
不同虫态菊方翅网蝽在高温逆境下的存活特性
2019-10-22 03:25:30崔旭红姚露晨潘颖佳李若南
生物灾害科学 2019年3期
崔旭红,姚露晨,潘颖佳,李若南,蔡 冲
不同虫态菊方翅网蝽在高温逆境下的存活特性
崔旭红,姚露晨,潘颖佳,李若南,蔡 冲*
(中国计量大学 生命科学学院/浙江省生物计量及检验检疫技术重点实验室,浙江 杭州 310018)
为明确菊方翅网蝽在高温逆境下的生存特性及其对种群发展的影响,研究了菊方翅网蝽卵、5龄若虫、雌雄成虫在35℃,37℃,39℃,41℃,43℃,45 ℃下暴露1~2 h后的存活率。结果表明:39℃及其以下的短时高温暴露对各虫态的存活没有显著影响,41℃及其以上的短时高温暴露会导致各虫态的存活率迅速降低。菊方翅网蝽3个不同虫态之间的存活率存在显著差异。41℃及其以上高温会对菊方翅网蝽的生存和种群发展造成不利的影响。
菊方翅网蝽;高温;存活
菊方翅网蝽((Uhler,1878)),属于半翅目(Hemiptera),网蝽科(Tingidae),方翅网蝽属(,1873),是近年来新入侵中国的外来昆虫。该虫原产于北美地区,主要取食菊科植物,可以严重危害加拿大一枝黄花、向日葵、菊花、菊芋、紫背天葵等多种野生杂草和栽培植物;此外还有危害甘薯的报道[1-5]。菊方翅网蝽的若虫和成虫喜欢聚集在植物叶片背部,可以通过刺吸植物汁液、排泄粪便诱发真菌病害等多种方式对农业生产造成直接或间接的危害。沈佳思等[6]通过对菊方翅网蝽生物学特性的研究推测8—10月是菊方翅网蝽在我国华中以及华东地区发生盛期。这与课题组在杭州地区的田间观察结果基本一致。在韩国和日本,也有报道7—8月是菊方翅网蝽的发生盛期[4,7]。目前菊方翅网蝽在中国台湾、浙江、湖北、江西等地均有发现[3,8-10],这些地区的7—8月正值夏季,日最高气温普遍高于30 ℃,部分地区的最高气温可以达到40 ℃[11]。加之菊方翅网蝽活动性不强,遭遇不利气候条件时难以主动逃避,因此菊方翅网蝽难免会遭受高温胁迫。已有研究表明,入侵昆虫对高温或低温的耐受性是影响其入侵新环境后的扩散以及种群发展的重要因素[12-14]。为了更好的了解菊方翅网蝽在我国的入侵潜能,课题组在室内对菊方翅网蝽的卵、5龄若虫和雌雄成虫进行了短时高温暴露实验,研究了菊方翅网蝽在高温逆境下的存活特性,以揭示该虫对高温的耐受性以及夏季高温对其种群发展的影响。
1 材料与方法
1.1 供试昆虫与寄主植物
供试昆虫:实验室于2016年在中国计量大学网室旱地中建立了菊方翅网蝽种群,寄主植物为加拿大一枝黄花。在进行高温胁迫实验之前,从建立的种群中采集菊方翅网蝽成虫接入养虫笼(60 cm×60 cm×60 cm)中进行继代饲养,随后获得的网蝽用于实验。饲养条件为温度(26±1)℃,相对湿度(75±10)%,光照14 h∶10 h (L∶D)。
寄主植物:剪取加拿大一枝黄花上端20 cm部分,保留干净的叶片,每株保留6~8张叶片,每张叶片长约5~8 cm。将植株茎干用海绵包裹,固定于装有约400 mL水的塑料杯杯盖中央。将其作为清洁苗供实验使用。
1.2 高温暴露处理方法
高温暴露处理在水浴循环仪(型号:9112AA2Y,Poly Science公司)中进行。设置6个靶标温度,分别为35 ℃、37 ℃、39 ℃、41 ℃、43 ℃和(45±0.02)℃。在每个温度梯度下,处理时间为1~2 h。以上每个处理重复5次,每重复卵、若虫、雌雄成虫各30~60头。以直接放置在26 ℃下同等时间的处理为对照,以校正各虫态的存活率。
1.3 不同虫态菊方翅网蝽经高温胁迫后的存活情况
卵:取上述清洁苗放入养虫笼中,并接入生育期成虫,24 h后清除叶片上的成虫,在体视镜下观察标记每张叶片上的卵量。将产卵苗上叶片连叶柄剪下,用湿棉球将叶柄包裹并插入装满水的1.5 mL离心管中,并用易拉膜包裹,防止水分散失。将产有卵的叶片进行随机分组,每组叶片保留60粒卵,多余的卵用解剖针扎死。将叶片放入玻璃试管中,管口塞上少量脱脂棉防止若虫孵化后逃逸。随后进行高温处理,处理结束后放回饲养条件下,10 d后开始调查卵孵化的情况,为了减少操作过程中对卵的损伤,以后每隔1 d调查1次,直到连续5 d都没有卵孵化为止。最后统计卵的孵化率。
若虫和成虫:将5龄若虫和不同性别成虫分别装入玻璃管中(=2.5 cm,=9 cm),在管内放入经过保湿处理的加拿大一枝黄花供取食,并在管口塞上少量脱脂棉防止若虫和成虫逃逸。每组30头若虫或成虫。经高温处理后将网蝽放回饲养条件让其苏醒,2 h后观察其存活情况,并统计其存活率。毛笔轻触若虫和成虫,若附肢不动则认为死亡。
1.4 数据处理与分析
在相同高温条件下不同性别成虫存活率用-Test来检验是否存在显著差异;用单因素方差(One-way ANOVA)分析来检验不同高温暴露条件下菊方翅网蝽的存活率和不同虫态间的耐热性是否存在显著性差异;用多因素方差分析检验温度和时间对粉虱存活率的效应。以上数据分析用统计软件SPSS进行(SPSS24.0,SPSS Inc., USA),显著性检验水平均为<0.05。
2 结果与分析
2.1 高温暴露对菊方翅网蝽卵孵化的影响
39 ℃及其以下较为缓和的短时高温暴露对菊方翅网蝽卵的孵化率没有显著影响(表1)。在35 ℃和37 ℃下菊方翅网蝽卵经过1 h高温胁迫后,其孵化率超过94%,39 ℃胁迫1 h后卵的孵化率约90%,三者之间没有显著差异。经过41~45 ℃高温暴露1 h后,该虫卵的孵化率迅速由85.7%下降至27%,不同暴露温度对菊方翅网蝽卵的孵化率存在显著性差异。在35~45 ℃内对卵暴露处理2 h,卵的孵化率也迅速下降,在43 ℃下暴露处理2 h,其孵化率约为50%,在45 ℃下暴露2 h,没有卵能够孵化。结果表明,菊方翅网蝽在相同温度下暴露不同时间,随着暴露时间的延长,卵的孵化率降低。通过检验结果,43 ℃及以上高温下分别处理1~2 h后其存活率差异显著。
通过双因素方差分析表明,高温和暴露时间对菊方翅网蝽卵的孵化均有显著影响(温度:(5,60)=251.6,<0.05;时间:(1,60)= 29.0,<0.05);温度和时间对菊方翅网蝽卵的孵化有显著的交互作用(5,60)= 5.52;其中温度的效应要高于暴露时间的效应(2温度0.96>2时间0.38)。
2.2 高温暴露对菊方翅网蝽若虫存活的影响
菊方翅网蝽5龄若虫经35~39 ℃暴露1 h后,其存活率没有变化,均为100%(表2)。在41~45 ℃暴露1 h后其存活率由90%迅速下降到10%,不同暴露温度间的存活率存在显著差异(<0.05)。在35~45 ℃暴露2 h后5龄若虫存活情况的变化趋势与处理1 h时相似。5龄若虫经35~39 ℃高温暴露2 h后存活率仍然能够达到96%,当暴露温度达到43 ℃时,5龄若虫的存活率为23.3%,在45 ℃下暴露处理2 h没有若虫能够存活。通过检验结果,43 ℃及以上高温下分别处理1~2 h后其存活率差异显著。
经过双因素方差分析表明,高温和暴露时间对菊方翅网蝽5龄若虫存活都有显著影响(温度:(5,60)=628.3,<0.05;时间:(1,60)= 87.4,<0.05)。温度和暴露时间对菊方翅网蝽若虫存活存在着显著的交互效应((5,180)= 44.9,<0.05);其中温度的效应要高于暴露时间的效应(2温度0.99>2时间0.65)。
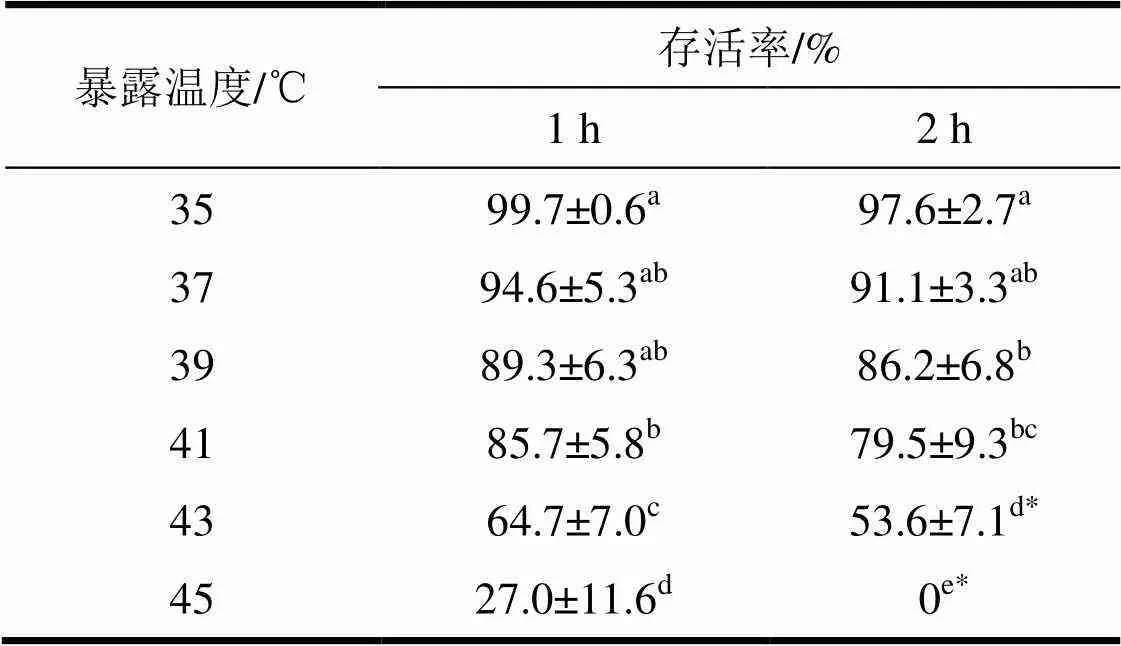
表1 菊方翅网蝽卵经1~2 h高温暴露后的存活率
表中数据为平均值±标准误,同列数据后标有不同字母表示差异显著(<0.05,Duncan检验)。同行数据后标有*号代表差异显著(<0.05,独立样本检验)
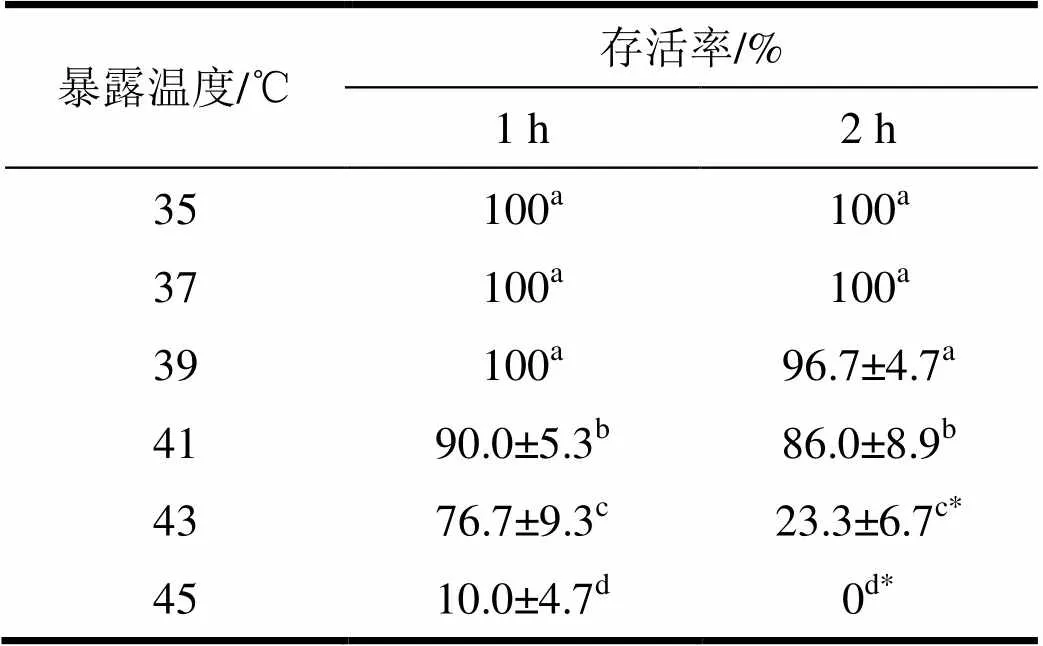
表2 菊方翅网蝽若虫经1~2 h高温暴露后的存活率
表中数据为平均值±标准误,同列数据后标有不同字母表示差异显著(<0.05,Duncan检验)。同行数据后标有*号代表差异显著(<0.05,独立样本检验)
2.3 高温暴露对菊方翅网蝽成虫存活的影响
由表3可知,菊方翅网蝽雌成虫经35~39 ℃暴露1 h后,其存活率没有显著变化,均超过98%。在41~45 ℃暴露1 h后其存活率迅速下降,不同温度间存在显著性差异。菊方翅网蝽的雌成虫在41 ℃下暴露1 h后其存活率为94.67%,在45 ℃暴露1 h后没有成虫能够存活。雌成虫经35~39 ℃暴露2 h后的存活趋势和暴露1 h后的存活趋势相似。菊方翅网蝽雌虫在35~39 ℃下将暴露2 h,其存活率均在100%;在41 ℃下暴露2 h,其存活率为92.0%;在43 ℃下暴露2 h后,雌成虫的存活率显著下降至40%。雄成虫在35~39 ℃下暴露2 h,其存活率略有降低。41 ℃暴露2 h,其存活率下降至88.7%。在43 ℃下暴露2 h,雄虫存活率迅速下降至34.7%,与其它暴露温度存在显著差异;在45 ℃下暴露2 h,没有成虫能够存活。因此,41 ℃及以上高温对菊方翅网蝽成虫的存活率有显著影响(<0.05)。在35~43 ℃范围内,同一暴露温度下,通过检验结果,雌虫比雄虫的存活率高,但差异均不显著(>0.05)。
经过多因素方差分析表明,高温、暴露时间和性别分别对菊方翅网蝽成虫存活有显著影响(温度:(5,120)= 3 646.9,<0.05;时间:(1,120)=192.9,< 0.05;性别:(1,120)= 14.5,<0.05)。温度和暴露时间对菊方翅网蝽成虫存活存在着显著的交互效应((5,120)=154.5,< 0.05);其中温度的效应要高于暴露时间的效应(2温度0.99>2时间0.67)。温度和性别对菊方翅网蝽成虫的存活存在着显著的交互效应((1,120)= 3.0,< 0.05);其中温度的效应要高于性别的效应(2温度0.99>2时间0.1)。暴露时间和性别对菊方翅网蝽成虫的存活没有显著的交互效应((1,120)= 0.27,=0.6)。
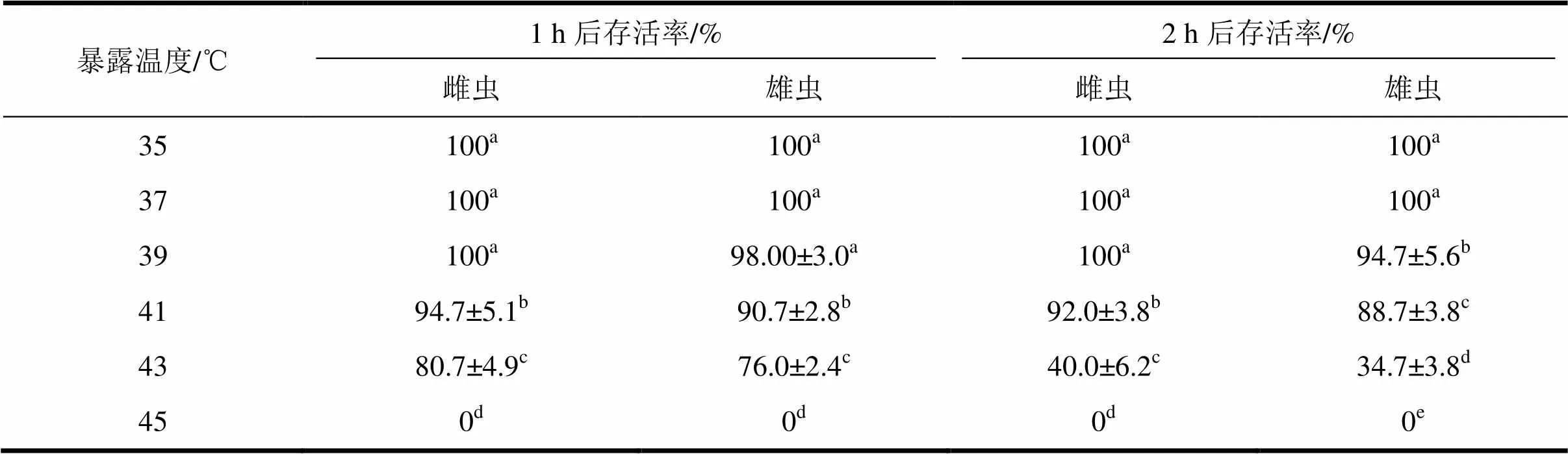
表3 菊方翅网蝽成虫经1~2 h高温暴露后的存活率
中数据为平均值±标准误,同列数据后标有不同字母表示差异显著(<0.05,Duncan检验)。同行数据后标有*号代表差异显著(<0.05,独立样本检验)
2.4 菊方翅网蝽不同虫态对高温暴露耐受性的比较
在39 ℃下暴露1~2 h菊方翅网蝽成虫的存活率均在94%以上,要显著高于卵孵化率。但雌雄成虫和5龄若虫的存活率之间不存在显著差异。当温度上升到41 ℃及其以上高温时,各虫态的菊方翅网蝽存活率下降得很快。例如,在45 ℃下暴露2 h,各虫态的菊方翅网蝽均不能存活,在45 ℃下暴露1 h,各虫态的存活率从大到小依次为:卵、5龄若虫、成虫。
根据多因素方差分析,菊方翅网蝽3个不同虫态之间的存活率存在显著差异((2,240)= 51.93,< 0.05),暴露温度和时间的作用显著(温度:(5,240)= 2 144.28,< 0.05;时间:(1,240)= 375.1,< 0.05)。不同虫态、温度以及时间之间有显著的交互作用(虫态*温度:(15,540)= 32.6,< 0.05;虫态*时间:(3,240)= 38.7,<0.05;温度*时间:(12,240)= 133.3,< 0.05;虫态*温度*时间:(15,240)= 35.2,< 0.05)。
3 结论与讨论
近年来,随着全球气温的上升,夏季最高温度的持续时间不断延长,极端高温天气出现的次数也在不断增加,昆虫遭受高温暴露的情况也变得难以避免。已有研究表明,多食性外来昆虫对高温的耐受性的高低是影响其能否成功入侵的重要因素。例如,B型烟粉虱和温室白粉虱在我国北方混合发生,其种群消长有明显的季节性规律,在夏天高温季节以B型烟粉虱种群为主,而到春秋两季气候趋凉时温室白粉虱种群上升为优势种群[15-16]。两种粉虱对高温的适应性存在差异,是导致它们种群季节动态格局和分布的重要因素。昆虫对高温最为直接的反应就是存活率降低,导致种群数量下降。例如,B型烟粉虱在37~45 ℃的高温条件下暴露1~2 h后,各种供试虫态的生存特性均要高于温室白粉虱,高温对B型烟粉虱存活的影响要显著低于后者;而温室白粉虱对高温敏感,短时间的高温暴露就能导致其存活率显著下降。课题组对菊方翅网蝽的研究表明,41 ℃及其以上高温下暴露1~2 h会导致该虫的3个虫态的存活率显著降低,尤其是43 ℃下随着暴露时间的延长,存活率下降的更为迅速[17]。因此,41 ℃及其以上高温会通过降低菊方翅网蝽的存活率来对其种群产生不利影响。
在自然界中,昆虫常常会遭受较为缓和的高温。这种高温虽然对存活率没有影响,但有可能会造成昆虫的生殖适应性降低,如:产卵量下降,产卵节律的改变等,甚至会影响到后代的存活率和性比[18-19]。这种影响虽然不像存活率降低那么直接,但也会造成后代种群的衰弱。本试验研究结果表明,35~39 ℃对菊方翅网蝽的存活率没有显著影响,该温度范围内遭受高温暴露的昆虫生殖特性是否发生变化还不清楚。因此,为了更好地了解菊方翅网蝽在我国的入侵潜能,高温对菊方翅网蝽生殖特性的影响还需要进一步的研究。
[1] Kato A, Ohbayashi N. Habitat expansion of an exotic lace bug,(Uhler)(Hemiptera: Tingidae), on the Kii Peninsula and Shikoku Island in Western Japan[J]. Entomological Science, 2009, 12(2): 130-134.
[2] Hoshino S. Geographical distribution ofin Hiroshima Prefecture[J]. Plant Protection, 2011, 65(11): 635-639.
[3] 党凯, 高磊, 朱瑾. 菊方翅网蝽在中国首次记述(半翅目, 网蝽科)[J]. 动物分类学报, 2012, 37(4): 894-898.
[4] Kim D E, Kil J. Geographical distribution and host plants of(Uhler) (Hemiptera: Tingidae) in Korea[J]. Korean Society of Applied Entomology, 2014, 53(2): 185-191.
[5] Yoon C S, Kim H G, Choi W Y. First record on the exotic lace bug of asteraceae,(Uhler) (Hemiptera: Tingidae) in Korea[J]. Journal of Environmental Science International, 2013, 22(12): 1611-1614.
[6] 沈佳思, 朱敏, 崔旭红, 等. 外来入侵菊方翅网蝽生物学特性及实验种群生命表[J]. 应用生态学报, 2016, 27(5): 1657-1662.
[7] Mamoru T, Hiroshi T, Manabu S B. Seasonal prevalence in occurrence of(Uhler) on tall goldenrod and the number of catches of the lace bugs by sticky trap[J]. Annual Report of the Kansai Plant Protection Society, 2006, 48: 129-130.
[8] 卢和平, 刘湘, 何益民, 等. 赣州新发现入侵物种菊方翅网蝽[J]. 生物灾害科学,2018, 41(1): 5557.
[9] 董立坤, 王志华, 张涵, 等. 菊方翅网蝽在武汉的分布与危害[J]. 湖北农业科学, 2015, 54(21): 5299-5301.
[10] 虞国跃. 菊方翅网蝽[J]. 植物保护, 2014, 40(5): 7.
[11] 张立生, 王维国, 薛建军, 等. 2013年夏季极端高温天气特征及气象服务分析[J]. 天气预报, 2014, 6(1): 56-60.
[12] 崔旭红, 谢明, 万方浩. 短时高温暴露对B型烟粉虱和温室白粉虱存活以及生殖适应性的影响[J]中国农业科学, 2008, 41(2): 424-430.
[13] Barthell J F, Hranitz J M, Thorp R W, et al. High temperature responses in two exotic leafcutting bee species:and(Hymenoptera: Megachilidae)[J]. The Pan-Pacific Entomologist, 2002, 78: 235-246.
[14] 陈兵. 外来斑潜蝇对热胁迫的适应: 温度、生理机制和生物地理分布[D]. 北京: 中国科学院动物研究所博士论文, 2003: 1-20.
[15] 罗晨, 张芝利. 烟粉虱(Gennadius)研究概述[J]. 北京农业科学, 2000, 18[增刊(烟粉虱专辑)]: 4-13.
[16] Ramos N E, Neto A F, Arsénio S, et al. Situation of the whitefliesandin protected tomato crops in Algarve (Portugal)[J]. Bulletin Oepp/eppo Bulletin, 2002, 32: 11-15.
[17] 崔旭红. B型烟粉虱和温室粉虱热胁迫适应性及其分子生态机制[D].北京:中国农业科学院研究生院博士论文, 2007: 17-25.
[18] Bodlah M A, Gu L L , Tan Y, et al. Behavioural adaptation of the rice leaf folderto short-term heat stress[J]. Journal of Insect Physiology, 2017, 100: 28-34.
[19] Zhou Z S, Guo J Y, Min Luo, et al. Effect of short-term high temperature stress on the development and fecundity of(Coleoptera: Chrysomelidae)[J].Biocontrol Science and Technology, 2011, 21(7):809-819.
Survival Characteristics of Different Developmental Stage ofUhler (Hemiptera: Tingidae) after Exposure to High Temperature Conditions
CUI Xu-hong, YAO Lu-chen, PAN Ying-jia, LI Ruo-nan, CAI Chong*
(College of Life Sciences/Zhejiang Provincial Key Laboratory of Biometrology and Inspection & Quarantine, China Jiliang University, Hangzhou 310018, China)
The survival rates ofdifferent developmental stage ofwere studied after exposure to high temperatures to determine the effects of high temperature conditions on its population development. The eggs, 5th- larvae and both male and female adults ofwere exposed to each of the six temperature regimes, 35, 37, 39, 41, 43 and 45 ℃ in the water bath for 1~2 hours. The results showed that the survival rates of different stage ofwere not significantly affected by the temperatures varied from 35 to 39 ℃. However, the survival rates of the three tested stagesdecreased quickly when the temperatures exceeded 41 ℃. There were significant differences in three tested stages of. The results meant thathad a certain tolerance to the moderate high temperatures, but the high temperatures above 41 ℃ could adversely affect the survival and population development of.
; high temperature; survival
http://xuebao.jxau.edu.cn
10.3969/j.issn.2095-3704.2019.03.47
S433.3
A
2095-3704(2019)03-0218-05
2019-08-20
国家自然科学基金项目(31601695)和浙江省自然科学基金项目(LY17C150003)
崔旭红(1977—),副教授,主要从事外来入侵昆虫种群形成与扩张等方面的研究,xuhongcui@cjlu.edu.cn;
蔡冲,副教授,ccjacn@cjlu.edu.cn。
崔旭红, 姚露晨, 潘颖佳, 等. 不同虫态菊方翅网蝽在高温逆境下的存活特性[J]. 生物灾害科学, 2019, 42(3): 218-222.
猜你喜欢
农业工程学报(2022年10期)2022-08-22 07:21:20
天津农业科学(2022年5期)2022-05-31 21:01:11
江苏农业科学(2022年2期)2022-02-15 14:47:16
湖南农业大学学报(自然科学版)(2021年2期)2021-05-06 00:22:12
爱你(2018年24期)2018-08-16 01:20:42
爱你·阳光少年(2018年8期)2018-05-14 09:01:18
江苏农业科学(2017年12期)2017-09-16 00:48:09
中国照明(2016年4期)2016-05-17 06:16:18
长江大学学报(自科版)(2015年21期)2015-12-04 01:34:18
植物保护(2015年4期)2015-11-28 07:28:28
