
降水和遮荫对侧柏林地种子萌发的影响
2019-10-15 01:42翟国锋3刘幸红吕雷昌吴德军邢世岩2葛忠强
种子 2019年9期
王 斐,翟国锋3,刘幸红,吕雷昌,吴德军,邢世岩2,葛忠强
(1.山东省林业科学研究院,济南 250014;2.山东农业大学,山东 泰安 271018;3.山东淄博沂源毫山林场,山东 沂源 256112)
我国华北和西北地区栽培的大面积侧柏(Platycladusorientalis)人工林对改善生态环境起到非常重要的作用,然而其更新也成为我们面临的重要课题之一。目前,有关侧柏林天然更新的研究并非少见[1-2]。一些报道通过侧柏林地内幼苗幼树的调查研究了侧柏林天然更新的潜在可能性[3-4],也有报道从侧柏林的区域划分中研究了侧柏幼苗幼树空间分布的异质性[5]以及林分自身的空间结构对侧柏林地更新的影响[6]。甚至有从侧柏林窗内日照轨迹的计算机模拟与侧柏苗的空间分布特征研究光照和湿度的变化对侧柏幼苗幼树生存的影响[7]。侧柏种子萌发受制于土壤水分的有效性和适度的光照条件[8]。这是否适用于大区域范围内的侧柏林分尚不清楚,而且一些夏季降水尤为充足的年份林地内侧柏种子萌发和籽苗出土也不多见。为此,在山东省内对侧柏当年生籽苗进行更大范围的调查和研究。应用地统计方法进行了降水的时空解析以及侧柏种子萌发和籽苗空间分布的研究。结合多树种种子萌发的室内试验研究,论述了侧柏林种子萌发和保存对于优化的时空环境的依赖性,从而进一步解析了侧柏种子集中萌发和幼苗幼树聚集分布与其环境适应性的关系,为人工促进侧柏林更新奠定理论基础。
1 材料和方法
1.1 山东省侧柏林地种子萌发和降水时空分布的统计分析
山东省107个气象站点2017—2018年逐日降雨量的数据来自于中国国家气象中心气象数据网的对应站点。雨季降水量为7—9月份降水量之和,前9个月降水量是1—9月份各月降水量之和。在统计这2项指标的基础上,用ArcGIS的径向基函数插值法(RBF)对其进行地统计分析。统计结果从小到大依次将降水量分成10个等级,然后用10种不同深度的蓝和红色绘制降水量地统计分布图。与此同时,按数量多少将单位面积上萌发的籽苗数量分为5级或10级。并用大小不等的圆标记在其相应的地理位置上。
侧柏种子萌发出苗数的调查,分别于2017年和2018年晚秋季节用Top定位法进行统计。调查地的选择以具有代表性且交通方便的石灰岩山地侧柏林为准,包括山东南部的枣庄、北部的淄博、济南和潍坊局部石灰岩山地共计8个市、县的18个地点。调查分别在阳坡和阴坡的汇水坡面水土再分配较为集中的地块进行。选择籽苗最为密集的地块设定1 m2样方,分别统计其中春夏萌生并保存下来的具有数对或十几对初生叶以及秋季萌发仅有一对子叶或一对初生叶的侧柏籽苗数,以便研究春、夏、秋降水量与侧柏种子萌发的关系。每地点各个坡向的样点分别重复10~15次。
1.2 室温环境中侧柏、荆条和酸枣种子的发芽试验
于2017年9月初采集侧柏成熟种子、10月初采集荆条成熟种子,10月中旬采集酸枣成熟果实经调制后获得的成熟种子在室内隔窗阳光下晒干。侧柏种子过8目筛后,置于室温环境中待用。2018年1月8日用初始温度为40 ℃的温水浸种、48 h后置入塑料盘内,盖湿纱布在自然室温环境(13~18 ℃)下进行冬季发芽试验。保持种子湿润,并随时补充水分。从种子开始发芽按一定的时间间隔统计发芽数,并计算发芽率。发芽率的计算公式如下:
Pi=100×Gi/Ti
式中i=1,2,3,分别表示1-侧柏,2-荆条和3-酸枣。Pi、Gi和Ti分别是第i树种的发芽率(%)、发芽种子数和试验种子总数。鉴于侧柏、荆条和酸枣单粒种子的重量差异较大,试验时使用的种子总数各不相同,如T1(侧柏)=198;T2(荆条)=513;T3(酸枣)=90。
春末夏初的发芽试验始于4月15日,同样用初始温度为40 ℃的温水浸种、48 h后置入塑料盘内,盖湿纱布在自然室温环境(20~25 ℃)下进行发芽试验,按公式1计算发芽率。盛夏的发芽试验始于7月23日,试验方法同前,而试验时的室温为30~31 ℃。
1.3 侧柏种子后熟特性的试验研究
分别于8月末采集变黄即将裂开的球果,9月初和9月中旬采集刚裂嘴或已经裂开的球果。在室内自然温度下晾干,获得的侧柏种子过8目筛待用。于9月下旬开始发芽试验,分别取8月30日、9月7日和9月15日采集的种子各30粒。用初始温度为40 ℃的温水浸种、48 h后置入沙包内,盖湿纱布在自然室温环境(26~30 ℃)下进行发芽试验,重复3次。按公式1计算发芽率,其中T=30。
1.4 侧柏母树生长指标观测
对于衡量侧柏母树生长状态的胸径指标的观测所采用的方法为胸围测量法。使用常用的钢卷尺在胸高部位测得。样树选自籽苗统计样方周围最近的4~5株下种母树,每个调查地点20~22株。
1.5 数据统计分析
方差分析和相关分析应用Office 2003软件完成。地统计分析用ArcGIS 12.02软件进行。
2 结果和分析
2.1 降水量的空间分布异质性与侧柏种子萌发和籽苗保存的对应关系
尽管有关林地侧柏种子萌发的研究已有许多报道,但是结合气象数据的解析进行大尺度的调查研究并不多见。为此在2017年和2018年在山东省进行了深入的调研。结果发现一定时期内的降雨量是侧柏林地土壤含水量多寡的决定因素,进而也是侧柏种子萌发的重要限制因素。图1 a是应用径向基函数插值法基于山东省境内107个气象站点2018年的前9个月之降水量数据制作的地统计分析图(底色)和一些典型林地侧柏出土籽苗的数量图(圆形标识)。结果表明该年度在遭受数次台风的袭扰后,山东省降水量偏多而且明显地分配不均。泰沂山北麓的淄河和弥河流域为中心的地域降水相对丰沛,个别地方甚至超过1 000 mm。这在该地有气象历史以来较为少见。持续的阴雨天气不仅给侧柏种子萌发提供了充足的吸胀水源。而且也降低了林地和土壤的温度。从而促进了种子夏季的萌发和保存。结果使得春夏萌发出土且保存下来的、具有数对初生叶的籽苗较多。最多者每平方米接近40个,而在平阴(图1 a-14,15)和长清(图1 a-13)周边因降水量较少而很少有籽苗出土或存活。在许多调查地点几乎没有发现此类籽苗的踪影。
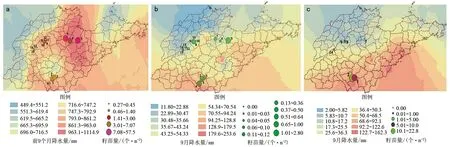
注:a.数次台风袭扰后2018年前9个月的降水量和山地阴坡具有数对初生叶的籽苗量(个·m-2),样点数n=15;b.2018年9月降水量和山地阴坡秋季萌发的仅有一对子叶或初生叶的籽苗量(个·m-2),n=15;c.2017年9月降水量(mm)和侧柏籽苗数(个·m-2)的分布,n=6。图1 山东省降水量径向基函数(RBF)插值法地统计分析和单位面积籽苗数量分布图
2018年秋季的9月以后,降雨量逐渐减少,尤其是北部地区。尽管该月有几次由南向北的降雨过程发生,但并没有影响到北部的泰沂山北麓地带。山东省9月降水量基本上维持南多北少的格局(图1 b)。经过对上述典型地区侧柏林分内秋季新萌生的、仅有一对子叶或初生叶的籽苗统计结果表明,在鲁南枣庄市的薛城(图1 b-4)和山亭区周边以及降水量稍多的昌乐(图1 b-5)等地侧柏新生苗稍多,而9月降水偏少的济南西部(图1 b-12,13,14,15)地区最少,甚至没有。尽管淄博、青州(图1 b-6,7)等地雨季降水量巨大,但是因为在侧柏种子集中成熟的9月降水量剧减而新生的侧柏籽苗也不多。
2.2 母树生长状态与侧柏种子萌发和籽苗生长
综上所述,侧柏种子的萌发和籽苗数量随降水的波动变化而有多有少。且降水多少对种子吸胀和萌发出土影响显著。然而,在一些降水量整体偏少的地区以及降水从多到少的过渡区,时而保存下来的籽苗并不算很少。其实,所调查的籽苗数量不仅受当年降水的影响,而且与上年秋季萌发的籽苗数等有关。在枣庄周边(图1 c-1,2)2017年秋季9月降水量较大且集中,这为侧柏种子萌发和籽苗保存创造了较好的条件,该年秋季萌生侧柏籽苗为数较多(10~20个·mm-2)。2018年在该地调查的籽苗数(图1 a)在一定程度上受2017年秋季侧柏籽苗较多的影响(图1 c-1,2)。不仅如此,2017年在侧柏较为集中栽培的区域研究表明,该年度侧柏籽苗出土的数量与降水量之间也存在较为显著的相关性(图1 c)。该年度9月份济南降水量大幅下降,甚至只有5.6 mm。所以2017年在济南周边(图1 c-7,8,9,14)的侧柏林地内很难找到新萌发的当年生侧柏籽苗。
在图1 a中值得注意的是,尽管个别样点地处降水丰沛的多雨区,其籽苗数并非很多(图1 a-6)。经调研发现,这种籽苗数量的分化在一定程度上受母树生长状态和立地环境的影响。图2 a是2018年雨季降水非常丰沛而集中的泰沂山北麓部分地区保存下来的侧柏籽苗与周边母树胸围呈正相关关系,即母树越大、生长越旺盛,树冠周边的籽苗越多。而且这些侧柏籽苗大多集中在汇水坡面、水沟两侧水土再分配受益最明显的地块。
尽管如此,经调查发现山地阴坡萌发的侧柏籽苗数量明显多于阳坡(图2 b,p<0.01),并且在调查样地的阳坡与阴坡之间并没有观测到母树胸围之间的显著性差异(图2 b,p=0.141)。也就是说,尽管阴坡往往比阳坡的侧柏生长量大,但是所观测到的侧柏籽苗大量萌发的林地之下二者之间的胸围没有显著差异。然而,2种立地环境下侧柏籽苗数量差异巨大,且达到极显著水平。这表明,阴坡更加适宜的阴湿环境是该年度侧柏种子萌发和生长的良好条件,侧柏种子萌发并非需要夏季的高温环境。然而,盛夏的高温和全光环境不仅提高了土壤和大气的温度,更加重要的增加了蒸气压差,加快了水分的蒸发蒸腾。这时即使降水充足、土壤含水量适宜,林窗等全光照环境中侧柏籽苗难以出土。侧柏种子属出土萌发类型,播下去的种子即使已经萌芽,在土壤表层高温干燥环境中下胚轴滞育而不能出土(图2 c)。久而久之,在没能建立起稳定的自营体系之前而夭折。然而应用石块侧方遮荫处理的种子不仅可以萌发,且籽苗宜生长正常(图2 d)。石块隔绝了阳光直射、降低了局部土壤温度、增加了土壤湿度,此类小环境的创建促进了种子萌发和幼苗的生长。

注:a为单位面积上籽苗数(个·m-2)与母树胸围之间的相关关系;b为阳坡和阴坡籽苗数(个·m-2)、母树胸围的差异;c为从山顶阳坡全光照下扒出的萌发侧柏籽苗,因胚轴滞育而不能出土;d.山顶阳坡全光照下用石块侧方遮荫的籽苗。图2 鲁北2018年雨季强降雨区侧柏籽苗春夏季萌发、保存与母树胸围的关系
2.3 侧柏种子萌发最优时期以及与伴生树种间的竞争关系
侧柏林更新受制于复杂的生态因素,除了非生物环境的影响外,一些植物种的竞争也是不可小视的因素之一。为此分别在冬季、春季和夏季进行了侧柏、酸枣和荆条种子萌发试验。始于2018年1月8日的发芽对比实验结果表明,在冬季室温(13~18 ℃)环境中,侧柏种子经催芽之后,萌发速度与秋季试验接近。第13天或第14天开始发芽,20 d达到或接近高峰值,以后持续较长时间陆续有少量种子萌发。这表明侧柏种子萌发并不要求较高的气温,而荆条在这种相对低温的环境中不能萌发。试验直到春季4月初气温升高到20 ℃以上,才逐渐有少量荆条种子发芽荆条种子在气温30 ℃时萌发最快[9],且萌芽率不高。酸枣种皮厚而木质化,吸胀和打破休眠过程较长,同样在发芽试验开始3个月后才开始有零星萌发(图3 a)。
始于4月中旬的重复试验中,室内气温已经上升到24~26 ℃。侧柏种子仍然需要十几天(4月30日)开始萌发而后迅速达到峰值,且发芽率较高(图3 b)。在这一相对较高的气温环境中,尽管峰值出现的稍晚荆条发芽的起始时间与侧柏不相上下且持续萌发。酸枣照样需要很长的吸胀破壳时间直到试验结束没有种子发芽。事实上,这与林地内秋季常看到侧柏籽苗出土、而春夏季节常看到荆条集中萌发,酸枣很少有大量集中萌发的现象相吻合。以往大量的试验研究表明,林地内侧柏种子在夏季萌发较为少见,7月下旬(气温30 ℃左右)进行的室内种子发芽试验表明,在5 d之内,荆条已经发芽10%以上,而侧柏和酸枣的发芽率为0(图3 c)。8 d以后荆条累计发芽率达到20%以上,且单日发芽已经很少;侧柏发芽率还不足5%,酸枣依然没有发芽。2周之后侧柏发芽率刚到10%左右。所以,夏季高温不利于侧柏种子的萌发。
这3次发芽试验结果有1个共同的趋势,侧柏种子基本上不存在休眠的问题,且要求不太高的温度。相对于温和的条件,高温环境中种子发芽较为缓慢;在林地环境中则大量集中萌发于4月和9月降雨量充足的年份。荆条在低温环境中难以萌发,只有到春夏之交气温转暖之时才开始萌发,高温环境中萌发较快。而酸枣受制于厚而硬的种皮需要2—3个月的破壳过程。这在自然环境中有利于长期持续地保持种子活力,有利于种子自身对环境的选择。相比之下,荆条只有在春季或夏季萌发的概率较大,而侧柏更加适合母树下种后气温凉爽的秋季或春季萌发出土。
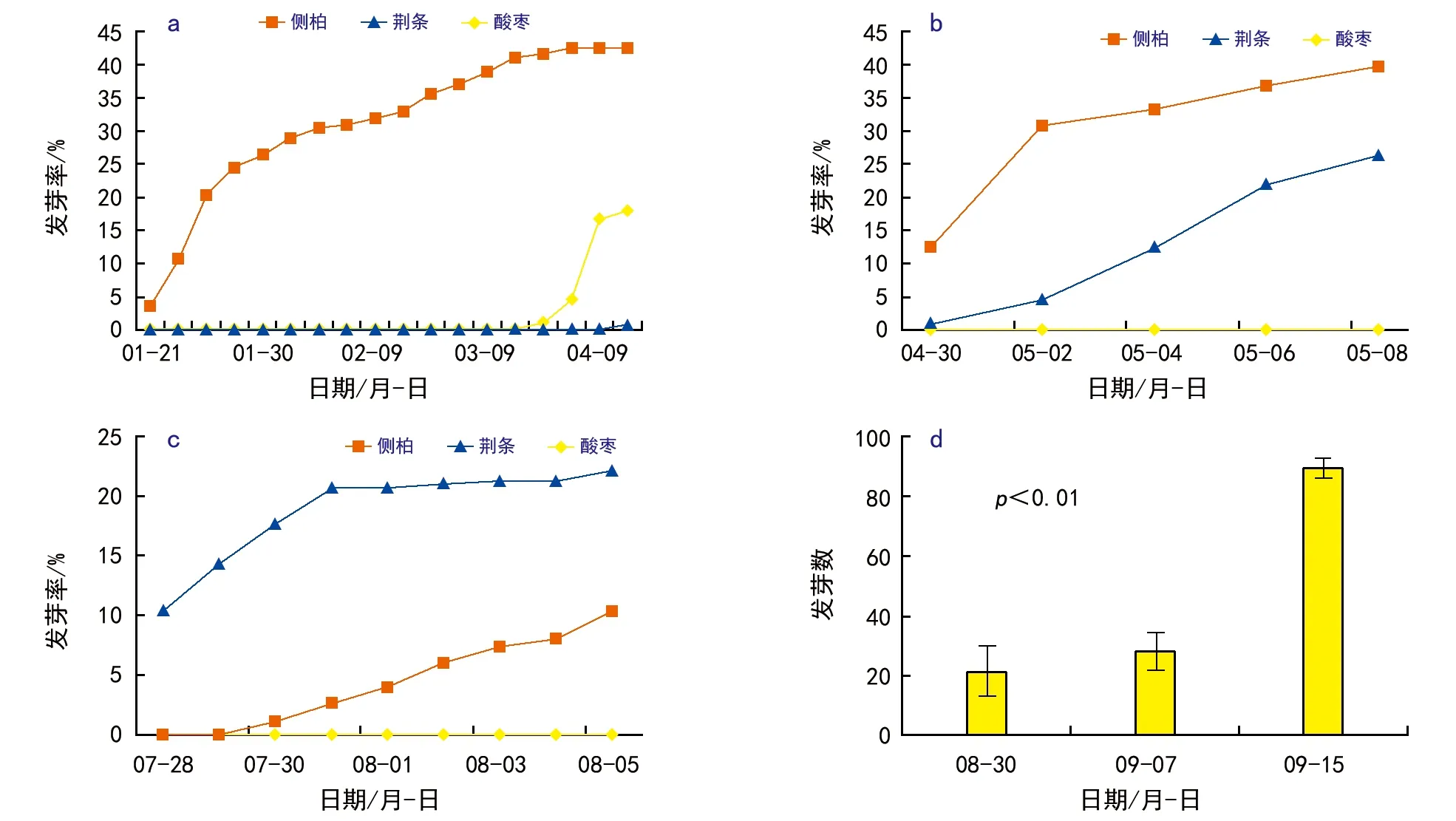
注:a.始于2018年1月8日的室内发芽试验;b.始于2018年4月15日的室内发芽试验;c.始于2018年7月19日的室内发芽试验的发芽率;d.侧柏种子不同采收时间的发芽率。图3 侧柏及其常见竞争种荆条和酸枣当年生种子萌发试验
试验表明在秋季播种使用未完全成熟的侧柏种子时,种子萌发需要渡过一个短暂的后熟期。2017年8月末,球果已经变黄或变褐,尚未完全成熟,实验室发芽率为20%左右。9月种子大量成熟落地后萌发即不存在问题,与球果尚未开裂的种子相比差异显著(图3 d,p<0.01)。
3 讨 论
植物种子从萌发到新的生命周期开始需要经过一个吸胀、增加含水率,再到胚根、胚芽发育,重建SPAC、形成独立的自营新生命体的过程。在此过程中水是至关重要而且必不可少的核心物质。水分和热量的平衡是植物生长和发育以及适应环境的重要平衡机制[10]。在水分和热量失衡状态下易于遭受环境胁迫而受害。植物种子萌发从异营到自营的过渡期和幼苗期本身对干旱等环境的胁迫更加敏感[11-13]。就侧柏播种和种子萌发出土而言,以往的研究表明侧柏种子在阳坡或山顶空旷的全光照林窗环境中幼苗幼树数量少[7-8],而在汇水坡面和集水池旁侧柏种子萌发容易、籽苗数量很多。
相比之下,在林冠遮蔽下的疏林地或树荫处,地温低、湿度大,播种和浇水试验出苗率较高。而到了晚秋,日照减弱,气温降低,无论全光照环境还是疏林下,侧柏种子在充足的水分保障下,籽苗大量出土[5,7-8]。这个季节往往是我国北方干旱的枯水期。水分又成为侧柏种子萌发的限制因素。在偶尔发生的晚秋多雨年份促生大量的籽苗,由于气候急转直下而更加干冷,籽苗很难保存下来。归根结底,维持种子和籽苗的水分和能量平衡是其萌发出土的关键因素。
事实上,在我国西北的黄土丘陵地区以及北方干旱的沙漠地区侧柏大多分布和生长在沟壑的绝壁上,究其原因在于这些立地环境中不仅有水分再分配的来源,而且还有光照热量的空间遮蔽。然而,极端干旱的气候又限制了种子的吸胀和萌发,因此侧柏仅生存在局部的小环境之中。相比之下,在山东半干旱或半湿润的气候相对有利于侧柏种子的萌发和成苗,林地天然更新苗木数量增多,侧柏林的覆被率也相应增加。即使在夏季种子同样不易萌发,但是在水分和能量再分配受益的沟谷、汇水坡面等地,水土条件优越,林木生长茂密,树体活力旺盛,种子优良。在高大母树树体的遮蔽之下,于降水充沛的年份春夏季节萌发出土一些侧柏籽苗也不足为奇。在这些立地环境中侧柏母树同样因环境优越而生长健壮。也就是说在年复一年的水土、营养再分配的偏爱下,侧柏母树生长的立地环境湿润又肥沃。优越的环境导致侧柏母树枝繁叶茂、种子质量相应地更加优良。浓密的树冠创造的适度遮荫环境又为种子萌发和生长创造良好的条件,所以成为侧柏幼苗幼树聚集生长的根源。这些立地环境也是一些耐阴的落叶阔叶树种适生的环境,甚至对于广泛分布的竞争种荆条等阔叶树种的生长也更加有利。在立地条件较为优越、水肥资源丰厚的环境中一些耐阴的高大落叶阔叶乔木成为侧柏更新的主要障碍。如常见的杂木树种榆树、苦楝、臭椿、君迁子、元宝槭、桑、槐、构树等,甚至在一些大型的沟谷内业已形成块状的阔叶落叶林。然而,这样的立地大多适宜农耕、也是枣、杏、山楂和苹果等果树栽培的立地,栽培侧柏者为数不多。侧柏大多栽培在以灌木为主的瘠薄山地环境中。在这些立地上最为常见的竞争植物种是荆条和酸枣,在山东省境内的石灰岩山地上尤为常见。林地侧柏种子萌发不仅在高温干旱环境中萌发较少,而且业已萌发和成活的幼苗也容易夭折。
侧柏、荆条和酸枣在不同季节种子萌发试验研究表明,在温和凉爽环境中可以在避免其他物种竞争的前提下促进侧柏种子萌发。9月母树下种高峰期也是北方旱季和低温逐渐来临之时,降水不足种子不能吸胀,往往错过萌发的最佳时期。春季补水不仅可以促生侧柏苗,而且可以促进鬼针草和荆条等竞争植物的种子萌生。事实上,侧柏籽苗与荆条等同时大量萌发,因生长缓慢而受压的情形并不少见。而且,春末夏初的高温干旱对籽苗成活和保存的限制性更加突出。因此,在秋季适时补水促进侧柏集中迅速萌发是人工促进侧柏更新的有效途径。依据侧柏种子萌发不要求较高的气温之习性,在秋季的9月人工促进种子萌发是促进侧柏林天然更新的有效措施。而夏季侧柏籽苗栽培中适度的遮荫措施也有利于其生长。也可以就地取材应用石块对刚萌生的籽苗实施侧方遮荫保护。
Daubenmire[14]对有关温度与植物物候之间关系的论述表示,形成于夏季的多年生植物芽制约着来年春天的发育,强降雨、高云量以及相对低温常导致发育的迟滞。据观察,在夏季降水量过多的年份侧柏种子成熟的推迟往往也是影响种子萌发的因素之一。
猜你喜欢
农业与技术(2022年22期)2022-12-02
新农民(2022年22期)2022-11-07
农村实用技术(2021年11期)2021-12-03
山东林业科技(2021年4期)2021-08-31
星星·诗歌原创(2021年3期)2021-06-01
福建林业科技(2020年3期)2020-10-25
现代园艺(2020年17期)2020-08-31
诗选刊(2019年9期)2019-11-20
扬子江(2018年1期)2018-01-26
学生天地·小学低年级版(2016年9期)2016-05-14
