
亚热带典型林分空间结构与林下草本物种多样性的差异特征分析及其关联度
2019-11-08 06:09曹小玉李际平
草业科学 2019年10期
曹小玉,李际平,委 霞
(中南林业科技大学林学院,湖南 长沙 410004)
森林是地球上最大、最重要的陆地生态系统,其物种多样性和保护一直是备受关注的生态学研究热点[1]。森林物种多样性包括森林生态系统乔、灌、草的物种多样性,其中林下草本层占据森林生态系统物种丰富度的90%以上[2],其对森林生态系统净初级生产力的贡献约是其地上生物量所占比例的20倍[3],是影响乔木层幼苗更新、防止水土流失和促进土壤养分元素(N、P、K等)循环的重要因素。但同时草本层物种灭绝率高达乔木的3倍[4]。因此,保护林下草本多样性对维持森林生态系统物种多样性具有十分重要的作用。作为森林生态系统主体的乔木层,其林木空间结构直接影响着林下草本物种生长的微环境,决定着草本的生长、繁殖和死亡以及林窗的形成[5]。进而影响着林下草本物种多样性。Thoms等[6]认为,如果忽略林分空间细节特征,维持林下生物多样性是不可能的。相比传统的林分非空间结构因子(林分年龄、密度、郁闭度、平均树高、平均直径等),林分空间结构(混交度、角尺度、竞争指数、林层比等)是表征林木的点格局及其属性的空间分布指标[7],是森林经营过程中最易调控的因子,研究林分空间结构对林下草本物种多样性的影响对揭示林分空间结构对林下草本的影响机理,维持林下草本物种多样性具有重要的理论和实践意义。
现阶段已有大量关于影响林下草本物种多样性因子的研究,Sabatini等[8]采用多元回归模型研究了森林结构、光照条件、土壤和地形特征等环境因素对山毛榉(Fagus sylvatica)林下草本物种多样性的影响;朱媛君等[9]在张北杨树人工林研究了树高、胸径、冠幅、密度等林分结构因子对林下草本物种多样性的影响。总体来说,这些研究主要集中在林分非空间结构因子和环境因子。而针对林分空间结构因子对林分林下植被物种多样性的研究却较少。曹小玉等[10]采用单因素分析了林层指数对杉木林下灌木物种多样性的影响;朱光玉等[11]采用典型相关性分析方法研究了林分空间结构对湖南栎类林下灌木物种多样性的影响,但其研究方法都是建立在变量之间为线性关系假设上的统计方法,无法科学地分析林分空间结构对林下草本物种多样性的影响。灰色系统分析方法是根据研究对象之间信息发展态势的相似或相异程度来揭示研究对象系统内部信息之间的关联程度,对研究“部分信息已知,部分信息未知”的“小样本”、“贫信息”的不确定性系统有着其他方法不可比拟的优势[12]。
确定林分空间结构单元是计算林分空间结构参数的基础,而邻近木株数的确定则是构建林分空间结构单元的关键,基于Voronoi图确定邻近木株数更具有灵活性,它克服了基于4株邻近木确定林分空间结构单元时可能将中心木的邻近木排除在外或将非邻近木计算在内的缺陷[13]。基于此,本研究以亚热带3种典型林分为例,在采用Voronoi图确定空间结构单元的基础上,量化分析3种典型林分的空间结构指数和林下草本物种多样性指数,采用灰色关联度分析林分空间结构和林下草本物种多样性的关系,以期为揭示林分空间结构对林下草本的影响提供基础数据,也为提高林下草本物种多样性为目标的林分空间结构调控提供科学的理论依据。
1 研究区概况
福寿林场位于湖南省平江县南部福寿山上,地处28°03′00″ - 28°32′30″ N,113°41′15″ - 113°45′00″ E。最高海拔1 573.2 m,最低海拔835 m,林场场部海拔1 078 m。山体下部多陡峭,中部较平缓,上部较陡,平均坡度22°~27°。处于中亚热带向北亚热带过渡的气候带,属湿润的大陆季风气候。年平均气温12.1 ℃,年降水量2 100~2 300 mm,年日照1 500 h,无霜期217 d,年相对湿度87%。场内海拔800 m以下的土壤为山地黄壤,800~1 400 m为山地黄棕壤,海拔1 400 m以上的山顶、山脊有小块草甸土。适宜多种树木生长,是汨罗河和浏阳河的重要水源涵养林区,森林类别属于公益林。
2 数据来源与研究方法
2.1 数据来源
在对研究区典型林分全面踏查的基础上,采用罗盘仪闭合导线测量法在立地条件基本一致的杉木纯林、杉木混交林和天然次生林中分别设置了6块20 m × 30 m的样地,一共18块样地(表1)。将每块样地分成6个10 m × 10 m的小样方作为林分乔木层的调查单元,调查因子为小样方内每株树木的坐标、胸径、树高、东西冠幅、南北冠幅等基本因子。在每个样地四角和中心地带设置5个1 m × 1 m草本样方,调查草本层的物种名称、高度、盖度、株(丛)数等因子。调查可知,3种典型林分林下草本主要物种有鸭跖草(Commelina communis)、博落回(Macleaya cordata)、五节芒(Miscanthus floridulus)、金星蕨(Parathelypteris glanduligera)、十字苔草(Carex cruciata)、竹叶草(Oplismenus compositus)、铁芒萁 (Dicranopteris dichotoma)、江南星蕨(Microsorum fortunei)、 鱼 腥 草 (Houttuynia cordata)、狗脊(Cibotium barometz)、山麦冬(Liriope spicata)、翠云草(Selaginella uncinata)、鸡矢藤(Paederia scandens)、野青茅(Deyeuxia arundinacea)等。

表1 各样地基本概况Table 1 Basic conditions of the sample plots
2.2 研究方法
2.2.1 林分空间结构单元的确定
本研究采用Voronoi图来确定林分空间结构单元中邻近林木的个数n,基于样地林木的坐标信息,借助ArcGIS由样地单株林木构建的Voronoi图,经距离缓冲区法(2 m缓冲区)边缘矫正后,如图1所示。

图1 林木点数据生成的Voronoi图Figure 1 Voronoi diagram based on the tree data
2.2.2 空间结构指数的计算
本研究采用混交度、角尺度、林层指数和林木竞争指数4个林分空间结构指数来分析林分的空间结构,每个参数代表的含义和计算公式如下:
1)混交度是用来说明林分内树种隔离程度的指标[14],其计算公式如下:

2)角尺度是反映林木空间分布格局的指标[15],其计算公式如下:

3)林层指数是反映林层多样性的指标[16],其计算公式如下:

4)竞争指数是反映林木个体所承受竞争压力大小的指标[17],本研究采用Hegyi竞争指数,计算公式如下:

2.2.3 物种多样性指标的计算
本研究采用物种丰富度指数(S)、物种多样性指数和均匀度指数来分析林下草本物种多样性水平。其计算公式如下:

式中:Pi为灌木第i种的个体数占所有种的个体总数的比例,S为所在样地内灌木物种种类的总数。
2.2.4 灰色关联度
灰色关联度分析是将林分林下草本物种多样性及其影响因子林分空间结构视为灰色系统,以林下草本物种多样性因子作为参考数列,将影响林下草本物种多样性的林分空间结构因子作为被比较数列,来分析林分空间结构因子与林下草本物种多样性的关联程度。则其关联系数公式如下:


2.2.5 数据处理与分析
数据分析利用SPSS 19.0和DPS 14.50软件,采用初值化的方法来消除各指标数据的量纲,采用ANOVA(方差分析)和LSD(最小显著差异法)对3种林分类型的林分空间结构与林下草本物种多样性进行差异显著性分析,图表绘制与数据处理在Microsoft Excel 2013中完成。
3 结果与分析
3.1 不同林分空间结构分析
天然次生林、杉木混交林和杉木纯林的混交度分别为强度混交(0.695 9)、中度混交(0.427 0)和弱度混交(0.194 5),天然次生林的树种隔离程度明显优于杉木混交林和杉木纯林,杉木纯林的林分混交度最低,且3种林分的混交度存在显著性差异(P < 0.05) (图2)。天然次生林、杉木混交林和杉木纯林的角尺度分别为0.346 8、0.356 1和0.346 7,均接近随机分布,且3种林分的角尺度间差异不显著(P > 0.05)。天然次生林、杉木混交林和杉木纯林的林层指数分别为0.614 7、0.428 7和0.259 4,天然次生林树高分化程度明显大于杉木混交林和杉木纯林,而杉木纯林对垂直空间利用程度最弱,且3种林分的林层指数存在显著性差异。天然次生林、杉木混交林和杉木纯林的竞争指数分别为4.736 1、6.039和6.632,天然次生林的林木承受的竞争压力明显小于杉木混交林和杉木纯林,且天然次生林与杉木纯林的竞争指数存在明显差异,而与杉木混交林无显著差异。

图2 3种林分的空间结构指数Figure 2 Spatial structure index of the 3 stand types
3.2 不同林分林下草本物种多样性分析
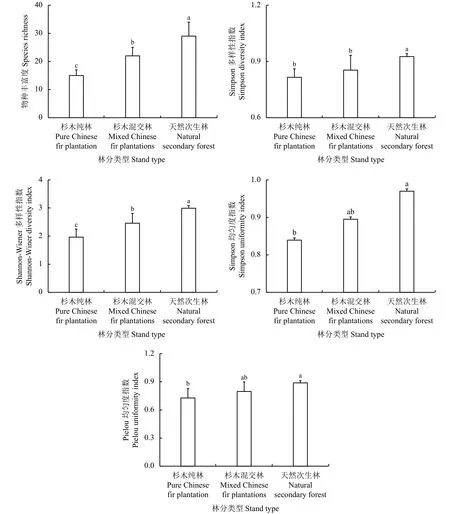
图3 3种林分的林下草本物种多样性指数Figure 3 Understory herbaceous species diversity index of the 3 stand types
天然次生林、杉木混交林和杉木纯林林下草本的物种数分别为29、22和15种,天然次生林林下草本的物种丰富度远大于杉木混交林和杉木纯林林下草本的物种丰富度,几乎是杉木纯林林下草本物种多样性的2倍和杉木混交林的1.3倍,且3种林分的林下草本物种丰富度存在显著性差异(P < 0.05) (图3)。天然次生林、杉木混交林、杉木纯林的林下草本的Simpson多样性指数和Shannon-Wiener多样性指数依次分别为0.925 9、0.853 7、0.815 1和2.990 3、2.458 0、1.961 0,3种典型林分的林下草本的Simpson多样性指数和Shannon-Wiener多样性指数都表现出了天然次生林 > 杉木混交林 >杉木纯林的规律。且3种林分的林下草本Shannon-Wiener多样性指数存在显著性差异,杉木天然林和杉木纯林、杉木混交林的林下草本的Simpson多样性指数也分别存在显著性差异。天然次生林、杉木混交林、杉木纯林林下草本的Simpson均匀性指数和Pielou均匀度指数依次分别为0.969 8、0.895 0、0.839 1和 0.889 0、0.795 0、0.728 1, 3种典型林分的林下草本的Simpson均匀性指数和Pielou均匀度指数同样都表现出了天然次生林 > 杉木混交林 > 杉木纯林的规律。且杉木纯林和天然次生林林下草本的Simpson均匀性指数和Pielou均匀度指数都存在显著差异性。
3.3 不同林分空间结构与林下草本物种多样性的灰色关联度分析
在3种不同的林分类型中,林分空间结构指标与林下草本物种多样性的关联度存在差异(表2)。在杉木纯林中,林分混交度与物种丰富度、Simpson多样性指数、Simpson均匀性指数的关联程度最大,林分角尺度与Shannon-Wiener多样性指数、Pielou均匀度指数的关联程度最大。影响杉木纯林林下草本物种多样性的主要空间结构因子是混交度和角尺度。在杉木混交林中,林分混交度与物种丰富度、Shannon-Wiener多样性指数、Pielou均匀度指数的关联程度最大,林分竞争指数与Simpson多样性指数、Simpson均匀性指数的关联程度最大。影响杉木混交林林下草本物种多样性的主要空间结构因子是混交度和竞争指数。在天然次生林中,林分混交度与物种丰富度、Pielou均匀度指数的关联程度最大,林分角尺度与Simpson多样性指数、Shannon-Wiener多样性指数、Simpson均匀性指数的关联程度最大。影响天然次生林的林下草本物种多样性的主要空间结构因子是角尺度和混交度。总体看来,林分混交度,即林木隔离程度是影响3种林分林下草本物种多样性的共同关键因子。

表2 林分空间结构与林下草本物种多样性的灰色关联度及其排序Table 2 Correlativity and sort of the understory herbaceous species diversity and stand spatial structure
4 讨论与结论
4.1 讨论
不同林分类型的林木在树种、大小、水平分布和垂直分布等方面都存在差异,导致林分空间结构各异,天然次生林是森林生物与环境相互影响、相互依存、长期进化的结果,形成了异龄、混交、复层较为理想的空间结构,而人工林由于受到采伐方式和更新方式的限制,基本上都是单层同龄的混交林或纯林,林分空间结构较差[19]。研究区的天然次生林的树种隔离程度(混交度)、林层多样性(林层指数)、竞争态势(竞争指数)和林木空间分布格局(角尺度)最优,杉木混交林次之,杉木纯林最差。这在一定程度上是因为林分空间结构的形成与林木更新方式和物种竞争有很大关系,天然次生林在生长演替过程中,同一树种由于激烈争夺有限环境资源产生自疏现象,导致其他物种侵入形成多树种混交,林木趋于随机分布的理想状态,自然更新形成多径级和多龄级林木同时并存的分布状态,使林木垂直结构分化明显,垂直空间利用充分[20]。而起源于人工造林的杉木纯林和混交林中杉木均占绝对优势,占据林分水热资源丰富的上层空间,导致林下喜光树种更新发育困难,这既限制了林木混交度的提高和林木水平分布向随机分布演变,也影响了林木对垂直空间的充分利用[21]。
不同森林类型的林分空间结构对林下草本物种多样性的影响程度并不相同,不同的林分空间结构代表着乔木层不同的树种隔离程度、不同的林层结构、不同的水平分布格局和竞争态势,导致了不同林分林下光照、土壤养分条件、土壤质地和凋落物性质的综合差异,从而间接地对林下草本的物种多样性产生影响[22]。相比于杉木纯林和杉木混交林,研究区的天然次生林有着较高的林下草本物种多样性。这是因为天然次生林水平方向多树种混交和林木垂直方向上树冠重叠交错,一方面使林地内形成大量较易分解的枯枝落叶,增加土壤有机质和肥力[23];另一方面复杂的垂直结构强烈地影响林分中不同高度和林下的总有效光强和光斑位置,使林下草本耐阴物种、需光量大的非耐阴的物种及其普通物种都能广泛存活,从而林下草本物种多样性较高[24]。而研究区的杉木纯林是树种单一的单层同龄林,林下光照和土壤资源条件在时空上的分布差异不明显,再加上针叶凋落物坚硬偏酸性,造成表层土壤养分低下,严苛的环境条件限制了需要大量营养物和光照植物的发展,导致林下草本生物多样性偏低[25]。杉木混交林虽是单层同龄林,但不同混交树种的生态位相互抑制,为林下草本的生长释放了营养空间,林下草本物种多样性比杉木纯林较高[26]。
林下植物群落物种多样性驱动因素的识别是森林生态学的一个重大挑战[27]。尽管大量的科学研究表明林分空间结构对林下草本物种多样性有显著的影响作用[28],但林分空间结构因子众多,再加上森林是一个复杂的生态系统,影响林下草本物种多样性的主导空间结构因子并不明确。本研究表明根据林分空间结构因子与林下草本物种多样性关联度系数排序结果可知:林分混交度是影响3种林分林下草本物种多样性的共同林分空间结构因子,这与朱光玉等[11]的研究结果一致。但3种林分林下草本物种多样性与混交度的关联系数并不是非常高,这说明林分混交度只能解释林下草本物种多样性的部分变异,相对较低的解释力表明,各种不可测量的生物和非生物因素、随机事件也可能对林下草本物种多样性产生重要影响[29]。因此,为了更深入地了解林分空间结构与林下草本物种多样性的关系,可以尝试构建非线性回归模型来进一步探讨。
4.2 结论
1)研究区天然次生林的空间结构整体上最优,杉木人工混交林次之,杉木人工纯林最差。因此,可以将天然次生林的空间结构作为模板,采取补植阔叶树种或乡土树种、人工促进天然更新、调整林分各树种的配置等人为干扰措施来优化杉木人工林空间结构,是其趋向近自然森林的林分空间结构特征。
2)研究区天然次生林、杉木混交林和杉木纯林的林下草本物种丰富度、Simpson多样性指数、Shannon-Wiener多样性指数、Simpson均匀性指数和Pielou均匀度指数同样都表现出了天然次生林 >杉木混交林 > 杉木纯林的规律。这在一定程度上说明林分空间结构是影响林下草本物种多样性的一个重要因素。
3)根据林分空间结构因子与林下草本物种多样性关联度系数排序分析可知:林分混交度是影响3种林分林下草本物种多样性的共同林分空间结构因子,因此,欲提高杉木纯林和混交林的林下草本物种多样性,可选择通过改善树种结构的人为干扰措施来实现。
猜你喜欢
现代农村科技(2022年5期)2022-11-18
青年文学家(2021年16期)2021-08-05
少儿美术(2020年8期)2020-11-06
共产党员·下(2018年6期)2018-12-05
现代园艺(2017年13期)2018-01-19
现代农业研究(2017年11期)2018-01-12
方圆(2018年23期)2018-01-07
现代园艺(2017年20期)2017-02-03
农业与技术(2016年6期)2016-05-14
小说月刊(2014年1期)2014-04-23
