
林窗大小对2种针叶林更新效果的初步分析
2019-10-31 09:07:34王敏彪杜有新尤根彪王军峰练发良何小勇
生态与农村环境学报 2019年10期
刘 伟,王敏彪,杜有新,尤根彪,王军峰,练发良,何小勇②
(1.丽水市白云山生态林场,浙江 丽水 323000;2.丽水市林业科学研究院,浙江 丽水 323000)
人工针叶林普遍存在生态服务功能退化的现象,如何优化林分结构及提高生态服务功能水平,已成为当前恢复生态学研究的重要课题之一[1-2]。通过更新采伐获得不同大小的林窗,是促进人工林更新、近自然培育及生态服务功能提升的重要干扰措施之一[3-5]。林窗内光照条件的改善和光照时间的延长导致水热条件发生相应的变化,直接或间接影响着植被群落的物种组成、群落结构和生态服务功能[6-7]。在影响林窗更新效应的诸多因素中,林窗面积起到至关重要的作用,是造成林内和林窗之间物种丰富度差异的主要原因[8]。研究林窗大小对森林更新的影响,对深入了解人工林改造及生态恢复具有重要的现实意义。
人工针叶林阔叶化改造的最终目的是培育较稳定健康的混交林结构,更新采伐林窗对森林结构的发展及生境条件的改善发挥着重要作用[9]。据报道,中大林窗的光照强度约为小林窗的2.8倍,小林窗又约为林下的3倍,林窗中灌木种类明显增多,灌木层和草本层盖度随着林窗的增大而增大[10]。采伐林窗能否对低效人工林生态系统产生持续的积极效应以及采伐面积的适宜性等问题仍无明确结论。该研究选取48年生马尾松(Pinusmassoniana)林和43年生杉木(Cunninghamialanceolata)林为对象,通过人工采伐获得不同大小的林窗,分析比较2种林分林窗间更新树种生长及天然更新植物多样性的差异,探讨人工更新的优良树种及较为适宜的林窗面积,为促进相似地区人工针叶林阔叶化改造及其可持续经营提供科学参考。
1 研究区域与研究方法
1.1 研究区概况
浙江省丽水市白云山生态林场(28°23′~28°27′ N, 119°52′~119°58′ E)属仙霞岭东支的括苍山脉,海拔为51.2~1 073.2 m,属于中亚热带季风气候区,气候温暖潮湿,雨量充沛,四季分明,年均气温为18.1 ℃,极端最高气温为43.5 ℃,极端最低温度为-7.7 ℃,年均降水量为1 392.8 mm,年均相对湿度为75.8%[11]。白云山属地质新构造运动强烈抬升而形成的山体,地表岩相以侏罗纪凝灰岩为主,白垩纪砂岩及花岗岩局部分布。土壤属中亚热带山地红黄壤,红壤主要分布于海拔600 m以下的低山丘陵,黄壤分布在600 m以上的山地。
1.2 样地设置
试验地位于白云山生态林场林相改造试验区,没有其他人为及农户放牧等干扰。2种林分基本概况见表1。
表1 2种林分基本概况
Table 1 Conditions ofPinusmassonianaplantations andCunninghamialanceolataplantations

林分类型林龄/a林分密度/(株·hm-2)郁闭度灌木盖度/%pH值土壤厚度/cmw(有机碳)/(mg·g-1)海拔/m坡度/(°)坡向 马尾松林488000.8±0.280.5±3.34.4±0.215.3±7.310.84±2.4141328东南 杉木林 431 2000.9±0.125.4±2.54.6±0.146.1±4.830.97±2.2835526 南
2015年12月对丽水市白云山生态林场马尾松林和杉木林的实地调查发现,自然形成的林窗多为近椭圆形,面积介于4~500 m2之间[12]。研究采用随机布点和典型抽样相结合的方法,在每种林分内采伐3种大小不等的近圆形林窗,分别采伐500 m2大林窗(G1)2个、250 m2中林窗(G2)2个和50 m2小林窗(G3)10个,各林窗边界即距林缘>10 m,以林下为对照(CK),2种林分通过采伐共形成林窗28个。砍伐的马尾松和杉木的树干及枝条等全部移出林窗,原林下灌草层植物连根清除,但马尾松和杉木树桩未掘出,林窗内植物更新主要靠种子及土壤种子库,杉木的萌蘖更新仅占极少数。
1.3 试验及调查方法
2016年3月,在采伐获得的林窗内,按2 m×3 m的株行距分别种植10个更新树种的2年生实生苗(表2),10个树种分别为枫香(Liquidambarformosana)、南方红豆杉(Taxuschinensisvar.mairei)、红豆树(Ormosiahosiei)、闽楠(Phoebebournei)、木荷(Schimasuperba)、朴树(Celtissinensis)、山乌桕(Sapiumdiscolor)、无患子(Sapindusmukorossi)、细柄蕈树(Altingiagracilipes)和浙江楠(Phoebechekiangensis)。G3林窗内每种种植1株,G2林窗内每种随机种植4株,G1林窗内每种随机种植8株。于2018年11月调查所有目标树种的地径和树高。
表2 10个树种的基本情况
Table 2 Conditions of ten tree species
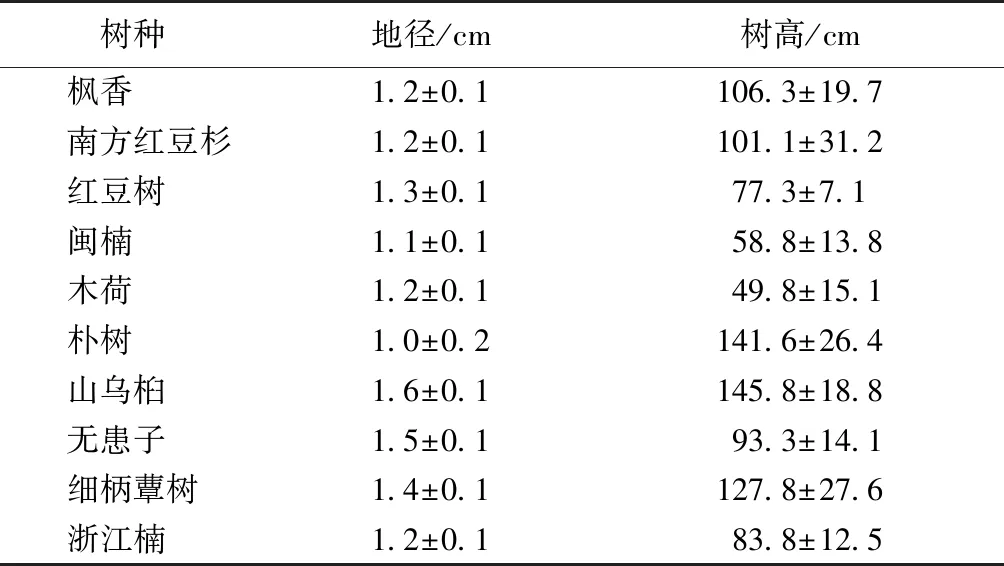
树种地径/cm树高/cm 枫香1.2±0.1106.3±19.7 南方红豆杉1.2±0.1101.1±31.2 红豆树1.3±0.177.3±7.1 闽楠1.1±0.158.8±13.8 木荷1.2±0.149.8±15.1 朴树1.0±0.2141.6±26.4 山乌桕1.6±0.1145.8±18.8 无患子1.5±0.193.3±14.1 细柄蕈树1.4±0.1127.8±27.6 浙江楠1.2±0.183.8±12.5
2017年9月,在每种规格的林窗及对照分别设置1~5个4 m×5 m的灌木样方,以林窗中心为起点,沿东西和南北方向每隔2 m平行设置1个样方;在距林窗边缘10 m之外的林下以相同方式设置5个林下对照样方。在每个大林窗内设置5个样方,中林窗内南北设置3个样方,小林窗中央设置1个样方,对照设置5个样方,每种林分分别设置灌木样方各31个。调查并记录样方内灌木的种类、株数、高度和冠幅,统计样方内草本植物的种类、株数或丛数、盖度等。
1.4 数据处理
统计林窗内和对照林下的物种数数目(S),计算物种重要值:物种重要值=(相对多度+相对冠幅或相对盖度+相对频度)/3。根据重要值求出Shannon物种多样性指数(H)、Simpson生态优势度(D)及Pielou均匀度指数(J)[13-14]:
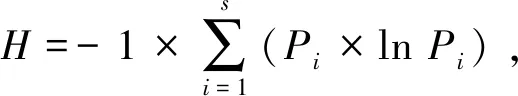
(1)
(2)
Pi=Ni/N,
(3)
J=H/lnS。
(4)
式(1)~(4)中,Ni为物种i的重要值;N为样地中所有物种的重要值之和;S为物种数。
植物群落Jaccard相似系数(CR)根据如下公式[15]计算:
CR=2a/(b+c)。
(5)
式(5)中,a为2个样地共有物种数;b为甲样地总种数;c为乙样地总种数。
采用Excel 2010和SPSS 20.0软件对数据进行统计分析,图表中数据为平均值±标准差。采用单因素方差分析(one-way ANOVA)和最小显著差异法(LSD)进行方差分析和多重比较(α=0.05),采用双因素方差分析(two-way ANOVA)检验林分和林窗大小及其交互作用对更新树种生长的影响。
2 结果与分析
2.1 人工更新树种的生长表现
10个树种在2种林分和不同大小林窗间的生长表现存在一定差异(图1)。比较树种生长量及成活情况,10个树种在杉木林中总体上优于马尾松林。在马尾松林中,枫香、无患子、山乌桕和细柄蕈树比其他树种生长表现好,山乌桕是大林窗内生长量最大的树种,但其在中小林窗未见成活。在杉木林中,枫香、山乌桕、南方红豆杉、无患子和细柄蕈树比其他树种生长情况好。闽楠和浙江楠在2种林分及不同大小林窗内均能适应。通过分析10个更新树种的地径和树高,发现马尾松林中地径大于3.5 cm的有山乌桕、枫香、细柄蕈树和无患子,树高超过2 m的有山乌桕、枫香、细柄蕈树、无患子和木荷。杉木林中地径大于2.5 cm的有细柄蕈树、南方红豆杉、山乌桕、红豆树、木荷、无患子、枫香和浙江楠,树高在2.5 m以上的有山乌桕、无患子、木荷、红豆树、枫香、浙江楠、阿丁枫和南方红豆杉。
在马尾松林中,随着林窗面积减小,枫香和无患子的地径表现为逐渐降低,闽楠和浙江楠规律不明显,其他树种在中小林窗表现不佳。马尾松林大林窗长势较好的有枫香、木荷、山乌桕和细柄蕈树,大中林窗没有差异的有无患子、闽楠和浙江楠,红豆树、南方红豆杉和朴树长势均不佳。在杉木林林窗中,随着林窗面积减小,枫香和山乌桕的地径和树高均表现为逐渐增大,红豆树、木荷、无患子和细柄蕈树在中等林窗较差,其他树种没有差异。杉木林大林窗长势较好的有红豆树,小林窗长势较好的有枫香、山乌桕和浙江楠,大小林窗均优于中林窗的有南方红豆杉、闽楠、木荷、无患子、细柄蕈树,朴树中等林窗长势较好。综合目标更新树种早期生长表现分析,马尾松林阔叶化改造可采取中等林窗,而杉木林改造采取大或小林窗较好。
2.2 林分和林窗对人工更新树种生长的影响
由表3可知,林分类型和林窗面积对不同树种生长的影响存在差异。
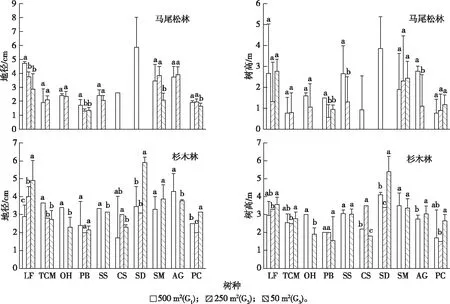
LF—枫香;TCM—南方红豆杉;OH—红豆树;PB—闽楠;SS—木荷; CS—朴树;SD—山乌桕;SM—无患子;AG—细柄蕈树;PC—浙江楠。 同一幅图中同一组直方柱上方英文小写字母不同表示不同林窗间某树种地径或树高差异显著(P<0.05)。图1 10个人工更新树种在马尾松林和杉木林中的生长表现Fig.1 Growth of 10 tree species in Pinus massoniana and Cunninghamia lanceolata plantations
表3 林分类型和林窗大小对10个更新树种生长影响的双因素分析
Table 3 Two-way ANOVA of effects of forest types and gap size on the growth of 10 tree species
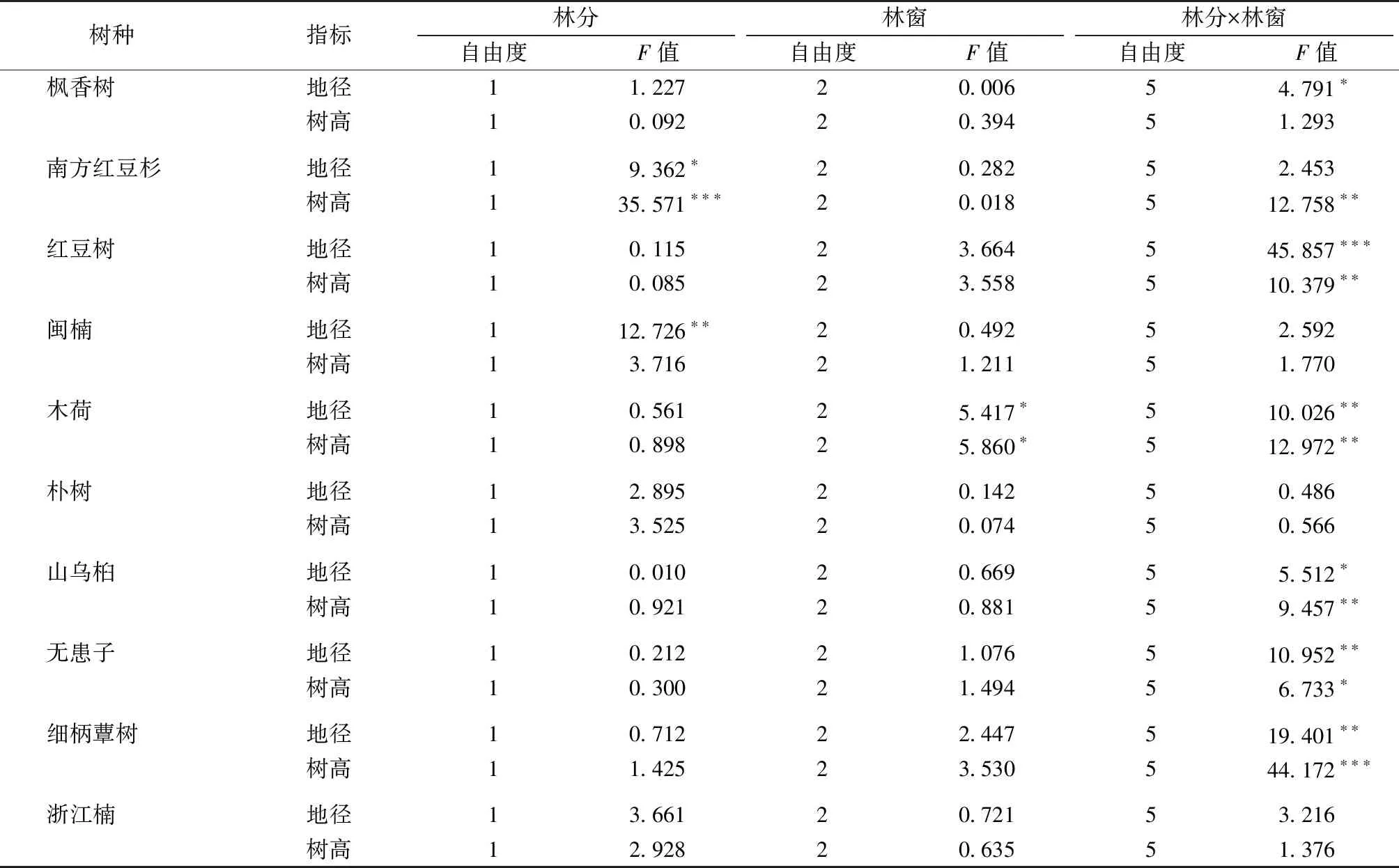
树种 指标林分林窗林分×林窗自由度F值自由度F值自由度F值 枫香树地径11.22720.00654.791∗树高10.09220.39451.293 南方红豆杉地径19.362∗20.28252.453树高135.571∗∗∗20.018512.758∗∗红豆树地径10.11523.664545.857∗∗∗树高10.08523.558510.379∗∗闽楠地径112.726∗∗20.49252.592树高13.71621.21151.770 木荷地径10.56125.417∗510.026∗∗树高10.89825.860∗512.972∗∗朴树地径12.89520.14250.486树高13.52520.07450.566 山乌桕地径10.01020.66955.512∗树高10.92120.88159.457∗∗无患子地径10.21221.076510.952∗∗树高10.30021.49456.733∗ 细柄蕈树地径10.71222.447519.401∗∗树高11.42523.530544.172∗∗∗浙江楠地径13.66120.72153.216树高12.92820.63551.376
*表示P<0.05;**表示P<0.01;***表示P<0.001。
林分类型显著影响南方红豆杉生长,特别是对树高的影响极为显著(P<0.001),对闽楠地径生长影响显著(P<0.01)。林窗大小只对木荷地径和树高有显著影响(P<0.05)。林分和林窗的交互作用对细柄蕈树、木荷、山乌桕、无患子和红豆树地径和树高均有显著或极显著影响,对枫香树的地径和南方红豆杉的树高具有显著影响(P<0.05或P<0.01)。林分类型和林窗大小对朴树、闽楠和浙江楠均没有影响。
2.3 天然更新植物的多样性特征
由表4可知,天然更新植物的物种数与Shannon多样性指数具有类似变化特征。马尾松对照林下物种数和Shannon多样性指数显著高于小林窗(P<0.05),与中大林窗没有差异;而杉木林对照林物种数和Shannon多样性指数显著低于其余各个林窗(P<0.05)。马尾松林窗内与其对照林下Simpson生态优势度没有显著差异,而杉木林大林窗和小林窗则显著高于对照(P<0.05)。从天然更新植物多样性来看,开辟林窗对马尾松林效果不大,而对杉木林改造效果明显,采伐小林窗即可有效丰富林下植物种类。
表4 2种林分林窗间物种多样性指数
Table 4 Diversity index of species among different size gaps in the two plantations

林窗马尾松林杉木林物种数Shannon指数Simpson优势度Pielou指数物种数Shannon指数Simpson优势度Pielou指数 G118.0±5.3a2.79±0.19a0.93±0.06a0.96±0.33a16.5±6.4a2.62±0.24a0.91±0.02a0.94±0.02a G216.0±3.7a2.69±0.26a0.93±0.14a0.97±0.22a17.0±4.2a2.21±0.55a0.74±0.15ab0.77±0.16ab G39.0±1.6b2.06±0.21b0.85±0.08a0.94±0.16a20.0±5.7a2.64±0.08a0.90±0.03a0.89±0.07a CK19.0±4.4a2.75±0.33a0.92±0.07a0.95±0.29a9.5±0.7b1.48±0.18b0.68±0.04b0.67±0.11b
G1、G2、G3林窗面积分别为500、250和50 m2。同一列数据后英文小写字母不同表示不同林窗间某指标差异显著(P<0.05)。
不同林分之间物种相似性系数的比较(表5)表明,马尾松林中等林窗与杉木林不同大小林窗间天然更新植物具有较高的相似度,均高于0.46。2种林分对照与其他林分林窗之间物种相似度大多低于0.3。相同林分不同林窗之间物种相似度高于其与对照之间,也明显高于不同林分林窗之间;相同林分不同大小林窗与不同林分对照之间的物种相似度均较低。2种林地均有出现的天然更新树种有格药柃(Euryamuricata)、白背叶(Mallotusapelta)、盐肤木(Rhuschinensis)、檫木(Sassafrastzumu)、檵木(Loropetalumchinense)和山鸡椒(Litseacubeba);马尾松林中单独出现的为耐瘠薄的阳性树种,如山油麻(Tremadielsiana)、枫香、白栎(Quercusfabri)、赤楠(Syzygiumbuxifolium)、华山矾(Symplocoschinensis)、赛山梅(Styraxconfusus)、毛柄连蕊茶(Camelliafraterna)及乌药(Linderaaggregata);杉木林中单独出现的多为喜深厚肥沃土壤或耐阴湿的树种,如臭辣树(Evodiafargesii)、南酸枣(Choerospondiasaxillaris)、杨梅(Myricarubra)、苦楝(Meliaazedarach)、浙江柿(Diospyrosglaucifolia)、赤杨叶(Alniphyllumfortunei)、白棠子(Callicarpabodinieri)和硃砂根(Ardisiacrenata)。说明除采伐林窗改善林地光照条件及其引起的空气温湿度变化这一因素外,2种林地土壤条件差异也影响着不同更新植物的定居和生长,具体原因有待进一步研究。
表5 2种林分林窗间物种相似性系数
Table 5 Semi-matrix of resembling coefficients among different forest gaps in the two plantations

林分类型林窗马尾松林杉木林 G2G3CKG1G2G3CK 马尾松林G10.4000.8000.2860.2940.2930.2700.174G20.5330.2140.5000.4670.5160.316G30.3160.1330.1940.1710.133CK0.0450.2860.0890.211 杉木林 G10.8670.8480.500G20.9090.357G30.429
G1、G2、G3林窗面积分别为500、250和50 m2。
3 讨论与结论
3.1 人工更新树种生长分析
不同林分土壤环境存在差异,林窗干扰导致光照强度、空气温湿度以及土壤理化性质等环境条件发生变化[16],这都会影响更新植物的生长。在杉木林中的长势优于马尾松林的更新树种有南方红豆杉和闽楠,这与2个树种较耐阴和喜阴湿肥沃的生境条件有关[17-19],在幼树阶段,杉木林下的弱光环境更适宜南方红豆杉和闽楠的生长[20-21],其他树种在2种林分中差异不大,朴树长势均表现不佳。山乌桕在马尾松中小林窗死亡,而在杉木林小林窗中长势优于大中林窗,这也许与山乌桕既耐瘠薄又喜湿润的环境有关[22]。枫香的地径在马尾松林中随林窗减小逐渐下降,而在杉木林中则逐渐增加,枫香在2种林分中的生长表现均较好,表明枫香适应性较强,可作为亚热带山区林相改造的主要树种之一[23]。木荷、红豆树和细柄蕈树为该地区的乡土树种,其中木荷是耐瘠薄的先锋树种[24],而红豆树[25-26]和细柄蕈树[27]为中性树种,但它们在马尾松林小林窗和杉木林中等林窗内均出现枯死现象,其原因也许与马尾松林的优势植物山油麻和杉木林下的芒萁(Dicranopterisdichotoma)对资源空间的激烈竞争有关,这与反映资源空间利用能力的种群生态位密切相关[28],有待进一步研究。无患子的地径在马尾松林中随着林窗面积减小表现为逐渐降低现象,而在杉木林各面积林窗中没有差异,也许是因为无患子对光强的适应范围广[29],也较喜欢肥沃的土壤环境[30]。
林分类型及林分和林窗的交互作用显著影响南方红豆杉生长,特别是对其树高的影响极为显著,这与南方红豆杉偏喜阴和肥沃土壤的生物生态学特性有关[20]。林分类型对闽楠地径具有显著影响,其在杉木林中的生长表现显著优于马尾松林,但林窗大小及林分和林窗的交互作用对其没有影响,说明闽楠幼树较为适应耐阴及土壤肥沃的微生境[21, 31]。虽然林分类型和林窗大小对细柄蕈树、木荷、山乌桕、无患子和红豆树的地径和树高没有影响,但林分和林窗的交互作用对它们均有显著或极显著影响,对枫香树的地径和南方红豆杉的树高也具有显著影响。这也许与林窗干扰改变了光照和土壤等微生境条件有关[16],林窗内与林下的物种相似度很低也说明了这种情况。
3.2 天然更新植物多样性分析
不同林分Shannon多样性指数和Simpson生态优势度不仅能反映物种数量,而且能较好地反映物种的空间分布格局[32]。研究结果表明,林分类型及林窗大小对杉木林物种多样性的影响较为显著。2种林分的中大林窗中Shannon多样性指数和Simpson生态优势度均无显著差异,小林窗物种数和Shannon多样性指数均为杉木林>马尾松,对照林下则为马尾松>杉木林,反映马尾松林下生境异质性较高[33]。随着林窗增大,物种数和Shannon多样性指数在马尾松林表现为大林窗=中林窗=对照>小林窗,杉木林为大林窗=中林窗=小林窗>对照,这2种林分的大中林窗之间没有差异,这也许与林分类型及土壤环境均质性较高有关[34]。
林窗干扰导致光照条件变化,这也是引起其他生态因子发生变化的重要原因,如林窗土壤速效态养分含量高于林内土壤[10],这些变化为林窗内灌草更新植物的入侵和生长创造了有别于林下的环境条件[4]。杉木林下的物种多样性显著低于林窗内,而马尾松林下生长的矩叶鼠刺(Iteaoblonga)、老鼠矢(Symplocosstellaris)和马银花(Rhododendronovatum)在其林窗内均没有出现。由于不同林分土壤条件存在差异,马尾松林不同大小林窗间物种相似度显著低于杉木林,说明马尾松林窗内不同树种间对水分和养分等的竞争更为激烈,这也许是林分类型和林窗面积的综合作用比单个因素对更新植物生存产生的影响更为显著的主要原因[35]。相同林分不同林窗之间物种相似度高于其与对照之间,也明显高于与其他林分林窗之间,不同大小林窗与其他林分对照之间的物种相似度均较低,充分说明林窗微环境与林窗边缘的叠加效应丰富了植物可利用的资源状况[36]。
总体来说,杉木林土层厚度和土壤有机质含量均高于马尾松林,而且天然更新的灌木层植物的竞争较小,在光照条件得到改善后,更新树种在杉木林中的长势优于马尾松林。根据目标更新树种早期生长状况,枫香、无患子、山乌桕和细柄蕈树表现较好,山乌桕生长量最大,木荷、红豆树、闽楠和浙江楠在2种林分中的表现一般,朴树表现最差。马尾松林改造以中等林窗为主,可选择的树种有枫香、无患子、细柄蕈树和木荷;杉木林改造以大或小林窗为主,可选择的树种有枫香、山乌桕、浙江楠、南方红豆杉、无患子、细柄蕈树、闽楠和木荷。林窗大小对林下植物多样性的影响不仅与林窗内气候条件有关,还与不同林地土壤环境及林窗发育阶段有关[34],今后应重点关注林窗内植物多样性水平随着林窗发育而呈现的变化规律。由于该研究所采伐的林窗面积梯度有限,具体采伐多大的林窗更有利于针叶纯林更新改造,还需进一步探讨。
猜你喜欢
大自然探索(2023年11期)2023-03-01 09:05:04
西南林业大学学报(2022年5期)2022-10-17 09:56:38
幼儿教育·父母孩子版(2022年8期)2022-05-30 18:27:54
林业与生态(2022年5期)2022-05-23 01:16:51
花卉·上半月(2022年9期)2022-04-29 00:44:03
农业机械学报(2020年1期)2020-03-13 00:27:38
山东林业科技(2019年2期)2019-06-03 10:10:54
天然产物研究与开发(2018年7期)2018-08-21 02:04:12
西部林业科学(2018年4期)2018-02-14 22:43:50
现代园艺(2017年21期)2018-01-03 06:42:18
