
新疆托克逊县极端环境土壤螨类群落多样性研究
2019-10-25 01:27阿丽亚司地克吾玛尔阿布力孜排孜丽耶合力力
江苏农业科学 2019年13期
阿丽亚?司地克 吾玛尔?阿布力孜 排孜丽耶?合力力
摘要:为了解干旱区极端环境下土壤螨类群落结构和多样性特征,采用形态特征分类和生态学方法对新疆托克逊县8种不同生境土壤螨类群落结构和多样性特征进行调查研究。结果表明,共捕获15 420头土壤螨类标本,隶属于蜱螨亚纲的有4亚目49科61属,其中长单翼甲螨属(Protoribates)为优势类群。不同生境土壤螨类个体数与类群数之间均有明显差异(P<0.05),其中个体数依次为 Ⅰ>Ⅱ>Ⅲ>Ⅵ>Ⅶ>Ⅴ>Ⅳ>Ⅷ,季节变化表现为秋季最高,夏季最少,垂直分布均呈现明显的表聚性。不同生境土壤螨类群落多样性指标之间存在显著差异(P<0.05),其中Shannon-Wiener多样性指数(H)在葡萄园和杏园内较高,而在灌木林和温室大棚中较低,其顺序依次为Ⅱ>Ⅲ>Ⅴ>Ⅳ>Ⅶ>Ⅰ>Ⅷ。Jaccard相似性系数表明,不同生境土壤螨类群落相似性较高,基本上属于中等相似(0.50≤q<0.75)。
关键词:干旱区;托克逊县;土壤螨类;多样性;季节变化
中图分类号: S154.6 文献标志码: A 文章编号:1002-1302(2019)13-0293-06
土壤螨类是陆地生态系统中重要的消费者和特殊的分解者,是具有种类多、分布广、种群密度大、生物多样性高、迁移性弱、敏感性强、食性复杂及营自由生活等特征的小型节肢动物[1],主要栖息于土壤表层和落叶层,取食菌类、藻类和腐烂的动植物残体,在土壤的形成、有机质的分解和提高肥力等方面具有重要的作用[2]。由于螨类种类组成、数量变化和生态分布特征与土壤类型和肥力、理化性质及人类的生产活动密切相关,并能反映土壤的细微变化,在土壤环境变化监测和生态毒理诊断等方面具有特殊的功能。因此,土壤螨类的研究一直受到国内外学者的高度重视和关注[2-4]。在国外,对土壤螨类的研究已有170多年的历史,而我国此类研究起步较晚,只有40多年的历史,但发展迅速。到目前为止,我国从热带、亚热带到温带,从森林、草地、沙漠到湿地、农田、城市等诸多生态系统都开展了较为详细的研究[5-6],但发展不均衡,沿海地区的研究开展得较早并广泛,而西北干旱区起步较晚且只有零星报道,尤其是有关干旱区特殊环境土壤螨类群落多样性方面的研究一直欠缺[7-11]。
托克逊县地处亚欧大陆腹地,远离海洋,三面环山,自然条件恶劣(干旱、高温、多风),属于极端干旱的暖温带内陆荒漠气候。由于该县在新疆具有重要的地缘优势,是古代丝绸之路的重要地段,是我国内地连接新疆、中亚地区的交通枢纽,也是南北疆及东疆之间的铁路和公路枢纽。自然和人为因素是该区生态环境变化的主要驱动因子,绿洲土壤的盐碱化、沙漠化和污染现象比较严重,生态环境处于脆弱状态[12-13]。全球气候变化、气候异常和区域人类活动等干扰对该地区生态系统结构和功能以及土壤螨类生态分布产生了深远的影响。本研究以托克逊县8种不同生境作为试验样地,旨在探讨极端干旱环境下土壤螨类群落多样性及其时空动态变化特征,为干旱区生态系统的保护和可持续发展提供科学依据。
1 材料与方法
1.1 研究区概况
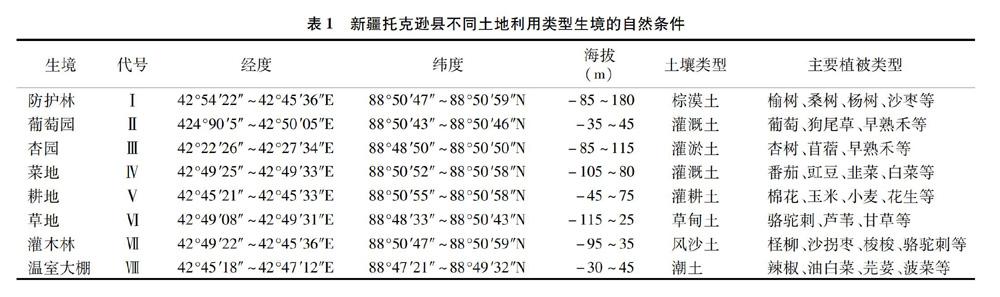
托克逊县位于新疆维吾尔自治区中部,天山南麓,吐鲁番盆地西部,东与吐鲁番市高昌区为邻,南与巴州尉犁县相接,西与巴州的和硕、和静县相连,北与乌鲁木齐市毗邻。地处87°14′05″E~89°11′08″E,41°21′14″N~43°18′11″N,南北长约204 km,东西宽146 km,平原绿洲海拔为-125~200 m,土地总面积为16 561 km2,包括8个乡(镇)。该区属于暖温带极端干旱荒漠气候,形成了日照长、气温高、昼夜温差大、降水少、风力强等5大特点。光热资源十分丰富,日照时数为 3 043.30 h,年均气温为13.8 ℃,极端最高温和最低温分别为48、-25.5 ℃。≥10 ℃积温为5 334.9 ℃,年均降水量为 6.3 mm,年均蒸发量达3 744 mm,无霜期为219 d[14]。该地区现有耕地、草地、林地、园地和未利用地等多种土地利用类型,自西向东分布着棕漠土、灌耕土、灌淤土、草甸土、潮土、盐土和风沙土,土壤肥力较低,主要农作物有春小麦、高粱、棉花、花生以及甜瓜、西瓜、杏、葡萄、蔬菜等[14]。本研究在托克逊县境内设2条长达50 km的样线,各样线分别选择防护林(Ⅰ)、葡萄园(Ⅱ)、杏园(Ⅲ)、菜地(Ⅳ)、耕地(Ⅴ)、草地(Ⅵ)、灌木林(Ⅶ)及温室大棚(Ⅷ)等8种土地利用类型生境,并用全球定位系统(GPS)测量各样地的地理位置、海拔等数据,各生境自然环境情况见表1。
1.2 样本采集
分别于2017年3、6、9、11月中旬在2条样带上选择8种生境,每个生境随机选取3个样地,每个样地按对角线法设5个采样点,并采用容量为100 cm3的环刀分4层(0~5、5~10、10~15、15~20 cm)取樣,共取576份土样,装入塑料自封袋后带回实验室用改进的Tullgren法分离收集土壤螨类。
1.3 分离与鉴定
标本处理详情见吾玛尔·阿布力孜的方法[15]。分类检索在Olympus SZX16体视显微镜和Leica DM4000B显微镜下参照尹文英等的《中国土壤动物检索图鉴》[16]、青木淳一的《日本土壤动物检索图鉴》[17]、江原昭三的《日本蜱螨类检索图鉴》[18]、Balogh等的《The Oribatid mites genera of the world》[19]及Krantz等的《A manual of acarology.》[20]等对土壤螨类进行分类鉴定和数量统计,一般鉴定到属级分类单元。
1.4 数据处理
不同生境下土壤螨类群落多样性分析采用以下公式计算:
式中:Pi为第i种类群个体数在群落总个体数中所占的比例;S为不同生境中的类群数;N为不同生境中的总个体数;ni为不同生境中的个体数;a为A群落类群数;b为B群落类群数;c为2个群落共有类群数;Xij和Xik分别为i类群在群落j、k中密度的4次方根。所有数据和分析采用SPSS 19.0软件进行,并在Excel 2007中作图。采用单因素方差分析(one-way anova)和最小显著差异法(least-significant difference,简称LSD)比较不同数据组间的差异。
2 结果与分析
2.1 土壤螨类群落组成和数量变化
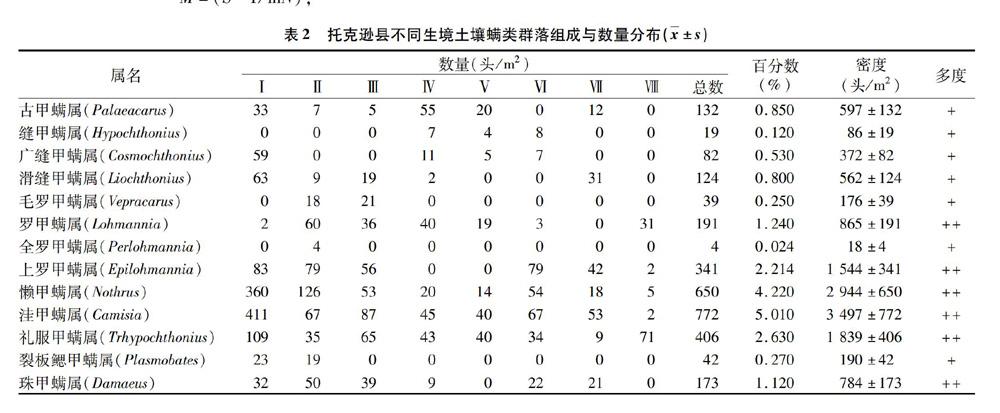
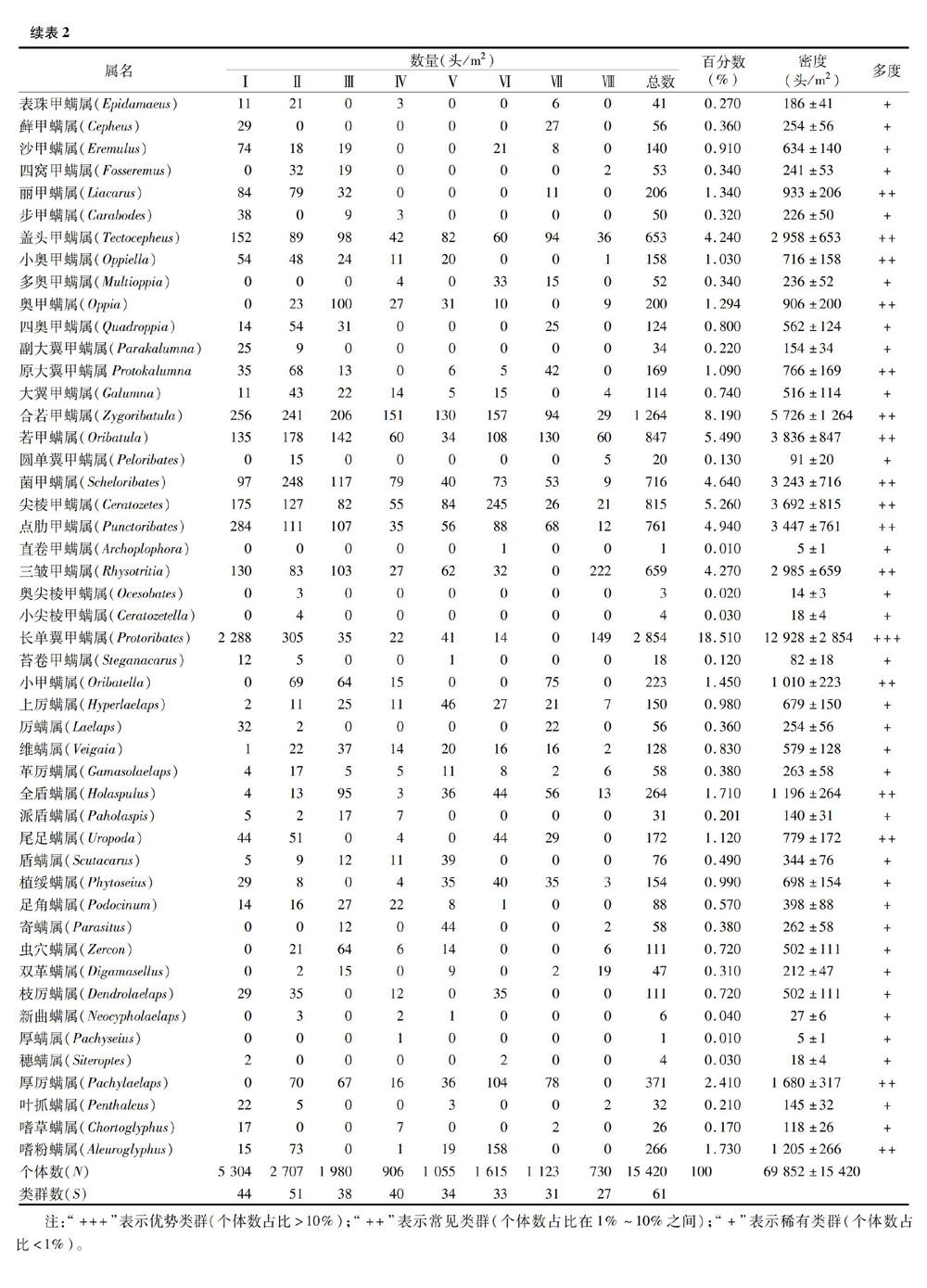
本研究共获得15 420头土壤螨类标本,隶属于蛛形纲蜱螨亚纲的有4亚目49科61属,其中甲螨亚目有13 210头,33科,40属,占总数量的85.67%;中气门亚目有150头,11科,15属,占总数量的9.73%;前气门亚目有408头,3科,4属,占总数量的2.65%;无气门亚目有292头,2科,2属,占总数量的 1.89%。对整个研究区来说,长单翼甲螨为优势类群,占总个体数的18.51%。懒甲螨属、洼甲螨属、盖头甲螨属、合若甲螨属、全盾螨属等22属为常见类群,占总个体数的66.65%。大翼甲螨属、广缝甲螨属、四窝甲螨属、步甲螨属、嗜草螨属等38属为稀有类群,占总数的14.85%(表2)。
方差分析结果表明,在8种生境土壤中的螨类个体数与类群数之间均有显著差异(P<0.05),其中个体数依次为Ⅰ>Ⅱ>Ⅲ>Ⅵ>Ⅶ>Ⅴ>Ⅳ>Ⅷ,而类群数依次为Ⅱ>Ⅰ>Ⅳ>Ⅲ>Ⅴ>Ⅵ>Ⅶ>Ⅷ。无论从个体数还是类群数来看,土壤螨类数量在防护林(Ⅰ)和葡萄园(Ⅱ)最多,而在菜地(Ⅳ)和温室大棚(Ⅷ)最少。其原因可能是葡萄园常用农家肥,而防护林有较厚的凋落物层,为螨类的生存繁衍提供了良好的生活环境。菜地(Ⅳ)和温室大棚(Ⅷ)生境虽然有一定的湿度和土壤肥力,而人为干扰较多,土壤成分单一,没有足够多的腐殖层。
2.2 土壤螨类群落的时空分布
在垂直分布上,8种生境土壤螨类个体数与类群数在不同土层之间均有明显差异,并呈现出明显的表聚性,研究区土壤螨类主要集中分布于表层土壤,即0~5 cm土层(8 248头,53.49%),其次为5~10 cm土层(4 088头,26.51%)、10~15 cm土层(2 204头,14.29%),而15~20 cm土层最少(880头,5.71%)(图1)。
从季节变化来看,不同生境土壤螨类个体数量均有明显差异,依次为秋季(7 838头,50.83%)>春季(4 683头,30.37%)>冬季(1 946头,12.62%)>夏季(953头6.18%),见图2。
2.3 土壤螨类群落多样性指数
群落多样性可以反映群落组成的复杂程度,用来评价群落生态的组织水平[22]。采用群落多样性指数(H)、丰富度指数(M)、均匀度指数(E)及优势度指数(C)对不同生境土壤螨类群落进行分析(图3)。
分析可知,8种生境土壤螨类群落多样性指标之间存在显著差异(P<0.05),其中Shannon-Wiener多样性指数(H)在葡萄园和杏园内较高,而在灌木林和温室大棚内较低,其顺序依次为Ⅱ>Ⅲ>Ⅴ>Ⅳ>Ⅶ>Ⅵ>Ⅰ>Ⅷ。Margalef丰富度指数(M)在葡萄园内最高,而在温室大棚内最低,其顺序依次为Ⅱ>Ⅳ>Ⅰ>Ⅲ>Ⅴ>Ⅵ>Ⅶ>Ⅷ。Simpson优势度指数(C)依次为Ⅰ>Ⅷ>Ⅵ>Ⅳ>Ⅶ>Ⅴ>Ⅱ>Ⅲ。Pielou均匀度指数(E)依次为 Ⅳ>Ⅴ>Ⅶ>Ⅲ>Ⅱ>Ⅵ>Ⅷ>Ⅰ。防护林Ⅰ土壤螨类优势度高,但多样性指数和丰富度指数并不高,均匀度指数较低,显示出优势度和均匀度呈负相关关系。
2.4 土壤螨类群落相似性分析
群落相似性分析是根据群落的物种组成与数量分布判断群落间的相似程度,分析影响群落结构的主要因素[21-22]。不同生境土壤螨类群落Jaccard相似性分析结果见表3。
由表3可知,不同生境间土壤螨类群落相似性较高,灌木林(Ⅶ)与防护林(Ⅰ)、菜地(Ⅳ)、耕地(Ⅴ)、草地(Ⅵ)和温室大棚(Ⅷ)间的相似性为中等不相似(0.25≤q<0.50),占总相似性比例的 21.43%。其他生境之间多数为中等相似(0.50≤q<0.75),占总相似性比例的78.57%。葡萄园(Ⅱ)和防护林(Ⅰ)之间的相似性系数最高(0.696),防护林(Ⅰ)和温室大棚(Ⅷ)之间的相似性系数最低(0.368),表明该区不同生境土壤螨类群落相似性基本上属于中等相似。
2.5 土壤螨类群落聚类分析
生态学研究中常采用聚类和排序分析方法,对不同生境生物群落进行比较。以8种生境土壤螨类个体数的4次方为原始数据进行欧式距离组间平均聚类分析(hierarchical cluster analysis,简称HCA)和多维尺度分析(multi dimensional scaling,简称MDS)[23](Stress值为0.053 10<0.2,说明图形吻合一般;RSQ值为0.976 69,说明不相似)。聚类和排序结果将8种生境分为4大类型,葡萄园(Ⅱ)和杏园(Ⅲ)合为1组,地表经常有凋落层,人为干扰较少并且土壤环境条件也相似,这些因素可能是这2种生境土壤螨类分为1组的原因;菜地(Ⅳ)和耕地(Ⅴ)合为1组,这可能是地表凋落层很少并且人为干扰较大;草地(Ⅵ)、灌木林(Ⅶ)和温室大棚(Ⅷ)为1组,这可能与地表凋落层较少和人为干扰较小有关,温室大棚人为干扰大并且土壤条件单一;防护林(Ⅰ)单独为1组,这可能是由于地表凋落层较厚并且人为干扰较少(图4、图5)。
3 结论与讨论
土壤螨类在土壤生态系统物质和能量循环中起着重要作用,土壤螨类可以通过自身行为活动改变土壤理化性质,同时土壤理化性质的改变可以引起土壤螨类群落结构及其多样性的变化[21-22]。研究表明,新疆托克逊县地区土壤螨类资源比较丰富,8种生境共获得15 420头土壤螨类标本,隶属于4亚目49科61属。其中优势类群和常见类群构成该地区土壤螨类群落的主体(85.16%),决定整个群落的特征和生态功能,而稀有类群比较多(38属),占总捕获量的14.85%,它们对环境因素有较高的敏感性,可以作为土壤环境的指示生物。研究区优势类群只有长单翼甲螨属,而鄯善县甲螨群落研究中的优势类群为尖棱甲螨属、菌甲螨属、合若甲螨属、若甲螨属,比托克逊县更加突出,这是因为鄯善县年均降水量高于托克逊县,土壤湿度高,适合更多甲螨生存[22]。因为湿度是影响土壤螨类密度和群落结构最重要的因素之一[23-25]。时空分布表明,从不同地层土壤螨类的垂直分布情况来看,0~5 cm 土层中土壤螨类的个体数和类群数最多,而其他土层较少,这与其他研究结果基本一致[5-13]。这是由于植被组成和土壤养分含量差异较大,表层土壤有机质含量比较丰富,容重低,通氣性好,土壤温湿度等条件都比较适合多种土壤螨类的生存与繁殖[23-26]。土壤动物的垂直分布差异与土壤的理化性质(土壤含水量、pH值、地温条件)、植被以及营养状况的垂直分布差异密切相关[24,27]。土壤螨类各类群个体数的季节动态变化显示,秋季最高,夏季最低,这与前人研究结果[10-11]基本一致。这可能是由于托克逊县夏季气温较高,土壤表面水分蒸发量大,从而使土壤湿度大大减少,不利于土壤螨类生存。
生物多样性是群落生物组成结构的重要指标,反映群落内物种的多少和生态系统食物网的复杂程度,从而反映各生境间的相似性和差异性[26]。研究表明,在不同土地利用类型土壤中,螨类群落多样性均有显著差异,这种差异可能与土壤环境的利用方式、土壤条件、植被类型以及凋落物层等多种因素有关[22-26]。结果表明,防护林和葡萄园多样性及丰富度均最高,且群落稳定性较好,防护林的优势度最高,这可能与凋落物含量较多和人为干扰较少有关。温室大棚螨类群落多样性和丰富度最低,这可能与凋落物很少并且人为干扰程度较大有关。防护林土壤螨类优势度最高,但均匀度指数最低,显示出优势度和均匀度呈负相关关系。分析表明,土壤螨类群落个体数越丰富,其多样性就越高,同时多样性指数也受均匀度指数的影响;群落内物种分布越均匀,则优势度越小,反之群落内物种分布不均匀,则优势度就越大[27]。相似性指数是衡量生境与土壤动物关系的重要指标[28],研究区8种生境整体上表现为“中等相似”。虽然地表植被有差异,但土壤螨类群落相似性仍然较高,说明该区影响土壤螨类生态分布的重要因素可能是土壤环境因子(土壤含水量、温度、有机质、pH值及容重)[28-29]。研究表明,该区域土壤螨类资源比较丰富,并且土壤螨类群落多样性指标在不同生境间均存在显著差异。由于土壤螨类群落结构与土壤理化性质密切相关,有待进一步研究来揭示不同生境环境因子与螨类群落之间的相关性。
参考文献:
[1]王东昌,杨振玲,张乃琴,等. 我国土壤螨研究现状进展[J]. 莱阳农学院学报,2001,18(1):61-65.
[2]尹文英,张荣祖,王世彰,等. 中国土壤动物[M]. 北京:科学出版社,2000:4-5.
[3]Fischer B M,Schatz H. Biodiversity of oribatid mites (Acari:Oribatida) along an altitudinal gradient in the central Alps[J]. Zootaxa,2013,3626(4):429-454.
[4]Bedano J C,Domínguez A,Arolfo R . Assessment of soil biological degradation using mesofauna[J]. Soil and Tillage Research,2011,117:55-60.
[5]吴东辉,尹文英,杨振明. 松嫩草原中度退化草地不同植被恢复方式下土壤螨类群落特征的差异[J]. 动物学报,2007,53(4):607-615.
[6]吾玛尔·阿布力孜,古丽布斯坦·努尔买买提,阿布都肉苏力·吐孙,等. 新疆阜康绿洲不同生境土壤动物群落多样性及其季节动态[J]. 生态学杂志,2013,32(6):1412-1420.
[7]陈 铮,刘大林,邓 波,等. 华北农牧交错带不同土地利用方式下土壤螨的分布特征[J]. 草原与草坪,2010,30(3):52-56.
[8]吾玛尔·阿布力孜,阿布都如苏力·吐逊,木卡热木·阿吉木,等. 乌鲁木齐地区不同生境土壤捕食性螨类群落结构及其多样性研究[J]. 新疆农业科学,2012,49(9):1748-1758.
[9]丁程成,戴征凯,薛晓峰,等. 退化红壤植被恢复对土壤螨类群落结构的影响[J]. 生态学报,2008,28(10):4771-4781.
[10]吴东辉,尹文英,陈 鹏. 刈割条件下松嫩平原碱化羊草草地土壤螨类群落变化特征的研究[J]. 土壤学报,2008,45(5):1007-1014.
[11]韩雪梅,李丹丹,梁子安,等. 北方常见农业土地利用方式对土壤螨群落结构的影响[J]. 生态学报,2013,33(16):5026-5034.
[12]郑经鸿,王丽真,张兴亚,等. 新疆草地甲螨分布规律研究[J]. 生态学报,1993,13(2):171-176.
[13]孜比尔妮莎·吾布力,阿尔孜古力·莎塔尔,吾玛尔·阿布力孜. 乌鲁木齐不同生境土壤甲螨群落特征初步研究[J]. 新疆农业科学,2010,47(4):832-841.
[14]赵海晶,高敏华,张 丽. 县级土地利用总体规划实施评价研究——以新疆托克逊县为例[J]. 中国农学通报,2015,31(19):191-198.
[15]吾玛尔·阿布力孜. 土壤螨类的采集与玻片标本的制作[J]. 生物学通报,2012,47(1):57-59.
[16]尹文英,胡圣豪,沈韫芬,等. 中国土壤动物检索图鉴[M]. 北京:科学出版社,1998:163-242.
[17]青木淳一. 日本土壤动物分类检索图鉴[M]. 东京:东海大学出版社,1999:173-436.
[18]江原昭三. 日本蜱螨类图鉴[M]. 东京:全国农村教育社会出版社,1980:1-562.
[19]Balogh J,Balogh P. The Oribatid mites genera of the world(Vol.1 & Vol.2)[M]. Budapest:Thehungarian National Museum Press,1992:1-263.
[20]Krantz G W,Walter D E. A manual of acarology[M]. 3rd ed ,Texas:Texas Tech University Press,2009:1-806.
[21]Dri J N,André H M. Soil mite diversity from Ivory Coast[J]. European Journal of Scientific Research,2011,64(2):263-276.
[22]排孜丽耶·合力力,吾玛尔·阿布力孜,阿加尔·恰肯. 干旱区不同土地利用类型下土壤螨类群落多样性比较[J]. 新疆农业科学,2017,54(12):2267-2281.
[23]阿加尔·恰肯,吾玛尔·阿布力孜,排孜力耶·合力力. 新疆玛纳斯河流域土壤螨类群落多样性[J]. 生态学报,2017,37(24):8385-8396.
[24]Ndri J K,Andre H M,Lagerlof J A,et al. Response of soil mite abundance and diversity to a monospecific timber Tectona grandis plantation in Ivory Coast[J]. Current Zoology,2013,59(5):633-643.
[25]阿提坎·吾布力喀斯木,吾玛尔·阿布力孜,努尔合尼穆·艾麦提. 玛纳斯河流域平原区土壤螨类群落特征研究[J]. 安徽农业科学,2014,42(23):7829-7834.
[26]阿布都如苏力·吐孙,吾玛尔·阿布力孜,木开热木·阿吉木. 准噶尔盆地东南缘绿洲-荒漠交错带土壤螨类初步研究[J]. 干旱区资源与环境,2013,27(3):160-166.
[27]李钰飞,郑春燕,李吉进,等. 不同管理模式对温室土壤螨群落结构的影响[J]. 生态学报,2016,36(12):3802-3811.
[28]Omar A,Ajar Q,Mukaram H,et al. Community diversity and its seasonal dynamics of soil mites in oasis of the Sangong river watershed of Xinjiang,China[J]. Journal of Xinjiang University(Natural Science Edition),2016,33(1):6-19.
[29]高梅香,劉 冬,张雪萍,等. 三江平原农田地表和地下土壤螨类丰富度与环境因子的空间关联性[J]. 生态学报,2016,36(6):1782-1792.
猜你喜欢
现代畜牧科技(2021年10期)2021-11-19
科学(2020年6期)2020-02-06
中央民族大学学报(自然科学版)(2018年2期)2018-11-09
水利规划与设计(2017年6期)2017-07-18
求知导刊(2016年30期)2016-12-03
科技资讯(2016年19期)2016-11-15
人间(2016年27期)2016-11-11
水利科技与经济(2016年5期)2016-04-22
水利规划与设计(2016年7期)2016-02-28
