
水分胁迫对双荚决明能量利用及生长的影响
2019-09-17 07:32廖飞勇文瑛黄琛斐
江苏农业科学 2019年7期
廖飞勇 文瑛 黄琛斐
摘要:为确定土壤含水量对双荚决明的影响以便在实际中指导养护,研究水分胁迫对双荚决明二年生苗木能量利用及生长的影响。结果表明,水淹双荚决明31 d后水淹茎基部长出许多不定根且漂浮在水面,其对能量的转换、传递和固定稍有下降,对能量的消耗增加,能适应90 d以上的水淹。轻度干旱(土壤含水量为15%~25%)对植物的生长和总的能量利用效率影响不大,指导园林实践可以降低养护成本。中度干旱(土壤含水量为10%~15%)19 d后明显抑制双荚决明的生长,对能量的转换、传递和固定稍下降,对能量的消耗增加。重度干旱(土壤含水量<10%)处理 13 d 后,双荚决明生长受到严重威胁,干旱处理23 d后死亡。养护过程中轻度干旱不影响双荚决明的生长,中度干旱时间不能超过19 d,重度干旱时间不能超过13 d。
关键词:水分胁迫;双荚决明;能量利用;生理影响
中图分类号: Q945.78 文献标志码: A 文章编号:1002-1302(2019)07-0156-05
水是植物生长必需的环境因子,过多过少都会影响植物的生长甚至导致植物死亡。双荚决明(Cassia bicapsularis)是苏木科决明属植物,10月左右开金黄色的花,观赏性强且时间长,在园林中各类型绿地中广泛应用并受到市民的喜爱。对双荚决明的部分研究主要集中在生物学特性[1]、花粉结构[2]、主要成分[3]、抗寒性[4]、抗高温[5]、组织培养[6-7]、生理生态调控[8]及引种驯化[9]等方面,水分对其生理的影响已经有初步的研究[10],但水分胁迫对其能量利用的影响还未见报道。为合理利用双荚决明,本研究分析了不同土壤水分含量对双荚决明能量利用的影响,以探明其适应机制,进而指导双荚决明在园林中的应用。
1 材料与方法
1.1 试验材料
选用二年生双荚决明实生苗为试验材料,种植于湖南省长沙市中南林业科技大学长沙校园内。选用40株上盆(盆直径220 cm,高320 cm),土壤为混合土(黏土、腐殖土体积比为 1 ∶ 1)。生长30 d后选择生长一致的25株植株移入室内,随机分为5组,每组5盆。利用5月份的自然温度和湿度进行培养,用人工碘钨灯进行光照,光照度为600 μmol/(m2·s)。
1.2 土壤含水量处理
试验分为5组,分别是对照(CK)(每天浇透土壤,其含水量在30%以上);水淹处理(WS)(将种植盆浸入水中,水淹茎基部5 cm,土壤的含水量大于70%);轻度干旱处理(SD)(土壤含水量的日变化在15%~25%之间);中度干旱处理(MD)(土壤含水量在10%~15%之间);重度干旱处理(HD)(试验开始后,一次性浇透水后持续不浇水直到植物死亡)。其中轻度干旱处理和中度干旱处理在正式进入试验前在室内就设置好水分梯度,在此期间其余3个处理均为正常水分管理。
1.3 测定内容和方法
土壤含水量使用高精度土壤水分测试仪TDR100(美国Spectrum公司生产)进行测定;叶绿素浓度用分光光度法测定[11];叶绿素荧光参数、光曲线、A-Ci曲线用Licor 6400便携式光合仪进行测定;叶生物量用干质量法测定;叶片中可溶性糖含量用蒽酮比色法测定;丙二醛(MDA)含量的测定参考杨柳青等的研究[12]。
2 结果与分析
2.1 不同土壤含水量对叶绿素含量的影响
叶绿素含量影响植物对光能的吸收和转化,由表1可知,处理12 d后,除SD处理外其他各处理叶绿素含量与对照相比均有不同程度的下降,其中MD组下降幅度最大,下降19.56%。处理20 d后,重度干旱植株的总叶绿素含量略有增加,但与对照差异不显著;中度干旱处理的植株叶绿素含量下降最多。在3次测定中,水淹处理的总叶绿素含量随水淹时间延长逐渐降低,水淹31 d后只有对照的74.79%。中度干旱处理植株的总叶绿素含量均较其他处理低,对照和轻度干旱处理的总叶绿素含量较高。
2.2 不同土壤含水量对叶绿素荧光参数的影响
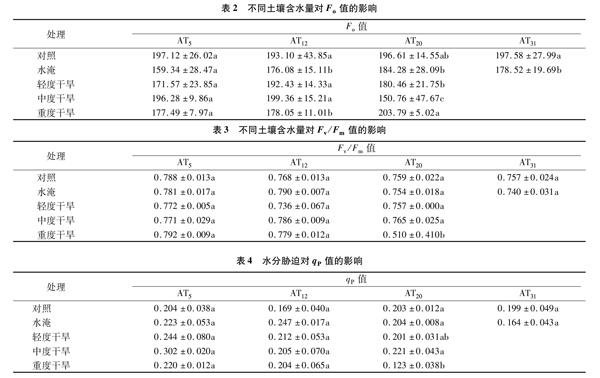
2.2.1 不同土壤含水量对最小初始荧光(Fo)值的影响 Fo是经过充分暗适应的光系统Ⅱ(PSⅡ)反应中心全部开放时的叶绿素荧光发射强度。表2结果表明,处理12 d后,各处理之间Fo值差异不大,其中水淹处理的值最低,与对照相比下降了8.81%。处理20 d后,中度干旱处理的Fo值是对照组的76.68%,而重度干旱处理的Fo值是对照组的1.04倍,水淹处理的Fo值低于对照,但差异未达显著水平。
2.2.2 不同土壤含水量对光化学量子效率(Fv/Fm)的影响 Fv/Fm值指没有环境胁迫并经过充分暗适应的叶片,其PSⅡ最大的(潜在)光化学量子效率,其值在0.75~0.85之间。表3结果表明,处理5 d后各处理的Fv/Fm值有变化,但都在正常范围内。处理12 d后,各处理Fv/Fm值差异较小,只有轻度干旱的Fv/Fm值下降。处理20 d后,重度干旱处理的Fv/Fm值是对照的67.19%,说明此时重度干旱处理的植株已受到严重胁迫,其余各组差异较小。
2.2.3 不同土壤含水量对光化学淬灭系数(qP)的影响 qP反映了PSⅡ反应中心的开放程度,1-qP则反映了反应中心的关闭程度,反映了质体醌(QA)的还原程度。表4结果表明,处理5 d后,水淹、轻度干旱、中度干旱、重度干旱4个处理的qP值均升高,分別是对照的1.09、1.20、1.48和1.08倍。处理12 d后,4个处理的qP值分别是对照的1.46、1.25、1.21和1.21倍。处理20 d后,重度干旱处理的qP值降至对照的60.59%,中度干旱处理的qP值是对照的1.09倍,其余2个处理的值与对照相近。综合看出,中度干旱处理的qP值均高于对照;轻度干旱胁迫使qP值上升,起初随着时间的延长,其值逐渐接近对照,时间过长,超过20 d后,其值低于对照;水淹处理的qP值随着时间的延长先升后降,说明短暂的水淹提高了植物对光能的利用效率,长时间处理导致其对光能的利用率下降。
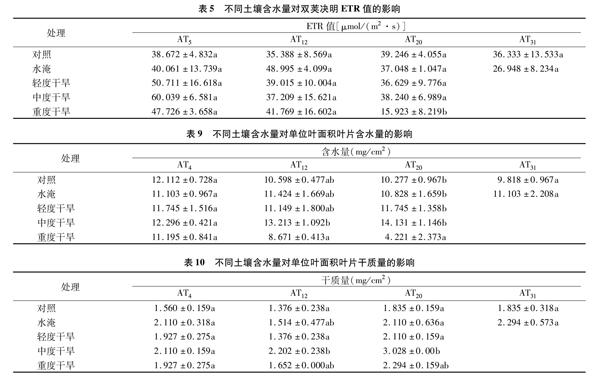
2.2.4 不同土壤含水量对表观电子传递速率(ETR)的影响 表5结果表明,处理5 d后,水淹、轻度干旱、中度干旱、重度干旱4个处理的ETR值均升高,分别是对照的1.04、1.31、1.55、1.23倍,但差异未达显著水平。处理12 d后,4个处理的ETR值分别是对照的1.38、1.10、1.05、1.18倍。处理20 d后,重度干旱处理的ETR值是对照的40.57%,其余处理的值与对照组相近。综合看出,水淹处理的ETR值随着时间的延长先升后降,说明短时间水淹促进光合电子的传递;长时间的重度干旱胁迫,抑制了表观光合电子的传递,从而抑制了光合能量的积累。
2.3 不同土壤含水量对最大羧化效率的影响
表6表明,水淹处理7、13、19和31 d后最大羧化效率(Vcmax)分别是对照的104.80%、70.43%、60.74%、49.08%,表明随水淹时间的延长,羧化效率逐渐降低。轻度干旱处理后Vcmax值升高,说明轻度干旱提高了双荚决明的羧化效率。中度干旱处理7、13 d后,Vcmax值分别是对照的1.03、1.30倍,说明短时间中度干旱使羧化效率提高;处理19 d后,Vcmax值下降,是对照的65.32%,说明长时间的中度干旱,使双荚决明的羧化效率降低。重度干旱处理7 d使Vcmax值下降,但是处理13 d后使Vcmax值上升。
2.4 不同土壤含水量对净光合速率的影响
表7结果表明,处理12 d后,水淹和輕度干旱处理的净光合速率分别是对照的 1.08 和1.03倍;中度干旱和重度干旱的净光合速率都低于对照,分别是对照的79.10%和95.96%。处理20 d后,部分处理间的差异显著,水淹处理、中度干旱处理的净光合速率分别是对照的69.26%、47.65%;重度干旱处理的净光合速率是负值,此时植物几乎没有进行光合作用,叶片接近死亡。处理31 d后,水淹处理的净光合速率只有对照的40.93%,但此时水淹组植株已长出许多不定根,且不定根漂浮在水面上。
2.5 不同土壤含水量对呼吸速率的影响
表8表明,水淹处理19 d内呼吸速率下降,31 d时呼吸速率上升,是对照的143.5%。轻度干旱处理13 d使呼吸速率下降,处理19 d后呼吸速率大于对照。中度干旱处理植株的暗呼吸速率较高,处理7、13、19 d分别是对照的 1.13、1.37、1.63倍。说明植物通过呼吸代谢来增强对环境的抵抗能力。
2.6 不同土壤含水量对叶片含水量的影响
由表9可见,处理后4 d,各处理间差异不显著。处理后 12 d,重度干旱组叶片含水量明显下降,是对照组的81.82%,与对照的差异未达到显著水平;中度干旱组双荚决明叶片含水量是对照组的1.25倍。处理20 d后,重度干旱组的单位叶面积含水量继续下降,是对照的41.07%,差异达显著水平;中度干旱组的单位叶面积含水量是对照组的1.38倍。
2.7 不同土壤含水量对叶片生物量的影响
由表10可以看出,与对照相比,各处理的单位叶面积生物量都增加,但是不同处理、不同时间的增加程度不同。处理20 d时,中度干旱处理的植株干质量增加的最多,其次是重度干旱处理。
3 结论与讨论
3.1 水淹胁迫对能量利用的影响
3.1.1 水淹胁迫对能量吸收的影响 光能可被叶绿素吸收,叶绿素含量的高低直接影响能量的吸收。本研究中,水淹处理后随着时间的延长,总叶绿素含量逐渐下降,表明通过叶绿素捕获的光能逐渐下降。
3.1.2 水淹胁迫对能量转换的影响 本研究Fo值的变化表明,水淹胁迫后使光能中以荧光形式耗散的能量下降,刚开始时下降较多,随着时间的延长,其下降程度略有上升。Fv/Fm值的变化表明,水淹处理31 d后植株才受到胁迫。qP值的变化表明,水淹处理20 d内用于光化学反应的能量增加,但处理 31 d 后又下降。ETR值的变化表明,处理12 d内ETR值增加,但是随着处理时间的延长,其值逐渐下降,时间越长,其下降程度也越大。
3.1.3 水淹胁迫对能量固定和消耗的影响 本研究中处理7 d后,水淹植株的最大羧化效率增加了5%,但是随着水淹时间的延长,最大羧化效率下降,处理31 d后,处理植株的值只有对照的49.08%,表明植株固定CO2形成碳水化合物的能力下降。本研究还表明,净光合速率的变化与最大羧化效率的变化相一致。水淹时间越长,其净光合速率也越低。结果显示,水淹处理19 d内,其呼吸速率下降,表明其能量的消耗下降;处理31 d后,其呼吸速率是对照的143.5%,表明此时其能量的消耗增大,以适应根系被淹的环境。
3.1.4 水淹胁迫对植物形态的影响 水淹处理10 d后,地上茎基部开始生长出不定根,随着水淹时间的延长,不定根的数量不断增加,水淹31 d时,其不定根的数量增加较多,但是不定根都没有露出水面;60 d后植株生长良好,茎生不定根有许多浮在水面上,90 d后植株依然生长良好。
以上分析表明,水淹5 d左右促进双荚决明的生长,表现在qP值、ETR值、净光合速率、Vcmax值升高,Fo值和呼吸速率下降,这些变化是由于双荚决明提高了其对于光能的捕获、电子的传递和能量的固定,同时又降低了其能量的消耗,最终生物量增加。水淹处理31 d时双荚决明叶绿素含量、Fo值、Fv/Fm值、qP值、ETR值、Vcmax值、净光合速率均下降,但是呼吸速率、叶片含水量和生物量上升,表明双荚决明随着水淹时间的延长,吸收、传递的能量下降,但是其固定的能量还较多,说明植株能适应水淹环境,但是长期水淹影响其生长。
3.2 轻度干旱胁迫对能量利用的影响
3.2.1 轻度干旱胁迫对能量吸收的影响 本研究中,轻度干旱处理后随着时间的延长,总叶绿素含量下降,处理20 d时,只有对照的92.14%,表明通过叶绿素捕获的光能可能下降。
3.2.2 轻度干旱胁迫对能量转换的影响 本研究结果表明,轻度干旱胁迫使得Fo值下降,处理5 d时下降的较多,处理12 d时下降程度略有缓解,处理20 d时下降程度又有所提高。Fo值的变化表明,轻度干旱胁迫使得光能中以荧光形式耗散的能量减少。本研究中Fv/Fm值的变化表明,轻度干旱处理12 d后植株受到胁迫,20 d后又恢复正常。qP值的变化表明,轻度干旱处理12 d内,用于光化学反应的能量略有增加,但在处理20 d时略有下降。ETR值的变化表明,处理 12 d 时ETR值增加,是对照的110.25%,但是随着处理时间的延长,ETR值逐渐下降,处理20 d时,ETR值只有对照93.33%。
3.2.3 轻度干旱胁迫对能量固定和消耗的影响 本研究结果表明,处理7 d时轻度干旱植株的最大羧化效率增加了9.69%;处理13 d时最大羧化效率继续增加,增加了46.56%;处理19 d后植株的最大羧化效率略有下降,只有对照的97.91%。本研究结果表明,净光合速率在处理12 d时略有增加,处理20 d时则只有对照的85.10%。研究结果表明,水淹处理13 d内其呼吸速率下降,表明其能量的消耗下降;处理19 d后其呼吸速率是对照的132.84%,表明此时其能量的消耗增大,以加强代谢。
3.2.4 轻度干旱胁迫对植物形态的影响 轻度干旱处理 20 d 后,植物生长良好。数据分析表明,轻度干旱处理5 d植株并没有受到胁迫,且促进双荚决明的生长,表现在qP值、ETR值、Vcmax值、生物量增加,Fo值、Fv/Fm值下降,這些变化是由于双荚决明提高了其对于光能的捕获、电子的传递和能量的固定,同时又降低了其能量的消耗,最终生物量增加。轻度干旱处理20 d双荚决明叶绿素含量、Fo值、Fv/Fm值、qP值、ETR值、Vcmax值、净光合速率均下降,但是呼吸速率、叶片含水量和生物量均上升,植物吸收、传递和固定的能量下降,但是其固定的能量还是较多,因而说明双荚决明能适应轻度干旱环境。轻度干旱对双荚决明的生长影响较小,其生物量、色素和生长与对照组相比均无明显差异,同时可以节约灌溉用水量,降低养护成本。因而在园林养护中轻度干旱(保持土壤含水量为15%~25%)不仅能保证双荚决明的生长,而且可以节约灌溉成本。
3.3 中度干旱胁迫处理对能量利用的影响
3.3.1 中度干旱胁迫对能量吸收的影响 研究结果表明,中度干旱处理后引起总叶绿素含量下降,处理20 d时,只有对照的75.24%,表明通过叶绿素捕获的光能可能下降。
3.3.2 中度干旱胁迫对能量转换的影响 研究结果表明,中度干旱胁迫使Fo值发生变化,处理5 d时下降,处理12 d时有所提高,处理20 d时下降程度加大,Fo值的变化表明中度干旱胁迫后使光能中以荧光形式耗散的能量下降。本研究中Fv/Fm值的变化表明,中度干旱没有使植株受到胁迫。qP值的变化表明,中度干旱处理使用于光化学反应的能量增加。ETR值的变化表明,处理5 d内ETR值增加,是对照的155.25%,但是随着处理时间的延长,其值逐渐下降,处理 20 d 时,其值只有对照97.44%。
3.3.3 中度干旱胁迫对能量固定和消耗的影响 本研究表明,处理7 d中度干旱处理植株的最大羧化效率增加了3.50%;处理13 d最大羧化效率继续增加,增加了30.04%;处理19 d后,处理植株的值略有下降,只有对照的65.32%。本研究表明,净光合速率处理12 d时只有对照的79.10%,处理20 d时则只有对照的47.65%。研究结果表明,中度干旱胁迫后呼吸速率开始上升,处理7 d时是对照的112.71%,处理19 d后其呼吸速率是对照的162.59%,表明此时其能量的消耗增大,以加强代谢应对水分胁迫。
3.3.4 中度干旱胁迫对植物形态的影响 处理20 d植物生长良好。20 d后停止浇水1 d,植物萎蔫。数据分析表明,中度干旱处理5 d左右植株影响不大,表现为叶绿素含量和Fo值略有下降,qP值和Vcmax值略有提高,ETR值提高,Fv/Fm值正常,这种变化是双荚决明提高了其对于光能的捕获、电子的传递和能量的消耗,但对能量的固定能力下降,短时间内叶片生物量增加。中度干旱处理20 d,双荚决明的生长受到抑制,表现为叶绿素含量、Fo值、ETR值、Vcmax值、净光合速率下降,qP值略有增加,但是呼吸速率增加,叶片含水量和生物量均上升,双荚决明光的吸收、传递和固定的能量下降,但是叶片生物量还是较多,因而说明双荚决明还能耐受中等干旱的胁迫,植株外形也证明了这点。因而在园林养护中轻度干旱(保持土壤含水量在15%~25%间)20 d是双荚决明可以忍受的范围。
3.4 重度干旱胁迫处理对能量利用的影响
3.4.1 重度干旱胁迫对能量吸收的影响 研究结果表明,重度干旱处理后12 d导致总叶绿素含量略有下降,处理20 d时叶绿素含量增加,是对照的101.11%,其原因可能是叶片中含水量下降引起的。
3.4.2 重度干旱胁迫对能量转换的影响 研究结果表明,重度干旱胁迫使Fo值下降,处理5 d时下降较多,处理12 d时下降程度略有缓解;处理20 d时其值增加,Fo值的变化表明重度干旱胁迫20 d后使光能中以荧光形式耗散的能量增加。本研究中Fv/Fm值的变化表明,重度干旱处理20 d后植株受到胁迫,其值低于0.75。qP值的变化表明,重度干旱处理5 d后qP值增加;处理 20 d 后qP值下降,表明用于光化学反应的能量下降。ETR值的变化表明,处理12 d内ETR值增加;处理20 d后其值只有对照的40.57%。
3.4.3 重度干旱胁迫对能量固定和消耗的影响 本研究结果表明,处理7 d,重度干旱处理植株的最大羧化效率下降,但是处理13 d最大羧化效率却增加。本研究结果表明,净光合速率处理12 d时只有对照的95.96%,处理20 d时植株没有光合能力,表现为其净光合速率为负值。本研究结果表明,重度干旱胁迫后呼吸速率开始下降,处理7 d只有对照的84.75%,处理13 d后是对照的78.22%,表明此时其能量的消耗增大,以加强代谢应对水分胁迫。
3.4.4 重度干旱胁迫对植物形态的影响 处理17 d后植物开始萎蔫,但无黄叶,此时测得土壤含水量为7.4%;处理 20 d 后整株萎蔫,大部分叶片脱水,变干,几乎不出现黄叶,此时土壤含水量为6.2%。本研究结果还表明,重度干旱处理5 d后对植株略有影响,表现为Fo值、Vcmax值和叶片含水量下降,Fv/Fm略有增加但在正常范围内,qP值、ETR值和单位面积叶片生物量增加。这些变化表明,干旱改变了双荚决明能量的转换、传递和固定,但是在短时间内可以促进叶片生物量的积累以适应水分供应的不足。重度干旱处理12 d已对植株产生较大影响,表现为净光合作用的下降。重度干旱处理20 d后,双荚决明的生长受到抑制,表现为叶绿素含量、Fo值略有上升,Fv/Fm值、ETR值下降,光合作用停止,但是呼吸速率和单位面积叶片的生物量增加,叶片含水量下降,说明双颊决明对光的吸收、传递和固定的能量降低,特别是呼吸作用的停止说明植株已接近死亡,植株的外形也证明了这点。因而在园林养护中连续重度干旱12 d后(土壤含水量低于7.4%)必须给植株进行灌溉,以維持植株的正常生长,20 d是双荚决明可以忍受的极限。
通过上述研究与分析,最终得出4点结论:(1)水淹双荚决明31 d后植株生长良好,从水淹茎基部长出许多不定根漂浮在水面;植株能忍受90 d以上的水淹胁迫,其对能量的转换、传递和固定稍下降,对能量的消耗增加,但是能适应水淹的环境。(2)轻度干旱(土壤含水量在15%~25%间)下植株有较高的净光合速率、ETR值、叶绿素总含量、Vcmax值,对植物总能量的利用效率影响不大,指导园林实践可以降低养护成本。(3)中度干旱(土壤含水量日变化在10%~15%间) 5 d 对植株影响不大,19 d后明显抑制双荚决明的生长,对能量的转换、传递和固定稍下降,对能量的消耗增加。(4)重度干旱处理13 d后,双荚决明生长受到严重威胁,干旱23 d后植株死亡。
因此,可以初步确定18%~25%的土壤含水量为双荚决明在园林绿地中的最佳含水量,但是植物对水分的适应较为复杂,不仅与光、温度和土壤含水量有关,同时也与个体生长状态有关。本研究仅仅分析了双荚决明对于能量的转换、传递、固定和消耗,对于其适应的酶活性及结构变化还需进一步探究。
参考文献:
[1]李荣林. 双荚决明的生物学特性与木本模式植物研究[D]. 南京:南京林业大学,2006.
[2]李荣林,王会宁. 双荚决明成熟花粉的超微结构观察[J]. 安徽农业科学,2011,39(26):16154-16155.
[3]梁 倩,刘蔚漪,徐文晖. 昆明引种的双荚决明花挥发油化学成分分析[J]. 西部林业科学,2012,41(4):108-109.
[4]卢 芳. 双荚槐和伞房决明抗寒性的测定[J]. 中南林业科技大学学报,2007,27(3):84-86.
[5]廖飞勇. 高温强光对双荚决明的影响及其园林应用[J]. 北方园艺,2010(7):96-99.
[6]韩 珊,石大兴,王米力,等. 双荚决明的组织培养与植株再生[J]. 植物生理学通讯,2006,42(2):245.
[7]刘正兰,冯秋平. 培养条件对双荚决明愈伤组织诱导的影响[J]. 亚热带植物科学,2011,40(2):44-46.
[8]杜宗现. 盆栽双荚决明生长发育的生理生态调控研究[D]. 郑州:河南农业大学,2010.
[9]邱记东,贺小秀. 双荚决明的引种与驯化研究[J]. 中国果菜,2010(9):29.
[10]蔡思琪,廖飞勇. 淹水胁迫对双荚决明生长及生理特性的影响[J]. 北方园艺,2017(3):71-75.
[11]蒋 慧,王秋姣,冉德龙,等. 不同水分胁迫对金焰绣线菊光合特性的影响[J]. 江苏农业科学,2015,43(5):178-180.
[12]杨柳青,曾 红,朱小青,等. 短蕊景天等4种植物的水分胁迫研究[J]. 湖南林业科技,2016,43(1):1-5.曹 萌,南冠君,高玉琼,等. 重金属对豌豆幼苗抗性生理指标的影响[J]. 江苏农业科学,2019,47(7):161-165.
