
日照紫菜养殖海域营养盐的时空分布特征及其与浮游植物群落结构的相关性分析*
2019-09-27 01:20梁洲瑞孙藤芹汪文俊刘福利梁启迪孙修涛王飞久
渔业科学进展 2019年5期
梁洲瑞 孙藤芹 汪文俊 刘福利 曹 原 梁启迪 孙 昕 孙修涛 王飞久
日照紫菜养殖海域营养盐的时空分布特征及其与浮游植物群落结构的相关性分析*
梁洲瑞1,2孙藤芹3汪文俊1,2①刘福利1,2曹 原3梁启迪1孙 昕1孙修涛1,2王飞久1
1. 中国水产科学研究院黄海水产研究所 农业农村部海洋渔业可持续发展重点实验室 青岛 266071;2. 青岛海洋科学与技术试点国家实验室海洋渔业科学与食物产出过程功能实验室 青岛 266071;3. 上海海洋大学水产与生命学院 上海 201306)
2016年11月~2017年3月对山东日照阜鑫渔港紫菜(sp.)养殖海域开展调查,分析该海域浮游植物群落结构和营养盐的时空分布特征及其相关性。结果显示,调查海区中无机氮(DIN)、磷酸盐(PO43–-P)及硅酸盐(SiO32–-Si)浓度均表现为自近岸海区到外海区逐渐降低的趋势;硝酸盐、SiO32–-Si、PO43–-P、DIN和溶解有机氮(DON)等营养盐的浓度均因月份的不同具有显著差异;调查海区营养水平总体为中营养型,浮游植物丰度为(0.049~3.031)×104cells/L,浮游植物的生长受控于SiO32–-Si和PO43–-P的几率较大;该海区共检出硅藻门27属37种,甲藻门7属8种,金藻门1属1种,主要优势种为骨条藻(sp.)、角毛藻(sp.)、圆筛藻(sp.)等;盐度的大幅降低和丰富的营养盐、氨氮(NH4+-N)和PO43–-P浓度的显著变化可能分别是导致1、3月浮游植物多样性指数显著降低的主要原因;调查海区NH4+-N浓度和DIN/P值与浮游植物多样性指数均呈负相关关系,其中,非紫菜养殖区的负相关程度达到显著水平,而养殖区的负相关程度不显著,表明条斑紫菜()养殖可能有利于降低该海域的NH4+-N水平和DIN/P值、提高浮游植物群落结构稳定性及物种多样性,从而有利于防止赤潮的发生。研究结果可为该海域环境保护、赤潮防治以及水产养殖活动的开展等提供基础数据。
紫菜养殖;浮游植物;营养盐;分布
山东日照阜鑫渔港海域近几年开始规模化养殖紫菜(sp.),目前,养殖规模已迅速扩大到了200 hm2,逐渐成为江苏以北的紫菜主产区。该养殖海区西边为巨峰河入海口(距离养殖区约2 km),北边有傅疃河入海口(距离养殖区约6 km)。受日照市涛雒地区工业生产及人类活动的影响,巨峰河和傅疃河的河水受到了较严重的污染,在河流入海口处检测出大量N、P等易引发水体富营养化的污染物(吴艳红等, 2004; 刘久潭等, 2017; 刘昕宇等, 2017)。随着日照市沿海地区经济的快速发展,阜鑫渔港附近海区的陆源入海污染导致该海域的富营养化问题日益严重,该海域的海洋生态环境将面临更严峻的挑战,亟需对其开展海水环境状况的评价与水体富营养化的防治工作。
浮游植物作为海洋生态系统中重要的初级生产力,其群落结构是海域水体理化状态和生态环境中其他物种相互作用的结果,其群落结构的时空变化可反映海域环境的动态变化,成为识别海域环境状况、评价海洋生态系统是否健康的重要指标(孙雪梅等, 2013)。丰富度(Richness)、均匀度(Evenness)、香农指数(Shannon index)、辛普森指数(Simpson index)等多样性指数都是反映浮游植物群落中的种类数和种间数量分配的量度(黄邦钦等, 1995),多样性指数越高,海域系统越趋于稳定。浮游植物多样性指数与营养盐结构密切相关(王育红等, 2014; 孙雪梅等, 2016)。而大型海藻和浮游植物之间对阳光、营养盐和生长空间均存在竞争,大型海藻养殖活动的开展可影响海水中营养盐的组成及浮游植物的群落结构(李宵, 2009; 王洪斌等, 2014)。另外,规模化的紫菜养殖生产对海水中营养盐的吸收作用可能是解决该海域富营养化问题的有效途径。因此,本研究在日照阜鑫渔港海域的紫菜养殖期间,对该海域的紫菜养殖区及其附近的非养殖区开展多个航次调查,分析该海域营养盐和浮游植物的时空分布特征及其相关性,并对该海域的富营养化等级进行评价,为该海域生态环境保护、污染防治与决策及海藻养殖或海洋动物养殖等水产养殖活动的开展等提供依据。
1 材料与方法
1.1 采样站位
本调查于2016年11月29日、2017年1月12日、2月23日、3月21日在日照阜鑫渔港附近紫菜养殖区及其周边采集水样,以紫菜养殖区为中心,分为近岸区、紫菜养殖区和外海区3个区域,每个区域各设6个站位,共18个调查站位见图1。
1.2 采样及分析方法
调查指标包括温度、盐度、硝酸氮(NO3–-N)、氨氮(NH4+-N)、亚硝酸氮(NO2–-N)、无机磷酸盐(PO43–-P)、硅酸盐(SiO32–-Si)、总溶解态氮(TDN)等环境参数以及浮游植物丰度。使用有机玻璃采水器采集表层水样,待测浮游植物水样用鲁哥氏液固定,待测营养盐水样过滤后进行冰冻保存。
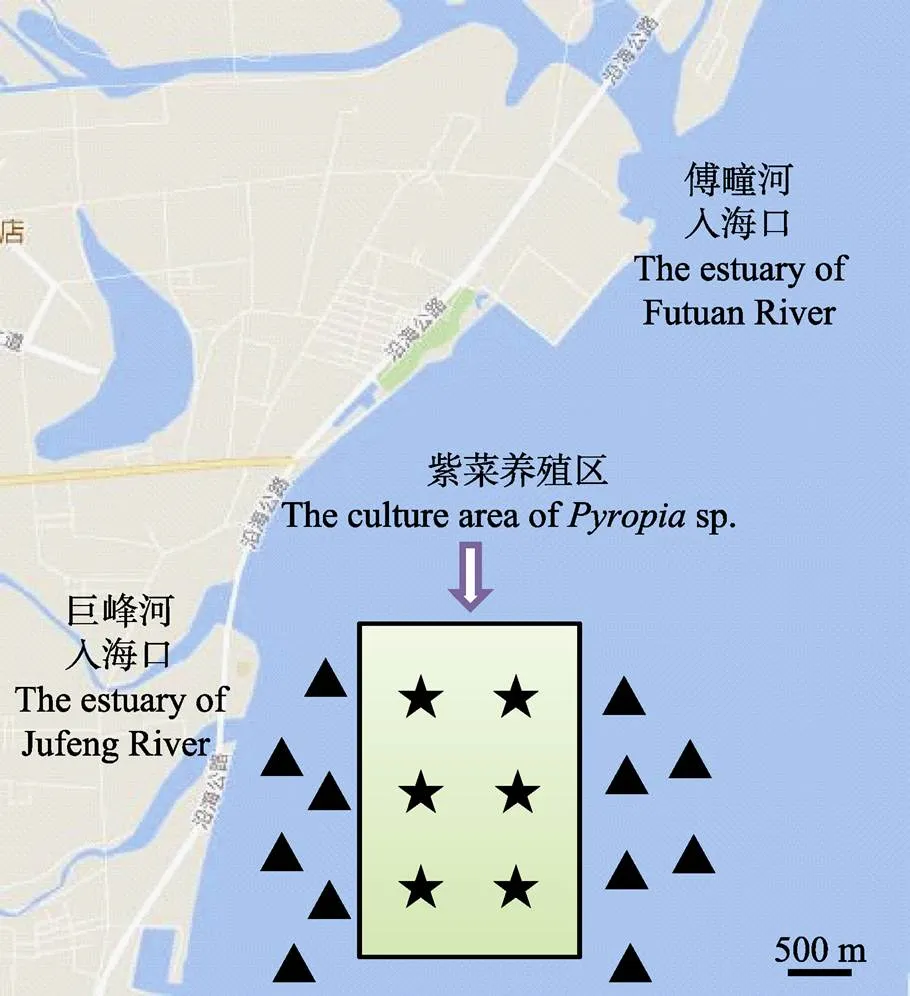
图1 采样点示意(以★、▲表示)
使用YSI多参数水质分析仪(Pro plus,美国)现场测定温度、盐度等环境参数。按照《海洋监测规范》(GB17378.7–2007)进行浮游植物的丰度和种类的分析。营养盐浓度的检测方法参照《海洋调查规范》(GB/T12763.4–2007),使用AA3连续流动分析仪测定。NO3–-N采用重氮–偶氮法测定;NH4+-N采用靛酚蓝法测定;NO2–-N采用铜镉柱还原法测定;SiO32–-Si采用硅钼蓝法测定;PO43–-P采用抗坏血酸还原磷钼蓝法测定。TDN采用总有机碳分析仪(TOC-VCPH,日本岛津)进行分析测定。溶解无机氮(DIN)浓度为NO3–-N、NH4+-N和NO2–-N浓度总和;溶解有机氮(DON)浓度等于TDN浓度减去DIN浓度。
1.3 数据处理
采用SPSS 19.0统计软件进行单因素方差分析(One-way ANOVA)、Duncan多重比较及Pearson相关性分析(<0.05为差异显著,<0.01为差异极显著),使用Excel软件绘制图形。采用PRIMER软件分析得到浮游植物的丰富度、均匀度、香农指数、辛普森指数等多样性指数。浮游植物优势度计算公式为:
=n/×f
式中,n为第种浮游植物的丰度;f为该种在各站位的出现频率;为该海域浮游植物丰度的总和。以≥0.02为优势种划分标准。
1.4 富营养化评价标准
为研究本调查海域的富营养化程度,结合不同研究中的富营养化等级评价标准(沈韫芬等, 1990; 李清雪等, 1999; 安斐等, 2008),划分富营养化等级标准见表1。
表1 浮游植物丰度、多样性指数及总溶解态氮与水体营养水平的关系
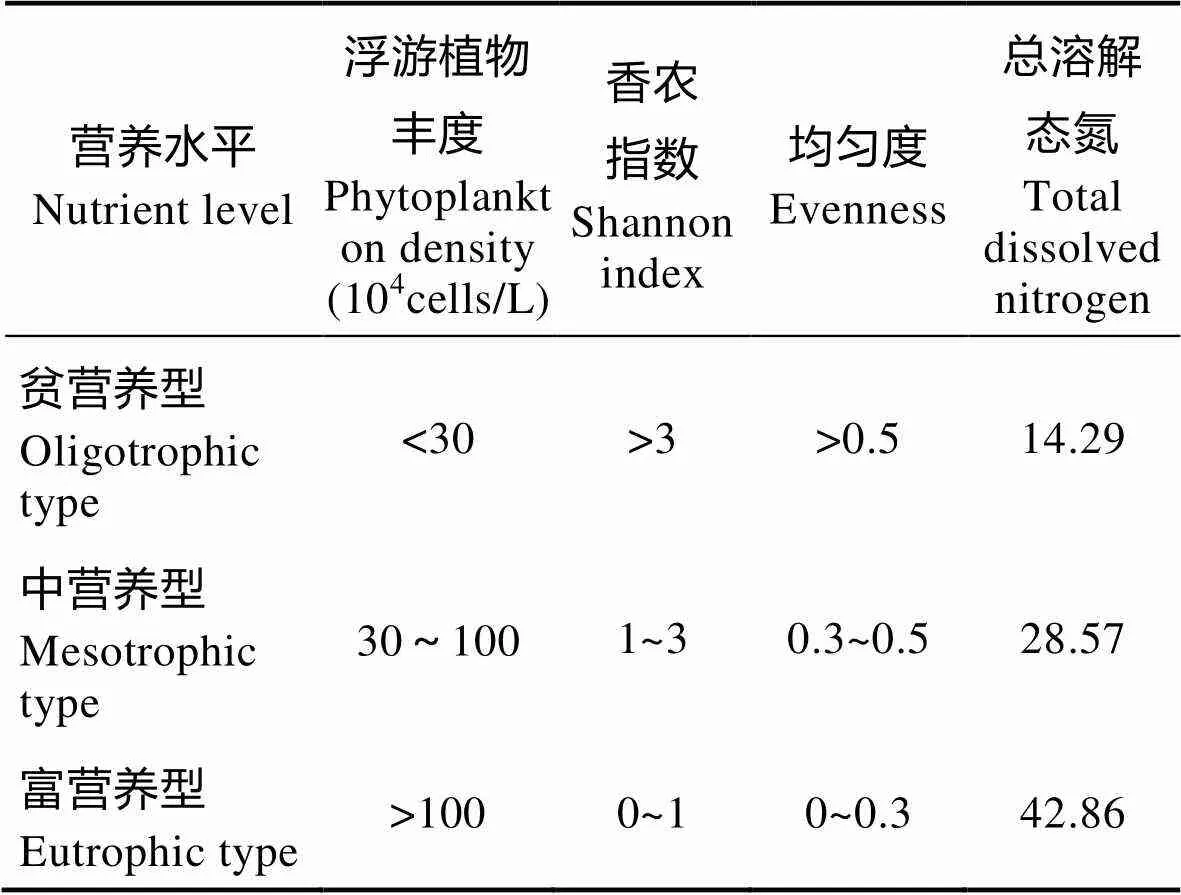
Tab.1 The relationship between phytoplankton density, diversity index, total dissolved nitrogen and nutrient level
本研究使用1、2、3分别代表贫营养、中营养、富营养型进行生物学评价模型拟定。将浮游植物丰度、香农指数、均匀度、TDN等要素的实测值与表中的标准相比较,确定各单项指标,然后,取各要素单项指标的平均值,得到各站位的综合营养等级指标。
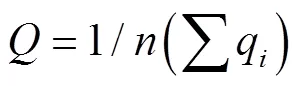
式中,为综合指标,q为单项指标数。值大于2.5为富营养水平,1.5~2.5为中营养水平,小于1.5为贫营养水平(李清雪等, 1999)。
1.5 营养盐限制因子判断标准
为研究本调查海域表层水体无机营养盐结构对浮游植物生长的可能限制因子,本实验结合不同研究中的无机营养盐限制因子判断标准(Justić, 1995; 史乾, 2012),营养盐限制因子的判断见表2。
表2 氮、磷、硅无机营养盐作为单一限制因子的判断标准

Tab.2 The judgment standard as a single limiting factor for inorganic nutrient of nitrogen, phosphorus and silicon
2 结果
2.1 温度和盐度的时空变化
从图2可以看出,调查海域的水温在1、2月较低(图2a),盐度从近岸海区到外海海区呈递增的趋势。方差分析结果显示,盐度因海区的不同及月份的不同均有显著性差异(<0.05)。多重比较分析结果显示,外海区的盐度显著高于近岸海区和养殖区(<0.05); 1月盐度较低,与其他月相比差异显著(<0.05);11月盐度较高,与其他月相比差异显著(<0.05) (图2b)。
2.2 氮、磷、硅营养盐的时空分布特征
方差分析结果显示,SiO32–-Si和PO43–-P浓度因月份的不同均呈显著性差异(<0.05);1月,不同海区的PO43–-P浓度差异显著(<0.05);2、3月,不同海区的SiO32–-Si浓度均呈显著性差异(<0.05)。多重比较分析结果显示,11月、翌年1月的SiO32–-Si浓度较高,与2、3月相比均差异显著(<0.05);2月养殖区和 3月近岸海区的SiO32–-Si浓度与当月外海区相比均差异显著(<0.05) (图3a)。从图3b可以看出,11月和翌年2月的PO43–-P浓度均较高,与3月相比均差异显著(<0.05);11月近岸海区的PO43–-P浓度较高,与外海区相比差异显著(<0.05);1月近岸海区和养殖区的PO43–-P浓度均较高,与外海区相比均差异显著(<0.05)。Pearson相关性分析表明,SiO32–-Si浓度与NO3–-N、NO2–-N和DIN浓度均呈极显著的正相关关系(<0.01),而PO43–-P浓度只与NO2–-N呈显著的正相关关系(<0.05);NH4+-N浓度和其他营养盐浓度之间均无显著差异(>0.05)。
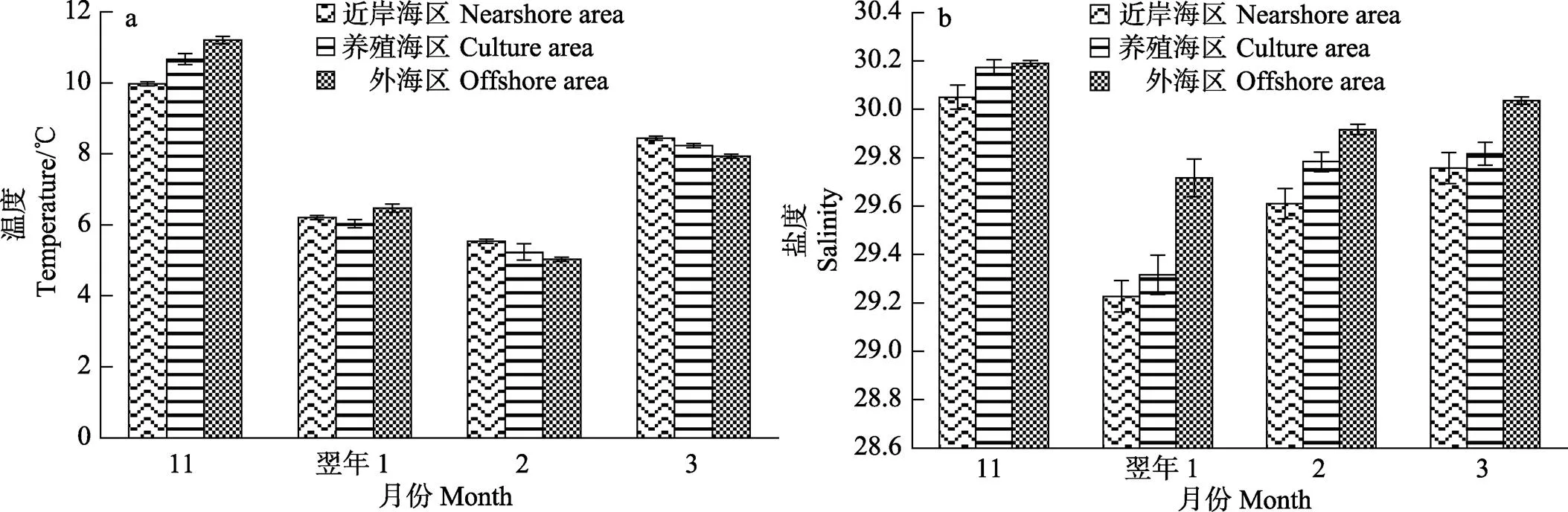
图2 不同月份各海区的温度(a)和盐度(b)
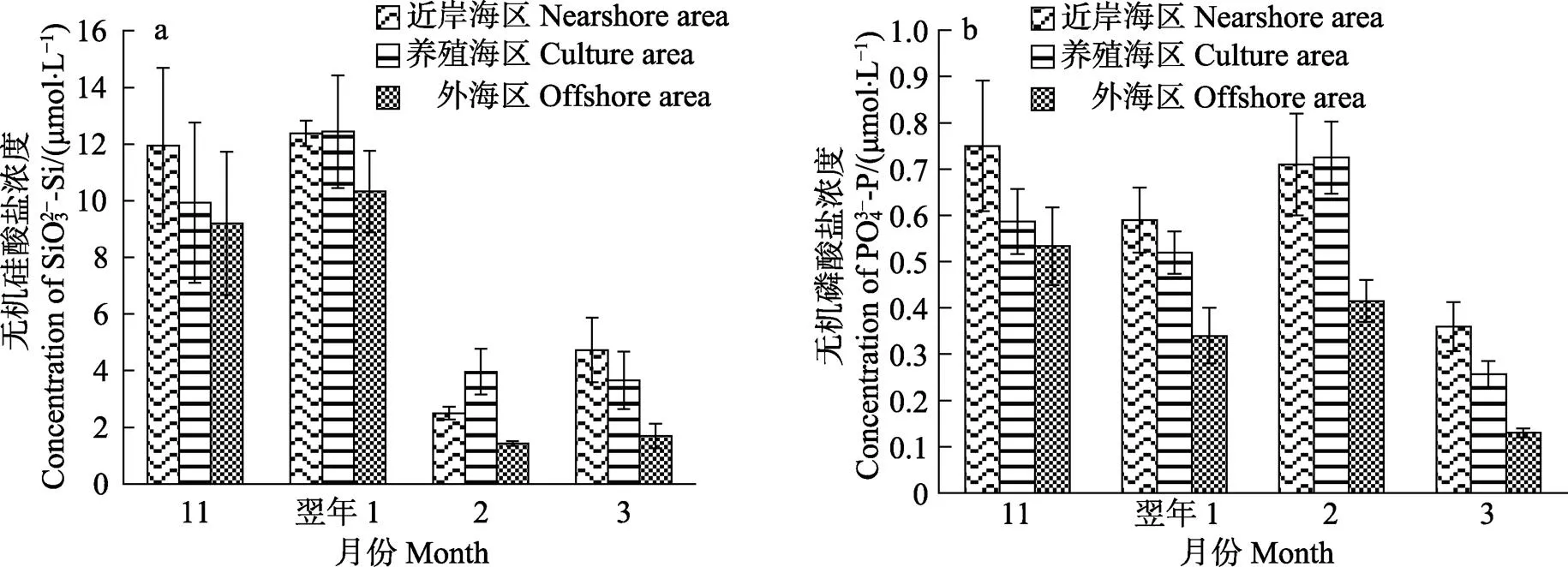
图3 不同月份各海区的硅酸盐(a)和磷酸盐(b)浓度
该海域DIN的组成中,NO3–-N、NH4+-N和NO2–-N平均占比分别为79.76%、11.67%和8.57%(图4)。方差分析结果显示,不同月份的NO3–-N浓度呈显著性差异(<0.05),而NH4+-N和NO2–-N均无显著性差异(>0.05)。多重比较分析显示,11月和翌年1月的NO3–-N浓度均较高,与2、3月相比均呈显著性差异(<0.05);2月的NH4+-N浓度较低,与3月相比差异显著(<0.05);3月的NO2–-N浓度较低,与1月相比差异显著(<0.05)。
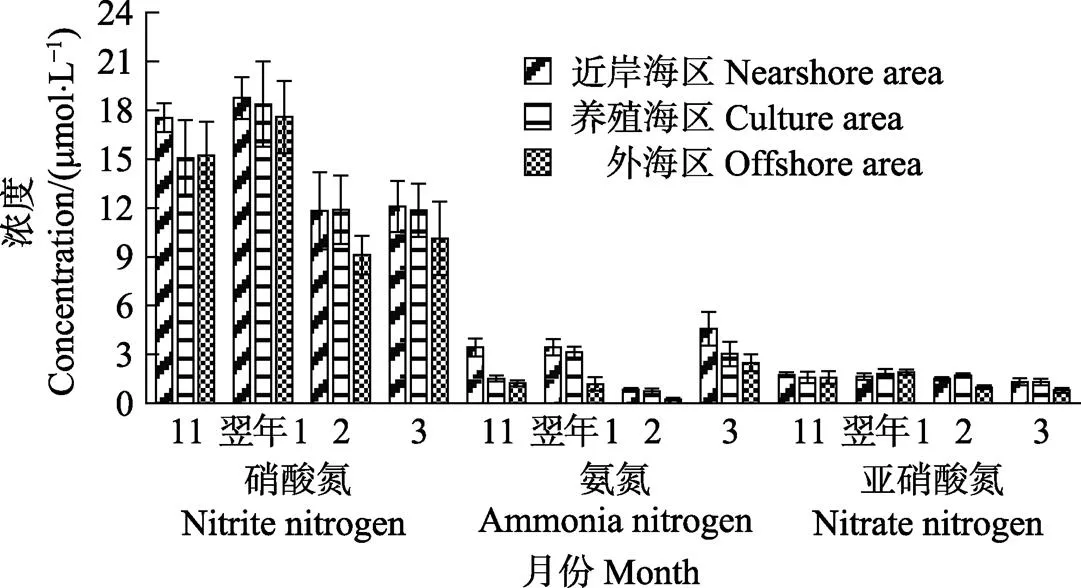
图4 不同月份各海区的溶解无机氮(包括硝酸氮、氨氮和亚硝酸氮)浓度
该海域中NO3–-N、NH4+-N和NO2–-N浓度均表现为自近岸海区到外海区逐渐降低的趋势。方差分析结果显示,每个月的NO3–-N浓度因海区的不同均无显著性差异(>0.05);2016年11月和翌年1月,不同海区的NH4+-N浓度均呈显著性差异(<0.05);2、3月,不同海区的NO2–-N浓度均呈显著性差异(<0.05)。多重比较分析结果显示,11月近岸海区的NH4+-N浓度较高,与养殖区和外海区相比差异显著(<0.05);1月近岸海区和养殖区的NH4+-N浓度较高,与外海区相比均呈显著性差异(<0.05);2、3月的近岸海区和养殖区NO2–-N浓度均较高,与外海区相比差异显著(<0.05)。
方差分析结果显示,不同月份的DIN及DON浓度均呈显著性差异(<0.05)。多重比较分析结果显示,11月和翌年1月的DIN浓度较高,与2月相比均呈显著性差异(<0.05);11月的DON浓度均较高,与其他月份相比均呈显著性差异(<0.05) (图5)。
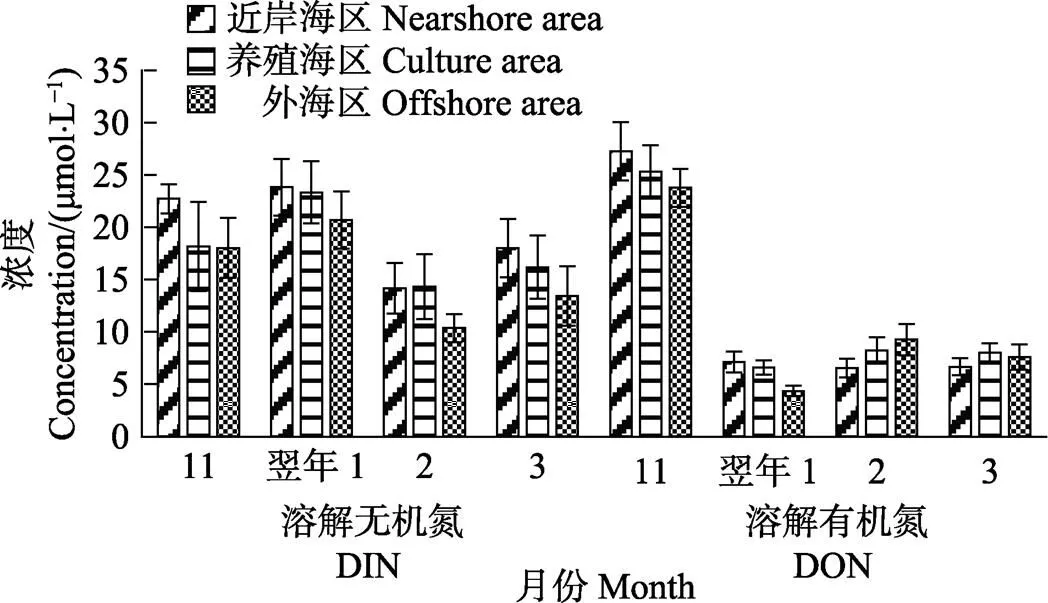
图5 不同月份各海区的溶解态氮浓度
该海域中DIN及DON的浓度一般表现为自近岸海区到外海区逐渐降低的趋势。方差分析结果显示,每个月的DON浓度因海区的不同均无显著性差异(>0.05);在2月,不同海区的DIN浓度呈显著性差异(>0.05)。多重比较分析结果显示,2月近岸海区和养殖区的DIN浓度较高,与外海区相比差异显著(<0.05)。
该海域的DIN/P值在调查期间一直维持在较高的水平,11月的DIN/P值为30.3~33.8,翌年1月的DIN/P值为40.4~60.8,2月的DIN/P值为19.9~24.9,3月的DIN/P值为50.0~103.3。
2.3 浮游植物的种类组成与分布
2.3.1 浮游植物的种类 从表3可以看出,调查海区水样中观察到浮游植物共3门35属46种。群落组成中,硅藻门(Bacillariophyta)的种类和数量最多,共27属37种,其中,角毛藻属()种类最多,检出5种,其次是菱形藻属(),检出3种。除此之外,还观察到甲藻门(Pyrrophyta)7属8种,金藻门(Chrysophyta) 1属1种。
2.3.2 浮游植物优势种及其优势度 从表4可以看出,调查海域中浮游植物主要优势种(本文取优势度大于0.1的浮游植物作为主要优势种)有骨条藻(sp.)、角毛藻(sp.)、圆筛藻(sp.)、柔弱几内亚藻()、宽角斜纹藻()、具槽帕拉藻()、布氏双尾藻()、辐射圆筛藻()、叉状角藻()等。调查海域每个月的浮游植物优势种不一致,调查时间范围内基本以硅藻为优势种,其中,只有2016年11月的养殖区同时出现硅藻和甲藻(梭形角藻和叉状角藻)的优势种。2017年的1~3月,各个海区均出现大量的骨条藻(1月骨条藻优势度为0.5125~0.8330,2月骨条藻优势度为0.3992~0.5973, 3月骨条藻优势度为0.6719~0.8014)。2017年3月, 3个不同海区的浮游植物优势种均为骨条藻和角毛藻。
2.3.3 浮游植物的丰度 从图6可以看出,浮游植物的细胞丰度为(0.049~3.031)×104cells/L。方差分析结果显示,不同月份的浮游植物丰度呈显著性差异(<0.05);在11月和翌年3月,不同海区的浮游植物丰度均无显著性差异(>0.05);而在1、2月,不同海区的浮游植物丰度均呈显著性差异(<0.05)。11月的浮游植物丰度极显著小于其他月份(<0.01)。近岸海区的浮游植物丰度整体上小于养殖海区和外海区的浮游植物丰度。Pearson相关性分析表明,浮游植物丰度与温度呈极显著的负相关关系(<0.01)。
表3 调查海区浮游植物的种类
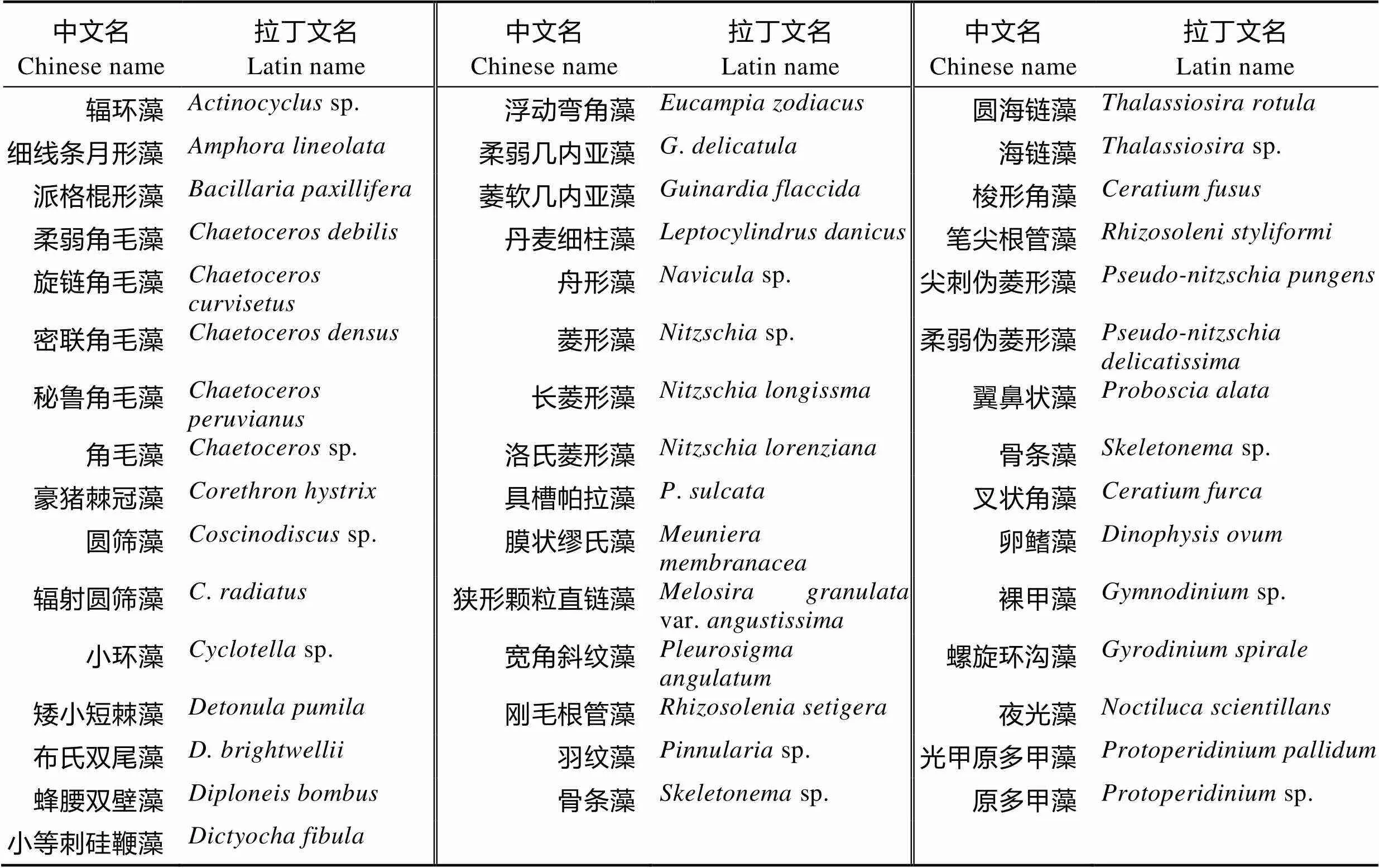
Tab.3 Species of phytoplankton at the surveyed sea area
表4 不同月各海区的浮游植物优势种及其优势度

Tab.4 The dominant species of phytoplankton and its dominance at different sea areas in different months
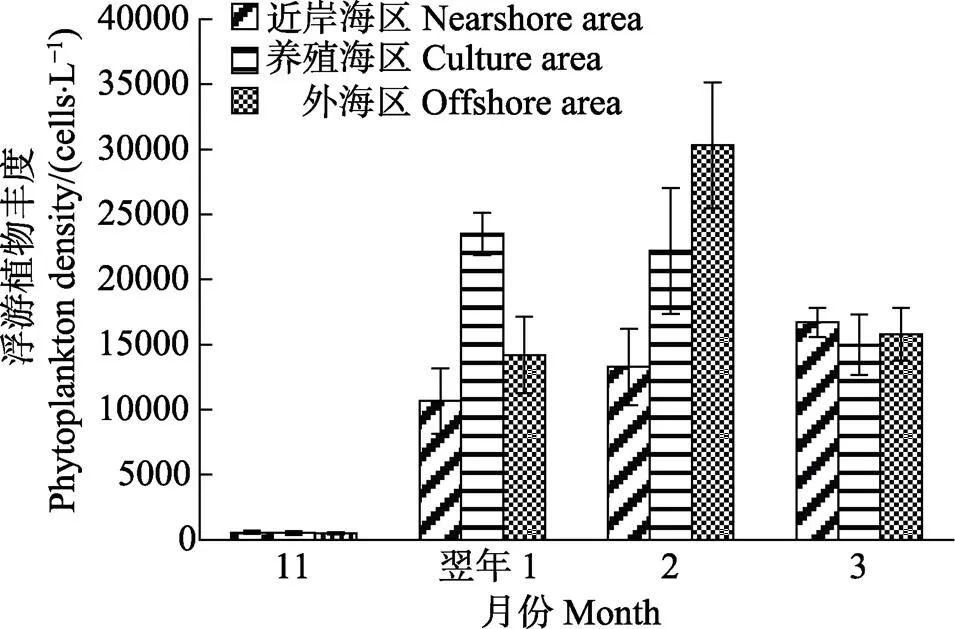
图6 不同月份各海区的浮游植物丰度
2.3.4 浮游植物的多样性 方差分析结果显示,不同月份的浮游植物丰度、均匀度、香农指数和辛普森指数均呈显著性差异(<0.05);不同海区的浮游植物丰度无显著性差异(>0.05);对于11月和翌年2月,不同海区的浮游植物均匀度、香农指数和辛普森指数均无显著性差异(>0.05);而1、3月,不同海区的浮游植物均匀度、香农指数和辛普森指数均呈显著性差异(<0.05) (图7)。浮游植物均匀度、香农指数和辛普森指数在2016年11月和翌年2月较高,与1、3月相比均呈显著性差异(<0.05)。
2.3.5 富营养化评价 根据1.4的富营养化评价标准,浮游植物丰度、多样性指数(香农指数和均匀度)、TDN等指标的营养水平得分见表5。从表5可以看出,该海域在2月的营养水平为贫营养型,其他月份为中营养型。
2.3.6 营养盐限制因子 从表6可以看出,该海域表层水体中浮游植物生长受控于N、P、Si的单一营养盐限制因子,出现几率较高的是SiO32–-Si(25.0%),NO3–-N次之(16.7%),DIN的限制几率为0。当考虑营养盐最低阈值时,受DIN、PO43–-P和SiO32–-Si的限制几率均为0。该海域11月和翌年3月单一营养盐限制因子出现的几率均为0。
2.3.7 营养盐浓度与浮游植物群落结构之间的相关性分析 从表7可以看出,NO3–-N浓度与近岸海区浮游植物丰度呈显著负相关关系(<0.05),而与3个调查海区的浮游植物多样性指数均无显著的相关性(>0.05)。SiO32–-Si浓度与近岸海区及外海区浮游植物丰度均呈显著的负相关关系(<0.05),而与3个调查海区的浮游植物多样性指数均无显著的相关性(>0.05)。PO43–-P浓度与近岸海区浮游植物丰度也呈显著的负相关关系(<0.05),且与3个调查海区的浮游植物多样性指数(丰度、均匀度、香农指数和辛普森指数)均呈正相关关系,其中,与香农指数的正相关程度均达到显著水平(<0.05)。而溶解无机营养盐(NO3–-N、NH4+-N、SiO32–-Si、PO43–-P)与养殖区浮游植物丰度的相关程度均没有达到显著水平(>0.05)。DON浓度与3个调查海区的浮游植物丰度均呈显著的负相关关系(<0.05)。
NH4+-N浓度和DIN/P的值与3个调查海区的浮游植物均匀度、香农指数和辛普森指数均呈负相关关系,其中,对于浮游植物香农指数与NH4+-N浓度的负相关程度,近岸海区与外海海区均达到显著水平(<0.05),而养殖区没有达到显著水平(>0.05);对于浮游植物均匀度和辛普森指数分别与NH4+-N浓度的负相关程度,养殖区均小于近岸海区与外海海区;对于浮游植物均匀度、香农指数和辛普森指数分别与DIN/P值的负相关程度,近岸海区与外海海区均达到显著水平(<0.05),而养殖区均没有达到显著水平(>0.05)。
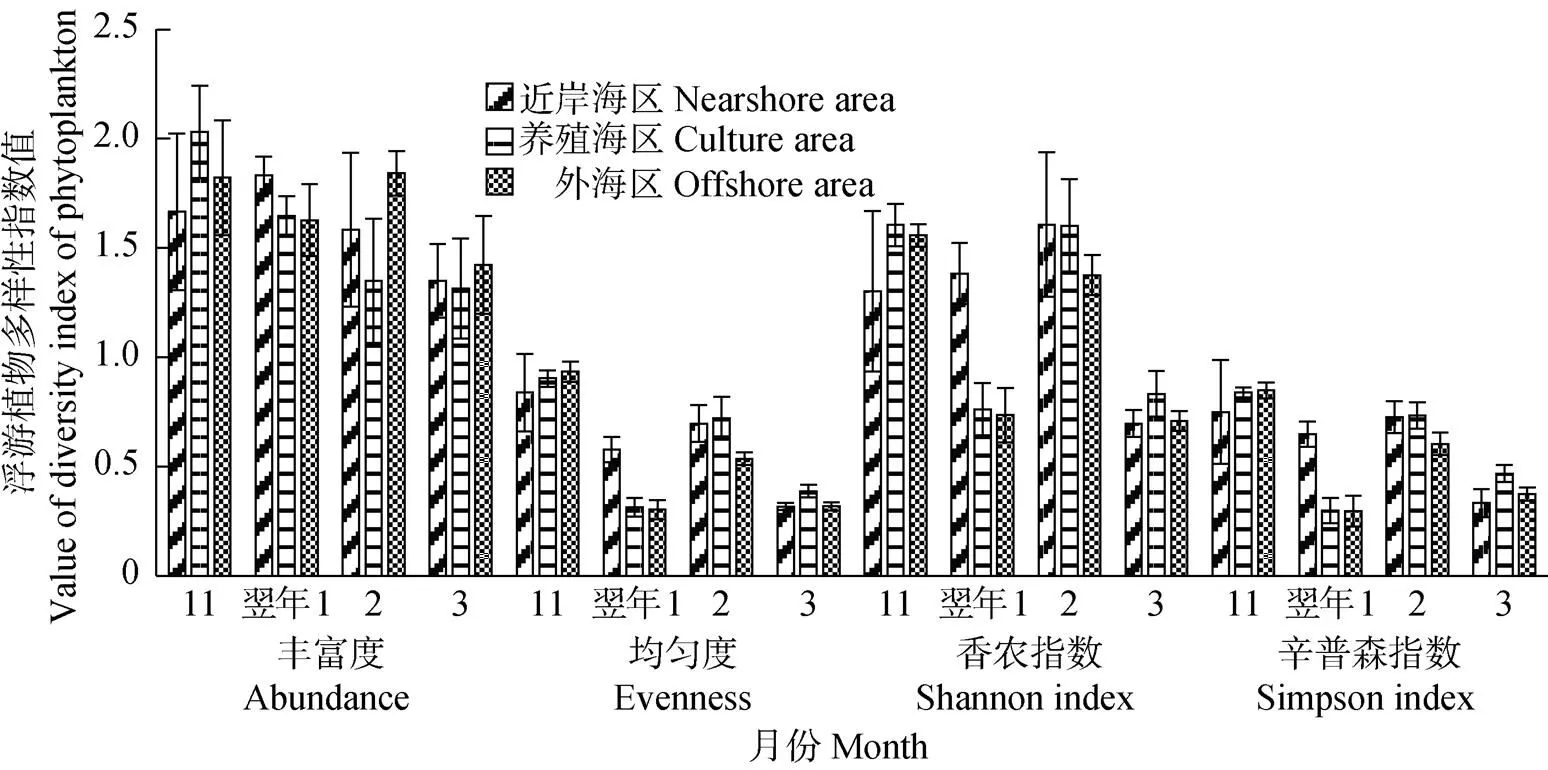
图7 不同月份各海区的浮游植物多样性指数
表5 不同月份各海区的营养水平得分
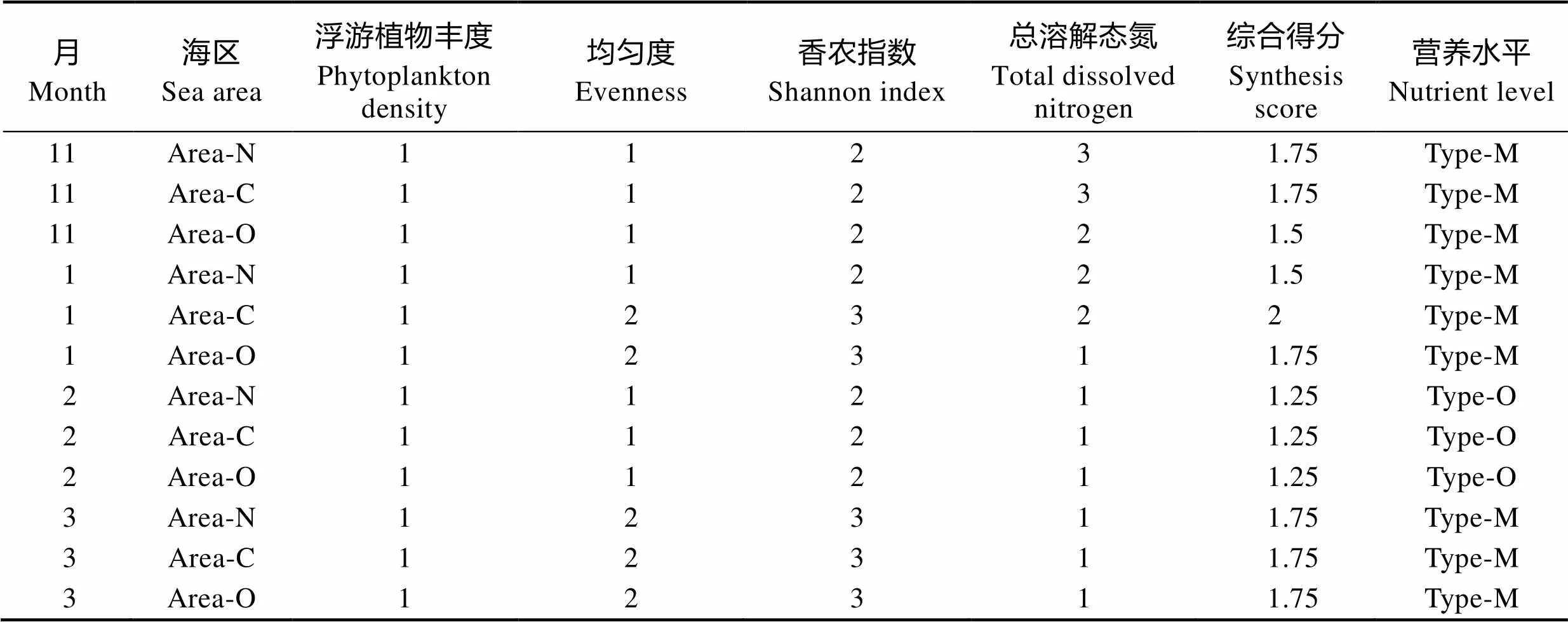
Tab.5 The score of nutrition level at different sea areas in different months
注:Area-N、Area-C和Area-O分别为近岸区、养殖区和外海区。Type-O和Type-M分别为贫营养型和中营养型,同表7
Note: Area-N, Area-C and Area-O represent nearshore area, culture area and offshore arearespectively. Type-O and Type-M represent oligotrophic type and mesotrophic type respectively, the same as Tab.7
表6 调查海域表层水体氮、磷、硅无机营养盐作为单一限制因子出现的几率
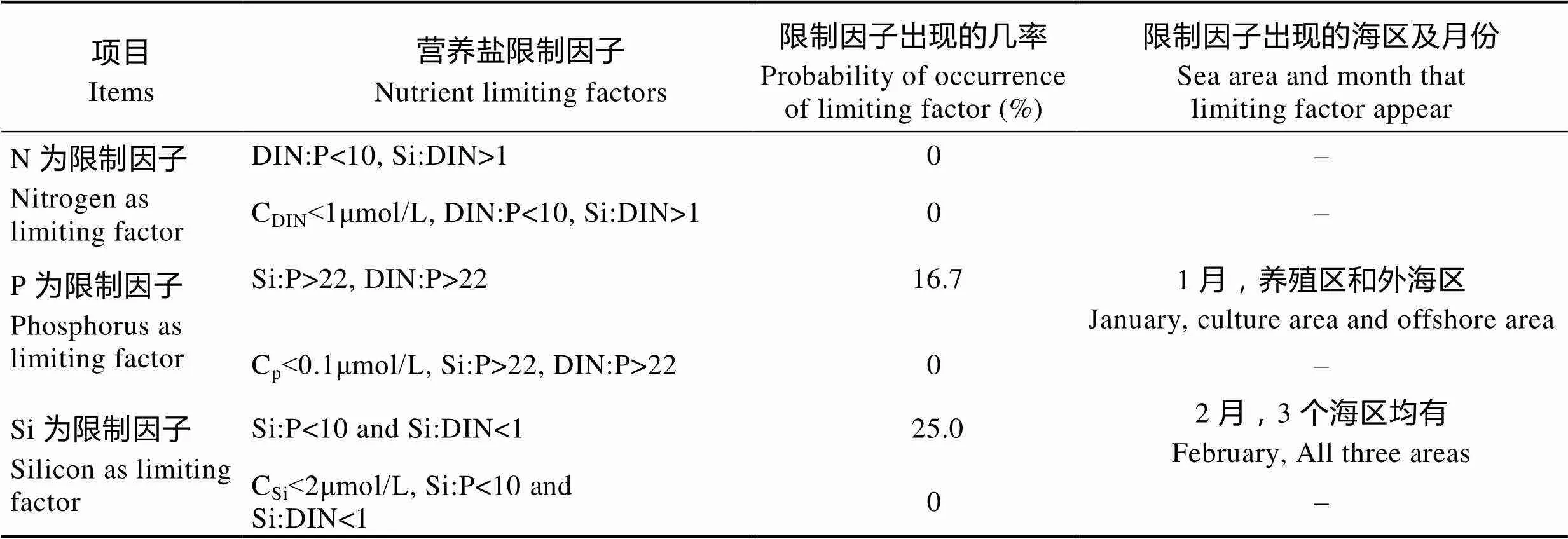
Tab.6 The probability of occurrence as a single limiting factor for inorganic nutrient of nitrogen, phosphorus and silicon in the surface water at the surveyed sea area
注:限制因子出现的几率(%)=出现限制因子的站位数量/总站位数量×100
Note: Probability of occurrence of limiting factor (%)=Number of positions that limiting factor appear/Total number of positions × 100
3 讨论
调查海域的盐度从近岸海区到外海海区呈递增的趋势,外海海区的盐度显著高于近岸海区和养殖区,表明该海域受到傅疃河和巨峰河入海径流的影响较大。该海域中DIN、PO43–-P及SiO32–-Si浓度的空间水平分布特征也表现为自近岸海区到外海海区逐渐降低的趋势,结合盐度的空间变化趋势可推断,陆地入海径流对该海域的营养盐含量有较大的贡献。河流是海洋中溶解Si和P的主要来源,分别占海洋总输入量的82% (Tréguer, 1995)和75%~94% (Benitez- Nelson, 2000)。1月盐度的显著降低和SiO32–-Si含量的增高,表明该月有较强的陆地径流输入。该海域在2月的表层水温是全年最低的,此时,表层海水的密度较大,上下水层易发生垂直混合,垂直上升流可把海底的营养盐带到表层。由此可推测,2月表层海水中PO43–-P浓度的增高可能是陆地径流和垂直上升流双重原因导致的。
导致赤潮发生的真正原因目前尚未定论,但富营养化往往是赤潮发生的先决条件,富营养化水体中的N、P、Si营养盐为赤潮生物藻类的大量繁殖提供了物质基础(黄小平等, 2002)。构成赤潮的浮游生物种类很多,但甲藻和硅藻门的藻类大多是优势种,甲藻门约占42%,硅藻门约占37%(刘丽娟, 2005)。2017年的1、3月,调查海域的浮游植物物种多样性较低,此时,各个海区均出现了大量的骨条藻,优势度高达0.5~0.8,由此推测,该海域发生骨条藻赤潮的几率较大,应注意加强骨条藻赤潮的防范。赵冉等(2014)研究表明,骨条藻可作为冲淡水流向指示物种。因此,本研究中傅疃河和巨峰河入海径流可能是造成骨条藻丰度及优势度较高的原因之一。安达六郎(1973)研究表明,发生赤潮时,浮游植物丰度一般为(3~100)×105cells/L,而调查海域浮游植物丰度为(0.049~3.031)× 104cells/L,并未达到赤潮级别。调查结果显示,浮游植物丰度与温度呈极显著的负相关关系,表明该海域的浮游植物群落结构季节演替性较明显。
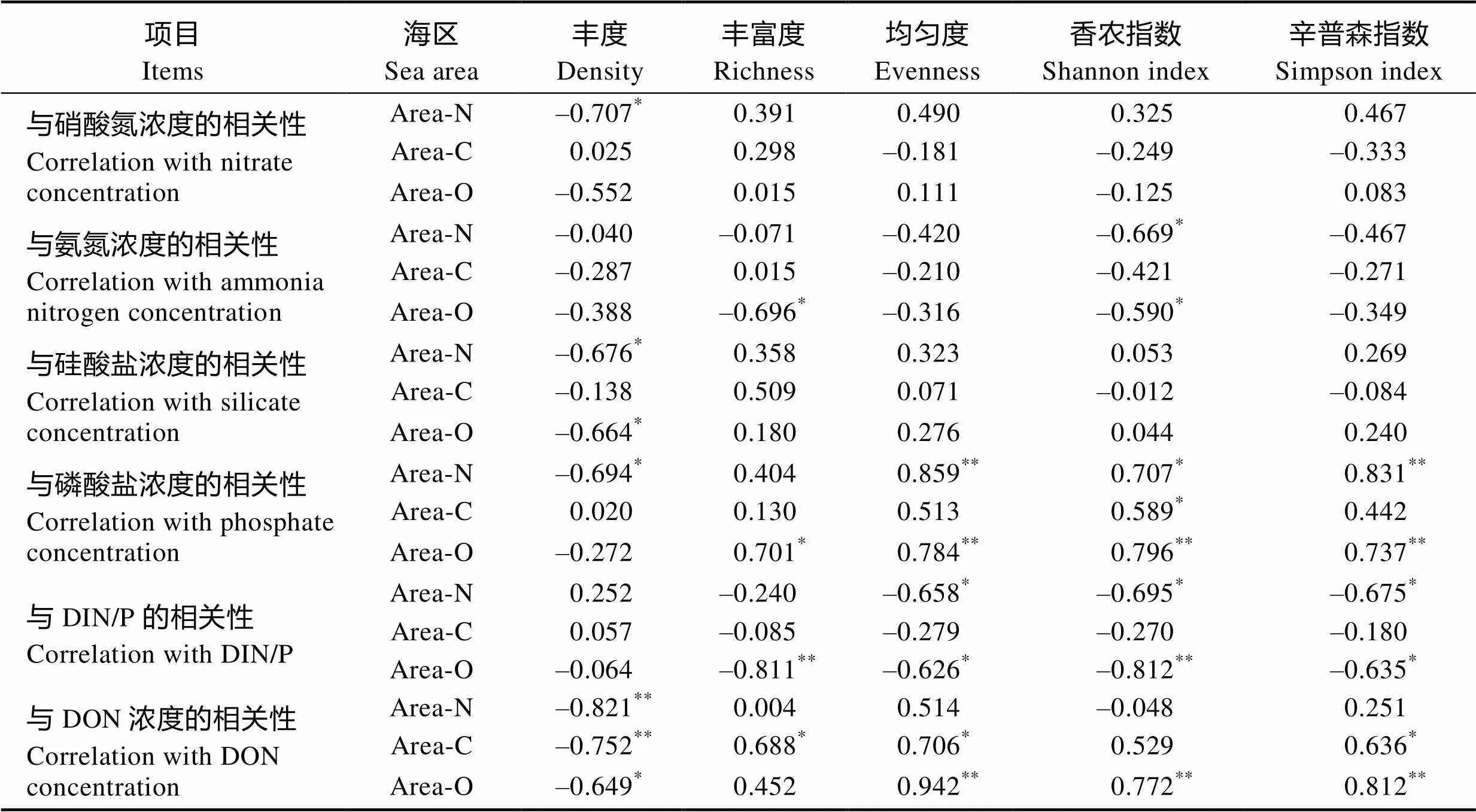
表7 不同海区浮游植物丰度和多样性指数与营养盐浓度的相关性
注:*为显著相关(<0.05);**为极显著相关(<0.01)
Note: * indicatedsignificant differences (<0.05), and ** indicated highly significant differences
DON是可被浮游植物直接或间接利用的重要营养源,它可与DIN之间进行相互转化,是构成近海富营养化的重要氮源物质,也是生物碎屑分解的中间产物,在赤潮的持续发展中起着关键作用(徐环等, 2016)。调查海域的浮游植物丰度与DON含量呈显著负相关关系,表明浮游植物大量繁殖可能对DON有较大的消耗。11月采集的样品中,浮游植物丰度极低,该月采样前7 d温度骤降(可通过历史天气查询得知),可能导致了浮游植物的大量衰亡,衰亡的浮游植物残骸可分解产生DON,从而导致11月的DON含量较高。近岸海区浮游植物丰度与多种溶解无机营养盐呈显著的负相关,外海海区浮游植物丰度与SiO32–-Si也呈显著负相关,表明浮游植物的大量繁殖对溶解无机营养盐也有较大的消耗,且对于外海海区,浮游植物对SiO32–-Si的消耗较明显。与此不同的是,紫菜养殖区浮游植物丰度虽与大部分溶解无机营养盐含量均呈负相关,却均没有达到显著水平,这可能是由于紫菜与浮游植物之间对溶解无机营养盐的竞争性吸收,从而减弱了浮游植物丰度与溶解无机营养盐的负相关性的程度。
浮游植物多样性指数的月份比较结果显示,该海域1、3月的浮游植物群落结构稳定性及物种多样性较低。钱宏林等(2000)研究表明,盐度大幅度变化对浮游植物群落结构稳定性影响较大。受到入海径流的影响,1月盐度显著低于其他月,由此可推测,盐度的大幅降低、丰富的营养盐及相对稳定的温度可导致骨条藻等硅藻的暴发性生长,造成了1月该海域浮游植物丰度升高,而群落结构稳定性及物种多样性降低。2月浮游植物的部分优势藻种发生演替衰亡,该月衰亡的浮游植物残骸分解形成的NH4+-N,可能是导致3月NH4+-N浓度升高的原因之一。研究表明,骨条藻在P胁迫情况下,可利用自身较高的吸收速率和较小的细胞体积有效摄取环境中的无机磷快速生长并取得优势,而角毛藻可利用细胞内源磷库以补偿外源磷的降低对细胞生长的影响,从而维持较高的细胞活性(王丹等, 2008)。由此可推测,3月时,NH4+-N浓度升高且PO43–-P浓度的下降可能导致了骨条藻、角毛藻等硅藻成为具有较高优势度的优势种,从而造成该月浮游植物群落结构稳定性及物种多样性的降低。
研究表明,海水中浮游植物的生长繁殖受N、P、Si等营养盐的限制,Redfield等(1963)首先提出化学计量的营养盐限制标准,即nDIN∶nSi∶nP=16∶16∶1 (Redfield比值),这主要是针对硅藻最适的营养需求。李京(2008)研究表明,DIN/P值随硅藻的生长而升高,随着甲藻的生长而下降,由此推测,调查海域以硅藻为主要优势种、甲藻优势种少的群落结构特征可能是适应该海域高DIN/P值(DIN/P=19.9~103.3)的自然选择结果。
NH4+-N浓度和DIN/P的值与3个调查海区的浮游植物均匀度、香农指数和辛普森指数的相关性研究表明,调查海域如果受到高浓度DIN(尤其是高浓度NH4+-N)的污染,且PO43–-P被过度消耗得不到有效补充,可能会引起浮游植物物种多样性和群落结构稳定性的降低,进而使赤潮发生概率提高。而紫菜养殖区的浮游植物多样性指数与NH4+-N浓度或DIN/P值的负相关程度均达不到显著水平,且负相关程度均低于近岸海区与外海区,这表明紫菜养殖对NH4+-N的消耗作用比较明显,且对该海区高DIN/P值的降低有一定的作用,有利于防止高NH4+-N或高DIN/P值引起的浮游植物群落结构的稳定性及物种多样性降低,从而降低赤潮发生概率。王洪斌等(2014)研究发现,江苏海州湾的条斑紫菜养殖海域的浮游藻类生物多样性指数高于非养殖海域;龙须菜()栽培有利于维持浮游植物的群落结构稳定(李宵, 2009)。这些研究结果与本研究结果相似,都表明大型海藻养殖活动的开展对浮游植物群落结构有较强的调控作用。
An DLL. Red tide organisms and red tide ecology. Fisheries and Civil, 1973, 9(1): 31-36 [安达六郎. 赤潮生物と赤潮生态. 水产土木, 1973, 9(1): 31-36]
An F, Tao JH. Method of two-grade fuzzy synthesis assessment for seawater eutrophication and its application in Bohai Bay. Marine Environmental Science, 2008, 27(4): 68-71 [安斐, 陶建华. 渤海湾海水富营养化的二级模糊综合评价方法及其应用.海洋环境科学, 2008, 27(4): 68-71]
Benitez-Nelson CR. The biogeochemical cycling of phosphorus in marine systems. Earth-Science Reviews, 2000, 51(1-4):109-135
Huang BQ, Hong HS, Wang HL. Application of microphytoplankton in environmental science research. Fujian Environment, 1995, 12(4): 18-19 [黄邦钦, 洪华生, 王海黎. 微型浮游植物在环境科学研究中的应用. 福建环境, 1995, 12(4): 18-19]
Huang XP, Huang LM, Tan YH,. Relationship between red tide and environmental conditions in coastal waters. Marine Environmental Science, 2002, 21(4): 63-69 [黄小平, 黄良民, 谭烨辉, 等. 近海赤潮发生与环境条件之间的关系. 海洋环境科学, 2002, 21(4): 63-69]
Justić D, Rabalais NN, Turner RE. Stoichiometric nutrient balance and origin of coastal eutrophication. Marine Pollution Bulletin, 1995, 30(1): 41-46
Li J. The research of nutrient structure and its function on the succession of phytoplankton predominant species in the high frequent harmful algae blooms occurrence areas in East China sea. Master's Thesis of Ocean University of China, 2008 [李京. 东海赤潮高发区营养盐结构及对浮游植物优势种演替的作用研究. 中国海洋大学硕士研究生学位论文, 2008]
Li QX, Tao JH. Application of phytoplankton community indexes in coastal eutrophication assessment. China Environmental Science, 1999, 19(6): 548-551 [李清雪, 陶建华. 应用浮游植物群落结构指数评价海域富营养化. 中国环境科学, 1999, 19(6): 548-551]
Li X. Effects of mariculture on the phytoplankton community and water quality. Master's Thesis of Jinan University, 2009 [李宵. 海水养殖对浮游植物群落结构和水质的影响. 暨南大学硕士研究生学位论文, 2009]
Liu JT, Wang M, Liu SJ,. Pollutant source apportionment of shallow groundwater in Taoluo area, Rizhao City. Yangtze River, 2017, 48(9): 12-15 [刘久潭, 王敏, 刘书江, 等. 日照市涛雒地区浅层地下水污染源解析. 人民长江, 2017, 48(9): 12-15]
Liu LJ. HABs' actuality and prevention. Shandong Fisheries, 2005(9): 19-21 [刘丽娟. 赤潮现状及防治对策. 齐鲁渔业, 2005(9): 19-21]
Liu XY, Yan JH. Investigation on water quality of Futuan River in Rizhao, China. Environment and Sustainable Development, 2017, 42(5): 172-173 [刘昕宇, 颜景浩. 关于日照傅疃河水质调查分析. 环境与可持续发展, 2017, 42(5): 172-173]
Qian HL, Liang S, Qi YZ. Study of the characteristics and the causes of formation on the red tides in coastal Guangdong sea. Ecologic Science, 2000, 19(3): 8-16 [钱宏林, 梁松, 齐雨藻. 广东沿海赤潮的特点及成因研究. 生态科学, 2000, 19(3): 8-16]
Redfield AC, Ketchum BH, Richards FA. The influence of organisms on the composition of seawater. Sea, 1963, 40(6): 640-644
Shen YF, Gu MR, Gong XJ,. New technology of microbiological monitoring. Beijing: China Architecture and Building Press, 1990 [沈韫芬, 顾曼如, 龚循矩, 等. 微型生物监测新技术. 北京:中国建筑工业出版社, 1990]
Shi Q. Research progress on nutrient limitation of phytoplankton. Science and Technology Information, 2012(21): 114-115 [史乾. 浮游植物营养盐限制的研究进展. 科技资讯, 2012 (21): 114-115]
Sun XM, Xia B, Guo F,. Structure characteristics of phytoplankton community and the relationship with environmental factors in offshore area of Laoshan, Qingdao. Progress in Fishery Sciences, 2013, 34(3): 46-53 [孙雪梅, 夏斌, 过锋, 等. 青岛崂山近岸海域浮游植物群落结构及其与环境因子的关系. 渔业科学进展, 2013, 34(3): 46-53]
Sun XM, Xu DH, Xia B,. Species composition and seasonal variation of netz-phytoplankton in the central Bohai Sea. Progress in Fishery Sciences, 2016, 37(4): 19-27 [孙雪梅, 徐东会, 夏斌, 等. 渤海中部网采浮游植物种类组成和季节变化. 渔业科学进展, 2016, 37(4): 19-27]
Tréguer P, Nelson DM, Van Bennekom AJ,. The silica balance in the world ocean: A reestimate. Science, 1995, 268(5209): 375-379
Wang D, Huang CX, Huang BQ,. Physiological responses of two typical species of diatoms to phosphorus stress in Yellow Sea. Marine Sciences, 2008, 32(5): 22-27 [王丹, 黄春秀, 黄邦钦, 等. 黄海两种典型硅藻的磷胁迫生理研究. 海洋科学, 2008, 32(5): 22-27]
Wang HB, Li XS, Song XM,. Effect offarming on the structure and genetic diversity of phytoplankton community in Haizhou Bay. Marine Sciences, 2014, 38(12): 103-109 [王洪斌, 李信书, 宋秀梅, 等. 海州湾条斑紫菜养殖对浮游藻类群落结构及遗传多样性的影响. 海洋科学, 2014, 38(12): 103-109]
Wang YH, Lȕ ZB, Fang Y,. Relationship between structure of nutrient and diversity index of nanophytoplankton in Weihai coastal seawater. Progress in Fishery Sciences, 2014, 35(2): 1-7 [王育红, 吕振波, 房燕, 等. 威海海域微型浮游植物多样性指数与营养盐结构的关系. 渔业科学进展, 2014, 35(2): 1-7]
Xu H, Zhang GC, Li YY,. Progress in bioavailability of dissolved organic nitrogen to phytoplankton in the coastal ecosystems. Transactions of Oceanology and Limnology, 2016(4): 59-67 [徐环, 张桂成, 李悦悦, 等. 近海溶解有机氮对浮游植物生物可利用性研究进展. 海洋湖沼通报, 2016(4): 59-67]
Wu YH, Wang LB. Water environment evaluation in Futuan watershed and industrial structure optimization.Territory and Natural Resources Study, 2004(4): 76-77 [吴艳红, 王立本. 傅疃河流域水环境现状评价及工业结构优化对策. 国土与自然资源研究, 2004(4): 76-77]
Zhao R, Sun ZZ, Zhang YP,. The netz-phytoplankton assemblage structure of Qingcaosha Reservoir adjacent water. Progress in Fishery Sciences, 2014, 35(3): 9-18 [赵冉, 孙振中, 张玉平, 等. 青草沙邻近水域网采浮游植物群落结构特征. 渔业科学进展, 2014, 35(3): 9-18]
The Temporal and Spatial Distribution Characteristics of the Nutrients and Community Structures of Phytoplankton and Their Relationships in theCulture Areas of Rizhao, China
LIANG Zhourui1,2, SUN Tengqin3, WANG Wenjun1,2①, LIU Fuli1,2, CAO Yuan3, LIANG Qidi1, SUN Xin1, SUN Xiutao1,2, WANG Feijiu1
(1. Yellow Sea Fisheries Research Institute, Chinese Academy of Fishery Sciences, Key Laboratory of Sustainable Development of Marine Fisheries, Ministry of Agriculture and Rural Affairs, Qingdao 266071; 2. Laboratory for Marine Fisheries Science and Food Production Processes, Polit National Laboratory for Marine Science and Technology (Qingdao), Qingdao 266071; 3. College of Fisheries and Life Science, Shanghai Ocean University, Shanghai 201306)
The culture area ofsp.at Fuxin fishing port, in Rizhao City, China, were investigated from November 2016 to March 2017. The temporal and spatial distribution characteristics of the nutrient and community structures of phytoplankton and their relationships were analyzed. The main conclusions were as follows: Generally, the spatial horizontal distribution of inorganic nitrogen, phosphate and silicate concentrations showed a decreasing trend between the nearshore area and offshore sea areas. Nitrate, silicate, phosphate, dissolved inorganic nitrogen (DIN) and dissolved organic nitrogen concentrations were significantly different in the different months. Over, all the surveyed sea area was at a mesotrophic level. The range of phytoplankton density was (0.049~3.031) × 104cells/L and the phytoplankton growth was likely to be controlled by the silicate and phosphate. 37 species of diatom belonging to 27 genera, followed by dinoflagellates represented by 8 taxa (7 genera) and chrysophyceae by 1 taxa (1 genera) were recorded. The most common and dominant species weresp.,sp., andsp. The diversity index of phytoplankton decreased significantly in January and March, which may be caused by the sharp decrease in salinity and richness of nutrients in January, significant rises in ammonia nitrogen concentrations, and significant decrease in phosphate concentration in March. Theammonia nitrogen concentrationand DIN/P ratio were both negatively correlated with the diversity index of phytoplankton. The negative relationship was significant for the non-cultured area, while it was not significant for the cultured area. It can be inferred from the above correlation analysis results thatsp.cultivation is beneficial for reducing the ammonia nitrogen levels and DIN/P ratio in the sea area, thus increasing the stability of phytoplankton community structures and the species diversity of phytoplankton, which will help to prevent the occurrence of red tides. These results provide information valuable for marine environment protection, red tide prevention and future developments of aquaculture systems.
cultivation; Phytoplankton; Nutrients; Distribution
WANG Wenjun, E-mail: wjwang@ysfri.ac.cn
* 山东省农业良种工程(南种北繁)项目(2017LZN013)、山东省重点研发计划项目(2016GSF115038)、中国水产科学研究院基本科研业务费专项资金(2015A02)和现代农业产业技术体系(CARS-50)共同资助 [This work was supported by the Shandong Agricultural Good Seed (South to North) Project (2017LZN013), Key Research and Development Plan of Shandong Province (2016GSF115038), Special Scientific Research Funds for Central Non-Profit Institutes, Chinese Academy of Fishery Sciences (2015A02), and China Agriculture Research System(CARS-50)]. 梁洲瑞,E-mail: liangzr@ysfri.ac.cn
汪文俊,研究员,E-mail: wjwang@ysfri.ac.cn
2018-07-02,
2018-08-28
Q178.1
A
2095-9869(2019)05-0078-11
10.19663/j.issn2095-9869.20180702001
http://www.yykxjz.cn/
梁洲瑞, 孙藤芹, 汪文俊, 刘福利, 曹原, 梁启迪, 孙昕, 孙修涛, 王飞久. 日照紫菜养殖海域营养盐的时空分布特征及其与浮游植物群落结构的相关性分析. 渔业科学进展, 2019, 40(5): 78–88
Liang ZR, Sun TQ, Wang WJ, Liu FL, Cao Y, Liang QD, Sun X, Sun XT, Wang FJ. The temporal and spatial distribution characteristics of the nutrient and community structures of phytoplankton and their relationships in theculture areas in Rizhao, China. Progress in Fishery Sciences, 2019, 40(5): 78–88
(编辑 陈严)
猜你喜欢
环境工程技术学报(2022年3期)2022-06-05
海洋与渔业(2022年2期)2022-05-18
东方教育(2017年19期)2017-12-05
江苏农业科学(2017年12期)2017-09-16
绿色科技(2016年24期)2017-03-30
河北渔业(2017年1期)2017-02-21
湖北农业科学(2014年1期)2014-03-22
棋艺(2001年9期)2001-07-17
棋艺(2001年11期)2001-05-21
棋艺(2001年11期)2001-05-21
