、拟杆菌门(4.3%)、厚壁菌门(3.1%)、奇古菌门(1.5%)、芽单胞菌门(1.3%)和硝化螺旋菌门(1.1%)。从数据分析结果可见,变形菌,绿弯菌和酸杆菌、放线菌是优势种群(相对丰度>5%),它们的相对丰度均大于5%,占了总序列的85.1%;此外还发现了相对丰度小于1%的其他44个稀有门类,但分布不均匀,仅在部分样本中发现。在“属”水平上进行分析,7个样本中均发现慢生根瘤菌属(1.4%),5个样本(S2, S3, S5, S6,S7)发现假单胞菌属(0.4%)。从结构来看,表现为7个样本共有大多数相同的细菌群落,但也存在一定差异。已有研究表明土壤pH、有机碳和全氮是土壤细菌群落结构和多样性的主导因子[13],据此分析认为本研究7个样本的细菌群落结构存在差异的原因可能与土壤理化性质有关。

表3 山药根茎中营养成分含量Table 3 Nutrient contents in rhizome of Chinese yam /(g∙100g-1)
2.3 α多样性指数与β多样性聚类
香农指数反映物种丰富度和均度的影响,用于说明微生物群落分布的均匀度,其数值大小主要衡量非均衡分布状况。Chao1指数仅简单指群落中物种的多样性,不考虑丰度情况。对表4的两种多样性指数进行比较,结果表明S5、S6、S7这3个样本的细菌群落物种丰度、多样性和均度比较接近;S1、S2和S4样本的在一个水平;而S3样本的菌群丰度、多样性与均度与其余样本比较处于中间。从细菌群落多样性指数上可将7个样本分为3个类别。经相关性分析发现,香农指数与Chao1指数和土壤磷酸酶活性为显著相关(相关系数分别为-0.644**和-0.506*),说明在山药根际土壤中细菌群落的多样性对磷酸酶活性有较大影响。
在β多样性研究中,PCoA(Principal coordinate analysis)是基于UniFrace的主坐标分析,可直观显示不同样品中微生物进化上的相似性或差异性。图2显示了在菌群β多样性中各样本的特征贡献值,其中S5、S6、S7样本的菌群多样性占71.44%,为菌群多样性的主要贡献者,与α多样性分析结果相互印证。
2.4 菌群丰度统计
采用LEfSe分析检测不同组别中具有显著丰度差异特征的菌群。从图3a可知,除平寨村(S2)外的6个样本在多个菌群丰度上存在显著差异,其中放线菌、绿弯菌、酸杆菌、变形菌分别在不同组别中起重要作用,说明这几个优势菌群对于土壤细菌群落结构产生重要影响。图3b将7个细菌门在6个不同样本中的门、纲、目、科、属水平的差异信息用饼形图表示,其中最大的变形菌门菌群类别较多(见图3b,a1~a9,b0~b4),且差异明显;乐平村(S6)土壤中所含细菌群落种类较丰富,且与其他样本的差异明显,与多样性指数分析的结果一致。
2.5 环境因子、根茎糖类物质与土壤细菌物种分布的相关性
CCA分析结果可反映菌群与环境因子之间的关系,从中可知影响样品分布的重要环境驱动因子。
从图4a可见,土壤化学指标可聚为两类,其中pH与样本中细菌群落分布的相关性最大,且位于第Ⅲ、Ⅳ象限之间的轴上,与AP、AK、TOC、TN呈正相关,而与碱解氮呈负相关。

图1 各样本在门水平的物种相对丰度条形图Fig.1 Relative abundances of the species of bactera in the samples at the phylum level in bar chart

图2 样本主坐标分析Fig.2 Sample principal coordinate analysis

表4 α多样性指数统计Table 4 Statistics of α diversity indices
图4b中优势菌群绿弯菌、奇古菌与TOC、AK、TN、AP和pH这5个环境因子呈正相关;而酸杆菌与pH、AP呈负相关,与TOC、AK呈正相关,进一步说明土壤pH因子是影响土壤细菌群落分布的主要环境驱动因子之一,且也是影响土壤养分状况的重要因子。碱解氮单独归为一类,位于第Ⅱ象限,与其他环境因子呈负相关,与硝化螺旋菌高度正相关,说明了土壤中硝化细菌在促进氮素同化的同时,可能对土壤中其他养分因子将会产生一定抑制作用。

图3 菌群统计结果Fig.3 Statistics chart of microflora

图4 山药根际土壤理化指标与样本及细菌群落CCA分析Fig.4 CCA analysis of soil physicochemical indexes and bacterial communities in yam rhizosphere
几种土壤酶分别位于4个象限,相互之间的相关性表现不强(图4b)。优势菌群奇古菌和绿弯菌与磷酸酶呈正相关关系;酸杆菌与蔗糖酶呈较高正相关关系,但与过氧化氢酶为负相关;硝化螺旋菌和放线菌与脲酶和过氧化氢酶均呈正相关关系,但又与磷酸酶和蔗糖酶呈负相关,说明土壤中几种酶活性对优势菌群有不同程度的影响,且这种影响比较复杂,这可能跟土壤酶活性的时空异质性[13]及菌群结构复杂性有关。
山药根茎中的3种糖类物质相互之间有一定相关关系,其中淀粉与可溶性糖、粗多糖为正相关,而粗多糖与可溶性糖为弱负相关,说明粗多糖的累积一方面随可溶性糖增加而增加,另一方面随着粗多糖的累积可能会抑制可溶性糖的积累。优势菌群中奇古菌和绿弯菌与淀粉正相关,并且这两种菌群与粗多糖为显著正相关(相关系数为0.54,P<0.01);变形菌和硝化螺旋菌与可溶性糖为正相关,与粗多糖为负相关;酸杆菌与可溶性糖的夹角几乎呈180°,说明这个菌群与可溶性糖呈负相关关系。上述结果说明在山药根际土壤中的奇古菌和绿弯菌菌群利于多糖和淀粉的累积。
3 讨 论
土壤不仅提供了细菌群落生长繁殖的微环境,其理化性质又间接影响土壤细菌群落结构的组成。土壤微生物参与了土壤中的碳、氮等营养素的循环和能量流动,参与了腐殖质的形成、土壤结构的形成与改良,是土壤生态的重要组分,也是植物生长及品质形成的主要影响因子。本研究在山药根际土壤中发现的4个优势菌群与于海玲[14]、金奇[15]的研究结果相似,均有变形菌、酸杆菌和放线菌,是多数酸性土壤中存在的菌群,这是因为土壤细菌群落是一个相对稳定的系统,施肥或连作等条件可能只影响某些种群的丰度,而没有改变群落种类[16]。此外,这些优势菌群参与土壤的氮素循环[17-18],有利于植物的生长。本研究中7个样本的优势菌群基本一致,而菌群丰度及结构呈现一定差异。
在土壤化学指标与细菌群落关系研究中,本研究结果表明,土壤pH是影响土壤细菌群落分布的重要因子。朱平等[19]对祁连山中部4种典型植被类型土壤细菌群落结构与理化性质相关性的研究结果表明,土壤细菌群落结构与土壤pH、含水量、总有机碳和总氮呈显著正相关,且土壤pH对细菌群落结构的影响最大,这与本研究结果一致。土壤细菌的影响因子大小为土壤有机碳、土壤总氮、含水量、电导率等,其中土壤有机碳和土壤总氮有显著性影响[20]。王鹏等[21]的研究发现总磷、铵态氮和有机质含量是与细菌群落相关性较大的土壤化学指标。本研究发现酸杆菌、绿弯菌、奇古菌和硝化螺旋菌分别与土壤的总有机碳(TOC)、AK、TN、AP、AN相关,说明细菌群落结构受土壤化学因子的影响,与赵丰云等[22]的研究结果一致。微生物通过向土壤分泌胞外酶及死亡后细胞自溶产生的胞内酶推动土壤中的生物化学反应,因此土壤酶活性及土壤化学指标相互间存在着密切配合和相互促进的复杂关系。本研究分析表明,酸杆菌与蔗糖酶和TOC均为正相关关系,蔗糖酶参与土壤中的碳循环,推测酸杆菌能够促进蔗糖酶活性,从而增加土壤中TOC的积累;硝化螺旋菌与脲酶和碱解氮为正相关,脲酶参与土壤中的氮循环,据此推测硝化螺旋菌通过增加脲酶活性,从而增加土壤中碱解氮含量。
本研究对山药根茎中几种糖类物质含量与土壤中部分优势菌群相关关系的分析表明,山药粗多糖的积累与土壤中的奇古菌和绿弯菌菌群显著正相关。奇古菌是近20年来在中温环境中发现的广泛大量存在的一类古菌,它们可能在土壤C、N的生物地球化学循环中发挥重要作用[23],其中的氨氧化古菌是酸性土壤中硝化作用的主要推动者[24],绿弯菌通过光合作用进行自养,增加土壤碳素,推测可能这些细菌通过改善土壤中的C、N循环来促进山药根系糖类物质的运输及贮藏。综上所述,土壤中酸杆菌、硝化螺旋菌、奇古菌和绿弯菌这4类细菌主要通过改善土壤中的碳、氮养分状况,促进山药根茎对可溶性糖、粗多糖及淀粉的积累。
高通量测序技术对山药根际土壤中细菌群落多样性的分析,以及细菌群落与土壤养分、山药根茎主要营养成分的关系分析结果显示,它们之间存在一定相关性,但由于土壤微生物与微生物之间、与土壤养分之间及与植物之间存在十分复杂的相互作用关系,研究这些相互作用关系并揭示其作用机理需要进行更加深入细致的研究,因此今后可分别就几种优势菌群、菌种组合与土壤理化特征、山药根茎营养成分等的互作进行机理研究和应用研究。
4 结 论
对山药根际土壤微生态环境及根茎营养成分的研究表明,其优势细菌群落为变形菌、绿弯菌、酸杆菌和放线菌;土壤pH是影响细菌群落结构的重要因子;酸杆菌和硝化螺旋菌可能通过促进土壤中蔗糖酶、脲酶活性提高土壤的碳、氮水平,奇古菌和绿弯菌促进根际土壤的C、N循环,增加土壤C、N养分,土壤中碳、氮养分能够促进山药根茎中可溶性糖、粗多糖及淀粉的积累。
猜你喜欢
中国土壤与肥料(2021年5期)2021-12-02 01:06:26
基层中医药(2020年9期)2020-11-27 01:55:28
今日农业(2020年13期)2020-08-24 07:35:28
文苑(2020年6期)2020-06-22 08:41:50
创新作文(小学版)(2019年22期)2019-12-07 05:44:40
天然产物研究与开发(2018年3期)2018-05-07 06:38:35
中成药(2017年3期)2017-05-17 06:09:00
中国蔬菜(2016年8期)2017-01-15 14:23:38
文苑(2015年5期)2015-05-11 10:47:30
天然产物研究与开发(2014年3期)2014-04-27 14:15:29
