
高陡岩质边坡覆绿植物成活的生态因子分析
——以安庆市大龙山集贤关为例
2019-10-10 07:00:48白冰珂赵国红宁立波周华健余启明
安全与环境工程 2019年5期
白冰珂,赵国红,尹 峰,宁立波,张 杨,周华健,余启明
(1.中国地质大学(武汉)环境学院,湖北 武汉430074;2.安徽省国土厅公益性地质调查管理中心,安徽 合肥 230092)
大量石料的开采,使得采石场的生态系统遭到破坏,系统的自组织功能基本丧失,自然的生产功能及自我调节的能力严重退化[1-2]。在采石场关闭后,遗留下来的裸露岩质边坡因缺乏土壤、水分等适宜植物生长的立地条件,使得边坡在无人工干预的条件下自然修复极其困难,且大面积裸露的岩质边坡与其周围的生态环境格格不入,严重影响了当地的地貌景观,并进一步造成生态系统的持续恶化[2-3],随着“生态文明建设”思想的提出,人们对环境问题的关注度不断提高,针对一些废弃石料采石场的高陡岩质边坡覆绿工作也逐渐开展起来。
目前针对岩质边坡的覆绿方法很多[3-8],常见的有格构法、鱼鳞坑法、喷播法和生态袋法。其中,格构法常用于公路及铁路边坡防护工程中;鱼鳞坑法只适用于小坡度边坡覆绿[8];而采用喷播法修复的边坡在形成稳定植被前,岩壁受雨水冲刷易导致基质脱落,致使覆绿效果不佳[9];生态袋法在边坡覆绿工程中常因出现再造坡体结构不稳定而使绿化效果不佳[10],且仅适用于坡度≤63°的边坡[11]。上述岩质边坡覆绿方法均是将植物与岩体割裂开来,侧重于对边坡的人工改造而忽视了对岩体本身这个储能场所的应用,从而影响了覆绿效果。这主要是因为没有从生态学的角度对高陡岩质边坡覆绿工程中影响植物成活的生态因子进行分析,进而重新构建植物与岩体这个生态系统。一般来说,在土体中影响植物成活的生态因子有光照、温度、水分等[12],但高陡岩质边坡覆绿工程中影响植物成活的生态因子有哪些?通过查阅文献得知这方面的研究尚属新的课题。为此,本文以安庆市大龙山集贤关高陡岩质边坡覆绿工程为例,通过对高陡岩质边坡的特点进行分析,得出影响高陡岩质边坡覆绿植物成活的生态因子,并通过野外试验对覆绿边坡岩体内水分、温度状况进行监测,同时对覆绿边坡岩体裂隙发育情况进行调查,最后对所获数据进行整理与分析,得到影响研究区内高陡岩质边坡覆绿植物成活的生态因子的阈值范围,以为高陡岩质边坡覆绿工程提供理论支撑。
1 研究区及覆绿工程概况
1.1 研究区概况
研究区集贤关位于安徽省安庆市区北部,经多年的露天开采,山体土壤被剥蚀,形成了许多高陡的裸露岩质边坡,坡度≥60°,相对坡脚高度约80 m。该地区属于扬子地层区下扬子地层分区安庆地层小区,出露的地层主要为三叠系下统南陵湖组(T1n),岩性可分为两部分,下部为灰色中厚层微晶灰岩,夹灰、灰黑色粉砂质黏土页岩,具缝合线构造及重结晶现象;上部为灰白色中厚层微晶灰岩,含黄色薄层泥质条带。研究区属季风性湿润气候区,气候温和,雨量充沛,无霜期长,光照充足,年平均气温在17~18°,12月份平均气温为3.5℃,7月份平均气温为29℃,年平均降雨量为1 350.1 mm,多年平均蒸发量为1 315.4 mm。
1.2 覆绿方法简介
对集贤关高陡岩质边坡所采取的覆绿方法是“地境再造法”[13],其覆绿工程示意图见图1。该方法依据生态地质学理论从植物生长所需的地境入手,以岩体为基础,运用工程技术手段对植物立地条件进行改造,使之能够形成一个初期适合植物成活的地下生境;植物成活后,通过根系与岩体及外界组成的开放系统进行物质与能量的交换,将其与岩体统一起来,进而维持植物自身的生长。
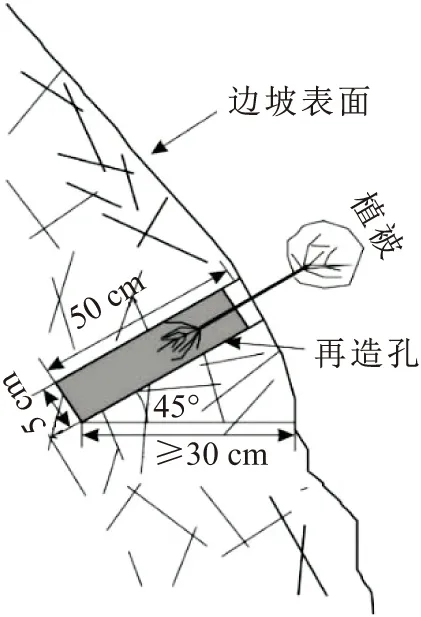
图1 地境再造法覆绿工程示意图
1.3 覆绿植物成活率调查
考虑到是在岩质边坡条件下种植植物,植物是在天然条件下汲取物质与能量来供给自身生长,因此本次研究均选用耐旱类植物:黄栌、连翘、樟子松、爬山虎、金银花、迎春、刺槐、构树,共计150株。植物种植2年后,于2018年6月课题组对研究区高陡岩质边坡覆绿植物的成活率进行了野外调查,其调查结果见表1。
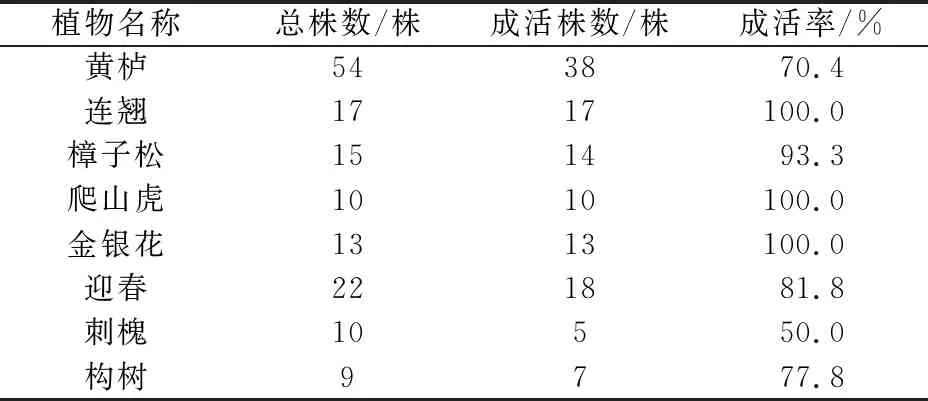
表1 研究区高陡岩质边坡覆绿植物成活率的调查结果
由表1可见,除刺槐和黄栌成活率较低外,其他覆绿植物成活率均较高,覆绿植物的总体成活率达到了81.3%,表明该覆绿方法在研究区切实可行,也说明研究区覆绿边坡岩体具备种植植物的条件。
2 高陡岩质边坡条件下影响覆绿值物成活的生态因子分析
植物的生长对环境有一定的要求,环境对植物的影响主要通过一些生态因子来体现。在高陡岩质边坡条件下种植植物与一般条件下种植植物大不相同,高陡岩质边坡具有如下鲜明的特点:边坡多为大于60°的裸露岩壁[14-15],岩体的导热率大于土壤[16],岩体的储水能力与土壤相比相差甚远[17]。由于受岩体胁迫作用的影响,一般在土体中不是限制植物成活的生态因子在岩质边坡条件下可能就显得尤为重要。根据研究区情况,将影响高陡岩质边坡覆绿植物成活的主要生态因子确定为:岩体裂隙因子、岩体内的温度因子、岩体内的绝对湿度因子。
2.1 岩体体裂隙因子分析
在高陡岩质边坡条件下,裂隙是否发育是限制植物能否成活的重要条件之一[18-19]。岩体体裂隙对于植物的成活有重大意义,它既是岩体内水分储存与运移的场所,也是植物根系在岩体中生长的唯一空间[20];岩体体裂隙发育程度决定植物根部的透气性,岩体内部良好的透气性使植物根部能更好地进行呼吸作用,更容易地吸收水分与营养物质;同时在岩质边坡这种苛刻的生境中,由于水分的缺乏,在岩体的胁迫作用下植物根系不得不增大其生长范围,进而需要从更大的裂隙空间中来获取满足自身生长发育所需的水分。本文主要从以下几个方面对岩体裂隙因子进行分析:
(1) 岩体裂隙率。在《地球科学大辞典》中,岩体裂隙率定义为:岩石中裂隙的体积与包括裂隙在内的岩石体积之比(即体积裂隙率)。本研究对研究区覆绿边坡试验场地内3个测量点内10组裂隙中的52条裂隙进行了调查与统计,并计算岩体裂隙率。结果发现:每个测量点内一般有3~4组裂隙,每组有2~10条裂隙,岩体体裂隙率为1.77%~4.58%,岩体体裂隙率平均值为2.95%;岩体裂隙隙宽为0.1~1.75 mm不等,其中隙宽大于0.5 mm的岩体裂隙约占总裂隙的41.3%,隙宽大于1 mm的岩体裂隙约占总岩体裂隙的18.3%。植物通过根系吸收养分和水分等,而植物根系所能伸入的最小岩体裂隙约为0.1 mm,且此时受窄裂隙的胁迫作用,植物根系为了更大效率地获取养分和水分等,根的形态发生了变化,呈扁平状生长,可极大地增加植物根系与岩体裂隙内有机质的接触面积[19-20]。
(2) 岩体裂隙发育优势方向。岩体裂隙发育优势方向指向岩体内部时,岩体能有效地截取被雨水从山坡顶部冲刷下来的含有养分的土壤;岩体裂隙发育优势方向指向岩体外部时,降雨时岩体内的水分能及时导出。对覆绿边坡3个测量点内的岩体裂隙倾向、倾角作赤平投影图,并分析岩体裂隙发育的主方向,见图2。
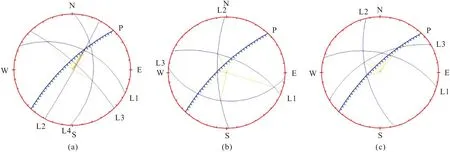
图2 覆绿边坡岩体裂隙赤平投影图
由图2可见,覆绿边坡的坡面方向与岩体裂隙的倾向多为相反,岩体裂隙的优势方向多与岩壁呈一定角度相交,其中裂隙以倾向北东15°~25°方向最为发育,倾角为46°~60°,岩体裂隙倾向南西256°~268°方向次之,倾角为64°~72°,岩体裂隙倾向南东和北西方向也有发育;岩体裂隙发育方向多与边坡呈一定角度相交并指向岩体内部,同时发育的岩体裂隙能有效地截取被雨水从山坡顶部所冲刷下来的含有养分的土壤。
(3) 岩体裂隙的连通性。对于岩质边坡,其岩体内部的透气性与导水性主要由岩体裂隙的连通性来控制。对覆绿边坡岩体裂隙的连通性进行调查的方法如下:在覆绿边坡共设计226个种植孔,种植孔施工完毕后将其中的220个种植孔内全部注满水(由于种植孔口向上倾斜45°,实际注水只有种植孔体积的一半),24 h后观察种植孔内水量的变化。结果发现:其中有103个种植孔内水量下降值超过了80%,有78个种植孔内水量下降了约45%,有39个种植孔内水量下降了约20%。根据种植孔内水量的变化情况,可知该覆绿边坡岩体裂隙的连通性较好,可为覆绿植物的成活提供有利条件。
岩体裂隙既是植物根系的生长空间,也是沟通植物与岩体及外界相互作用的媒介;岩体裂隙发育的优势方向与岩体裂隙的连通性共同指示出岩体的导水性及贮水性,当岩体裂隙发育优势方向倾向于岩体内部且连通性较好时,有利于边坡覆绿植物对大气降水的利用。因此,当岩质边坡岩体体裂隙率为1.77%以上且岩体裂隙连通性较好时,则认为适合采用地境再造法对高陡岩质边坡进行覆绿。
2.2 岩体内温度和湿度因子分析
裸露的岩体受到太阳的直接辐射,岩体内温度的变化较快。在植物的生长过程中,低温会造成植物冻害,高温会使其因缺少水分而枯萎,适宜的温度是植物能成活的保障[20]。夏季植物蒸腾作用会消耗大量水分,虽然植物会通过闭合气孔这种生理现象来调节蒸腾作用散失的水分,但若没有持续的水分补给则植物会很快凋萎,可见持续的水分补给也是影响植物能否成活的关键因素[21]。
岩体内的温度一般在夏季和冬季会达到其极限值,且夏季植物蒸腾量最大,冬季岩体内结冰可能会对植物根系造成冻伤乃至死亡,因此只要夏、冬两季岩体内的温度和湿度条件可以满足植物成活,就认为该岩体适合植物生长,故本文选取夏、冬两个季节岩体内温度和湿度的监测数据进行研究。为了探究岩体内的温度及水分分布情况,本文在研究区设计了3个水平监测孔(见图3),监测孔深度设计为1#、2#孔深为5 m,3#孔深为2 m(考虑到成本以及随深度的增加岩体内的温度对植物根系的影响逐渐降低等因素),在每个监测孔内不同深度处安置温湿度仪(见图3中小黑点),并对孔口做一定的防水处理。岩体内的温湿度监测仪器选用DS-1923温湿度纽扣式记录仪,其工作时的温度监测范围为-20℃~+85℃,绝对湿度监测范围为0%~100%RH,及岩体内水汽的饱和状态;温度精度为0.062 5℃,湿度精度为0.04%RH。本次监测试验每个季节监测7 d,筛选后(记录仪器从监测孔取出的时间,并以此时间为节点剔除该时间以后的数据)一共得到91 553组温度、湿度监测数据。
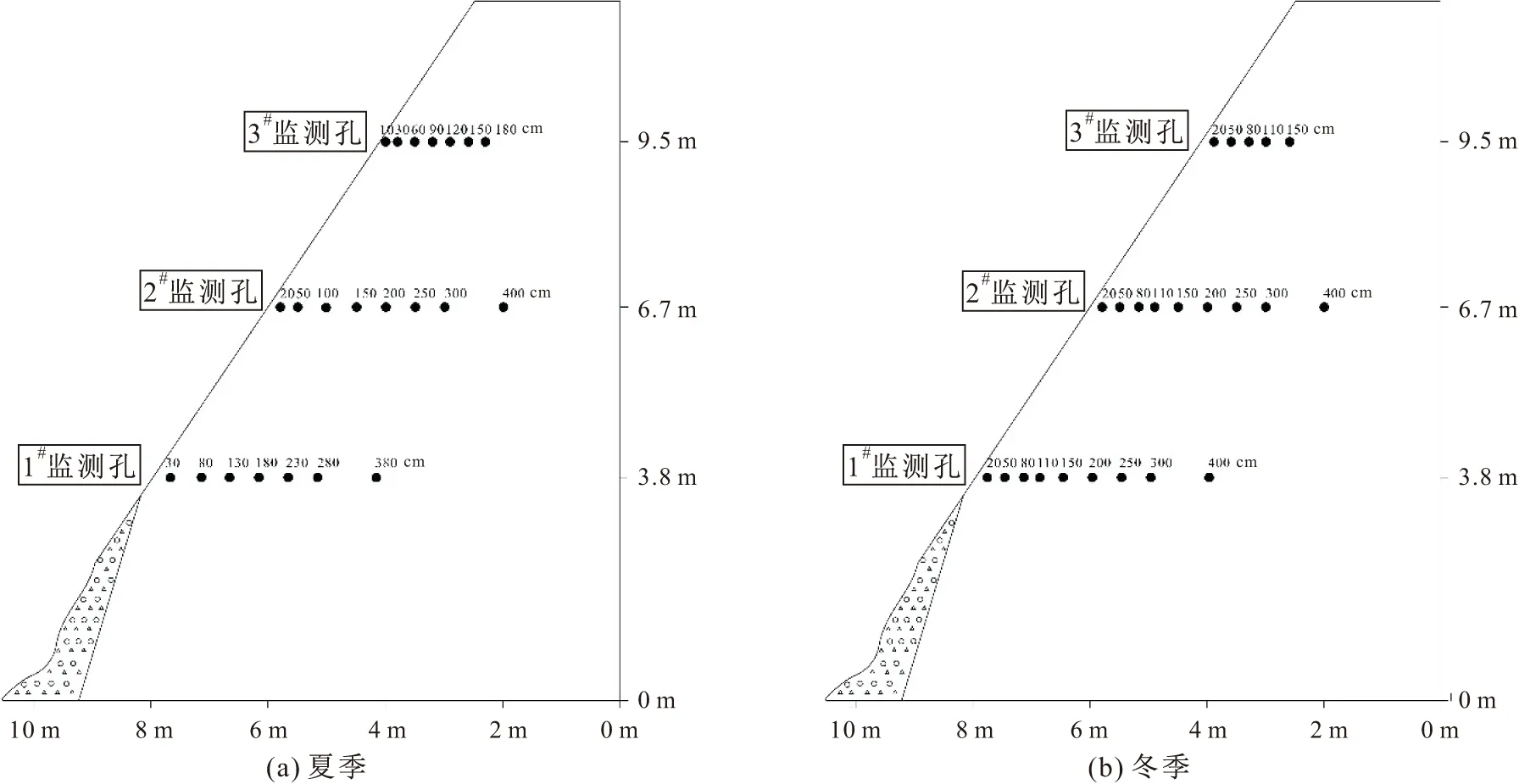
图3 研究区夏、冬季岩体内温度和湿度监测孔位置分布示意图
2.2.1 夏、冬季岩体内温度分布情况分析
为了探究岩体内的温度分布能否满足岩质边坡覆绿植物的生长及成活要求,本文对研究区夏、冬季(夏季监测时间为2017年6月17日至2017年6月23日,冬季监测时间为2018年1月8日至2018年1月15日)各监测孔7 d内的温度监测数据筛选后求其均值,其结果见表2和表3,研究区夏、冬季岩体内平均温度随孔深的变化曲线,见图4和图5。
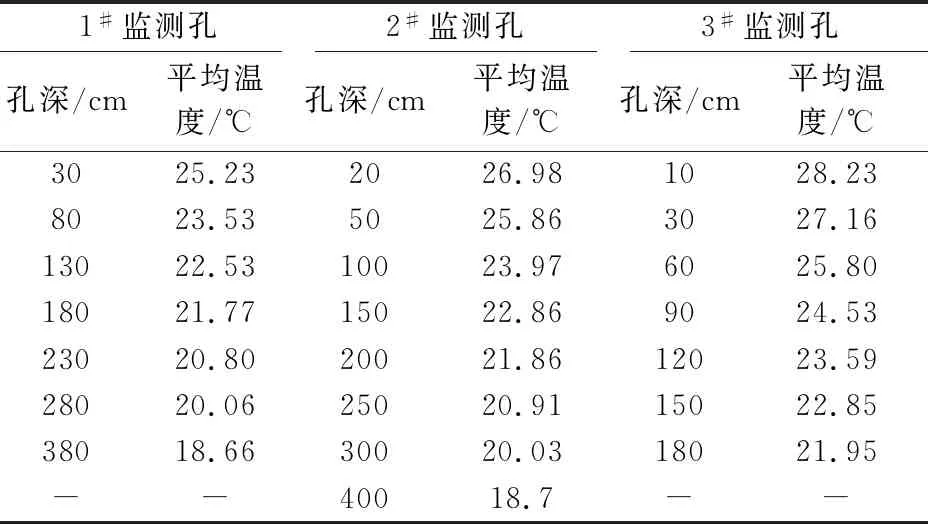
表2 研究区夏季岩体内平均温度的变化情况
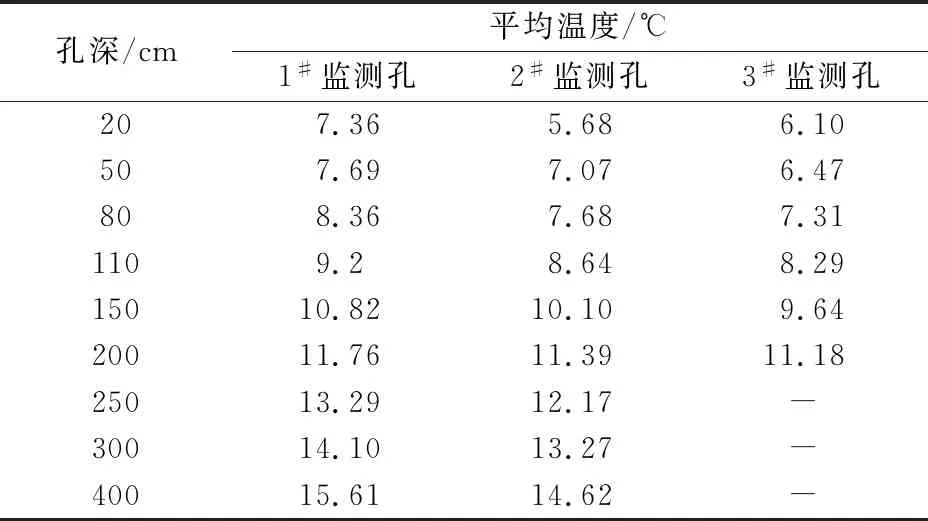
表3 研究区冬季岩体内平均温度的变化情况
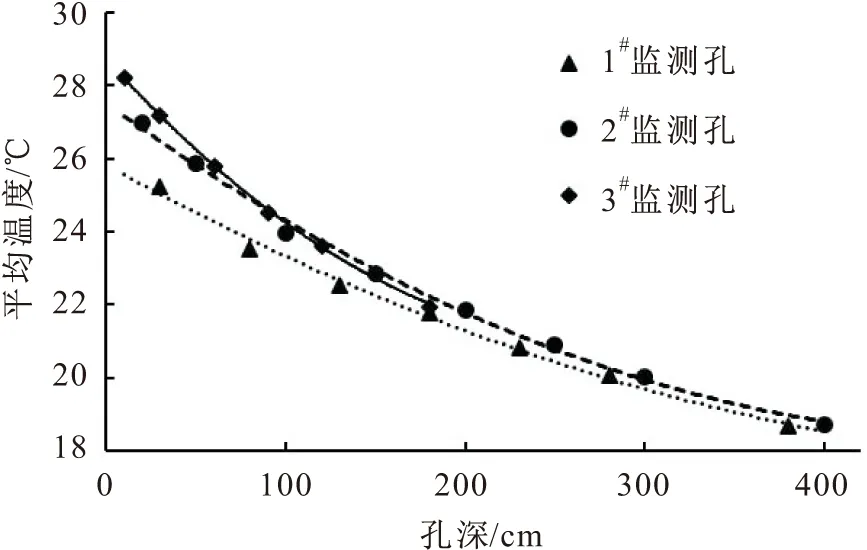
图4 研究区夏季岩体内平均温度随孔深的变化曲线
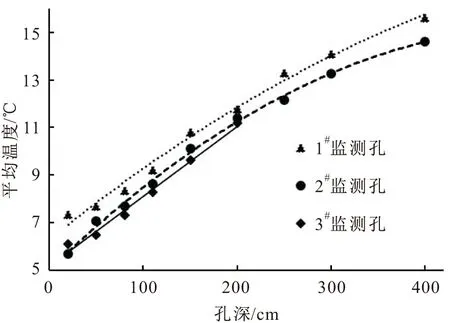
图5 研究区冬季岩体内平均温度随孔深的变化曲线
由表2和图4可见,研究区夏季岩体内20~400 cm深度范围内,随着监测孔深度的增加,岩体内温度逐渐降低,且温度的变化率逐渐降低。这是因为:夏季外界气温高,太阳辐射强烈,岩体外部温度上升快,热量通过岩体与裂隙逐渐向岩体内部传递,传递过程中热量逐渐被消耗,故而随着孔深的增加岩体内温度的变化率逐渐降低;夏季岩体外部温度高,主要是岩体内热量累加的一个过程。
由表3和图5可见,研究区冬季岩体内20~400 cm深度范围内,由岩体内部至岩体外部温度逐渐降低,且温度的变化率逐渐升高。这是因为:冬季外界气温低,太阳辐射弱,岩体外部温度下降快,热量通过岩体与裂隙逐渐向岩体外部传递,传递过程中温度梯度逐渐变大,导致温度变化率升高;冬季岩体外部温度低,主要是岩体内热量耗散的一个过程。
为了研究岩体内温度的昼夜变化情况,本文对岩体种植孔内温度监测数据进行处理后,得到研究区夏、冬季岩体内平均温度的昼夜变化情况,见表4和表5。
由表4和表5可见,研究区夏季岩体内平均温度的昼夜变化不大,平均温度的昼夜变化最大处为岩体内20~30 cm深度处,温度的变化率为0.03℃~0.38℃;冬季岩体内平均温度的昼夜变化最大处为岩体内20 cm深度处,温度的变化率为0.14℃~0.29℃。
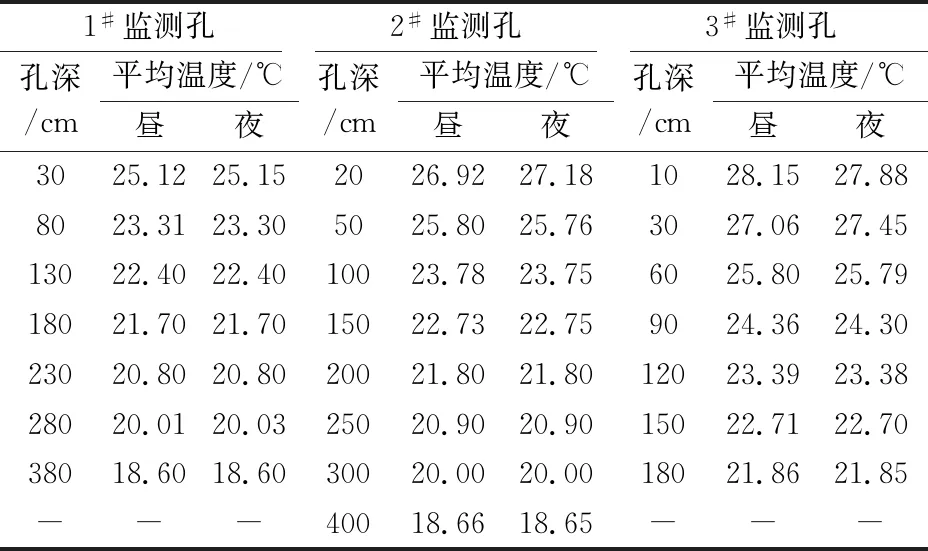
表4 研究区夏季岩体内平均温度的昼夜变化情况
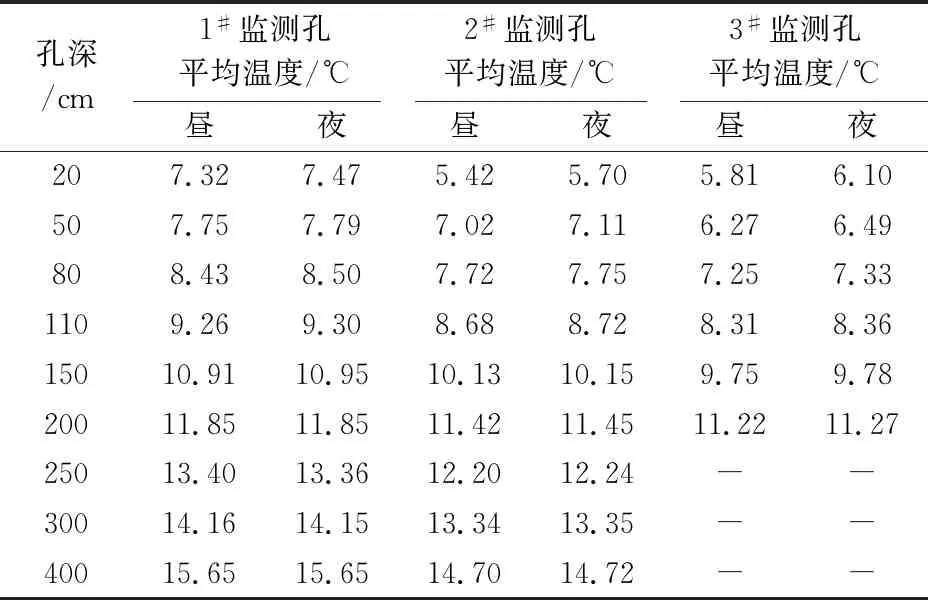
表5 研究区冬季岩体内平均温度的昼夜变化情况
由表2至表5可知,研究区夏季岩体内平均温度的变化范围为18.60℃~28.23℃,冬季岩体内平均温度的变化范围为5.42℃~15.65℃。在温带地区,冬季休眠期多年生植被根系对温度的要求不低于2℃~4℃,夏季生长旺盛时期植物根系对温度的要求温度则不超过30℃[22]。一般植物的最适生长温度为0℃~35℃之间[23],如刺槐的极限耐受温度为-35℃~35℃[24-25]。因此,岩体内的温度情况能够满足植物的成活要求。
2.2.2 夏、冬季岩体内绝对湿度分布情况分析
(1) 夏、冬季岩体内水汽运移情况分析:岩体内部与外界大气构成一个开放系统,不停地进行着物质与能量的交换,即水分与热量的交换。岩体内水汽的运移主要受到水汽分压差的驱动,水汽由绝对湿度(即某温度下的饱和水汽值乘以监测所得对应温度下的相对湿度)大的部位向绝对湿度小的部位运移,且在水汽运移过程中,绝对湿度小的部位水汽受到补给,在接受补给时水汽绝对湿度不断升高,当水汽达到饱和状态且有凝结核存在时,气态水便会凝结形成液态水。为探究研究区岩体内的水分分布状况,对夏、冬季各监测孔位7 d内绝对温度监测数据筛选后求其均值,并绘制研究区夏、冬季岩体内绝对湿度随孔深的变化曲线,见图6和图7。
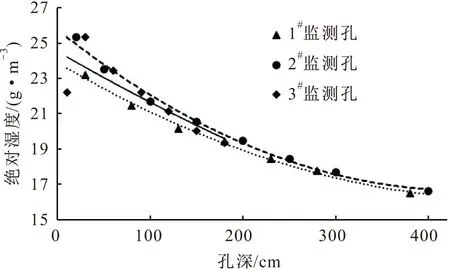
图6 研究区夏季岩体内绝对湿度随孔深的变化曲线
水在裂隙岩体非饱和带中以多种形式存在,有气态水、结合水、孔角毛细水、悬挂毛细水和过路重力水。生长在裂隙岩体内的植物,能吸收利用其内部的少量气态水、毛细水和过路重力水,这些水分可以由大气降水来补给,也可以由岩体内的气态水相互转化来补给。
由图6可见,研究区夏季岩体外部空气的绝对湿度高,岩体内水汽的运移方向主要由空气中的绝对湿度大小来决定,岩体内水汽由绝对湿度大的部位向绝对湿度小的部位运移。因此,夏季岩体内的水汽运移情况是:水汽由岩体外部向岩体内部运移,在水汽运移过程中伴随着温度由岩体外部至岩体内部逐渐降低,水汽在达到饱和且有凝结核存在的条件下会凝结成液态水,其作为岩体的水分补给来源,可供植物吸收与利用。
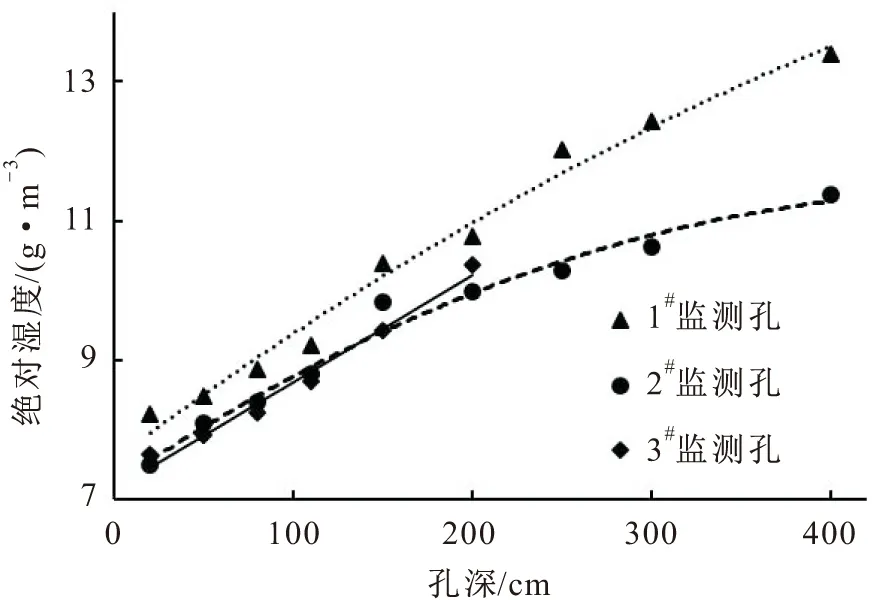
图7 研究区冬季岩体内绝对湿度随孔深的变化曲线
由图7可见,研究区冬季岩体内部空气的绝对湿度高,由岩体内部至岩体外部其空气绝对湿度逐渐降低。因此,冬季岩体内的水汽运移情况是:水汽由岩体内部向岩体外部运移,在水汽运移过程中伴随着温度的降低,水汽会凝结成液态水,可供植物越冬吸收与利用。
(2) 夏、冬季岩体内体积含水率计算:
岩体内的气态水转化为液态水可为覆绿植物提供必要的水分来源。岩体内体积含水率的计算公式为[26]
(1)
式中:WV(T)为岩体内体积含水率(%);β(T)为液、气密度组分比,β(T)=198 857.41×exp(-T/15.793 49)+2 323.367 8;α(T)为液、气质量组分比,α(T)=100 525×exp(-0.052T);T为温度(℃)。
根据公式(1),研究区对夏、冬季岩体内体积含水率进行计算,其计算结果见表6。
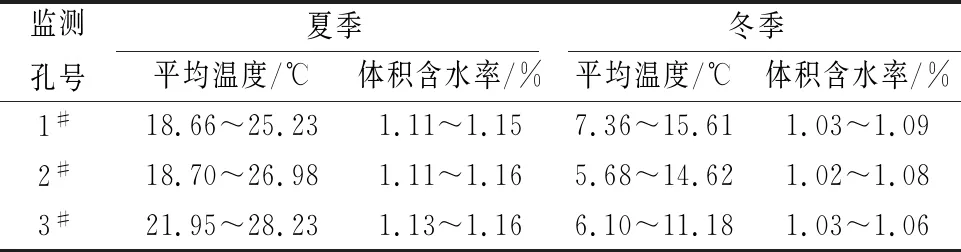
表6 研究区夏、冬季岩体内体积含水率的计算结果
不同种类的植物所具备的抗旱能力差异性较大,已有研究指出当油松和华北落叶松所处的地下生境土壤含水率下降到4.4%和6.9%时苗木的根系活力降低,甚至造成苗木枯死[27];而山杏和小叶锦鸡儿成活所要求的最低土壤含水率分别为0.82%和0.87%[28];1%的土壤含水率是樟子松幼苗的干旱胁迫临界值[29-30]。
由表6可知,研究区裂隙岩体非饱和带岩体内体积含水率在夏、冬季均大于1%,也大于或接近部分植物存活的最小需水量。因此,若植物选择正确,研究区内高陡岩质边坡覆绿工作有很大可能取得成功并可实现后期人工养护较少甚至无人养护。
3 结 论
(1) 在高陡岩质边坡条件下影响植物成活的主要生态因子为岩体裂隙因子、岩体内的温度因子、岩体内的绝对湿度因子。
(2) 裂隙是植物根系在岩体上唯一的生长空间,且是植物生长所需的水分和养分的储存与运移的场所,认为岩体体裂隙率达到1.77%~4.58%时,能满足覆绿植物的生长需求,较适宜覆绿植物成活。
(3) 温度过高或过低都会影响植物的生长发育,植物只有在其适宜的温度范围内才能正常生长。岩体内夏、冬季的极限温度分别约为28℃、5℃,均能满足植物的生长。
(4) 研究区裂隙岩体非饱和带岩体内的体积含水率均大于1%,大于某些耐旱植物成活对其所处地下生境土壤含水率的要求。同时,裂隙岩体内部大量的水汽会做规律性的迁移运动,并伴随着水、汽的相互转化,且随着气态水向液态水转化过程的不断进行,就会有液态水产生,可对裂隙岩体非饱和带水分进行补充,这成为非饱和带裂隙岩体重要的水分来源。
猜你喜欢
成都信息工程大学学报(2022年4期)2022-11-18 07:31:56
昆明医科大学学报(2021年5期)2021-07-22 07:32:54
水利规划与设计(2020年1期)2020-05-25 08:01:30
智能城市(2018年7期)2018-07-10 08:30:08
江西建材(2018年2期)2018-04-14 08:00:54
中国眼镜科技杂志(2018年1期)2018-02-08 02:15:28
高原山地气象研究(2016年1期)2016-11-10 06:05:53
水科学与工程技术(2016年6期)2016-02-27 13:29:15
电影新作(2016年1期)2016-02-27 09:16:48
广东海洋大学学报(2015年3期)2015-12-22 10:05:28
