
NDV/FMW对耐维罗非尼的恶性黑色素瘤细胞免疫原性死亡相关分子表达的影响
2019-09-11 07:59邵晓雁王雪珂陈健华朱忠政孟松树
同济大学学报(医学版) 2019年4期
邵晓雁, 王雪珂, 陈健华, 朱忠政, 孟松树, 许 青
(1. 同济大学附属第十人民医院肿瘤科,上海 200072; 2. 上海市皮肤病医院肿瘤科,上海 200443; 3. 大连医科大学肿瘤干细胞研究院,大连 116044)
恶性黑色素瘤是一种高度侵袭性的肿瘤,预后差、严重威胁人类健康[1]。据报道,中国的晚期黑色素瘤患者约25.5%存在鼠类肉瘤病毒癌基因同源B1(v-Raf murine sarcoma viral oncogene homolog B1, BRAF)基因突变[2]。2011年8月,维罗非尼(vemurafenib)被批准用于BRAF突变的转移性晚期黑色素瘤患者[3]。维罗非尼可提高患者的无进展生存期(progression-free survival, PFS),但大多数患者6~7个月出现耐药[4]。维罗非尼耐药以后,治疗手段有限且疗效不佳,因此需要探讨维罗非尼耐药后恶性黑色素瘤新的治疗手段。
溶瘤病毒(oncolytic virus, OVs)在肿瘤治疗方面取得重要突破,但是OVs在恶性黑色素瘤vemurafenib耐药治疗中的价值尚不清楚。既往文献[5-6]显示,OVs除对肿瘤细胞产生直接的溶解作用外,还可诱导表达或释放钙网蛋白(calreticulin, CRT)、三磷酸腺苷(adenosine triphosphate, ATP)、高迁移率族蛋白1(high mobility group protein 1, HMGB1)、热休克蛋白90(heat shock protein 90, HSP90)等激起细胞免疫原性的损伤相关分子模式(damage associated molecular patterns, DAMPs),进而诱发非耐药肿瘤细胞免疫原性死亡(immunogenic cell death, ICD)[8-9]。
肿瘤的发生发展与免疫机制关联密切[9-10],本研究旨在细胞水平上验证新城疫病毒毒株FMW(Newcastle disease virus, NDV/FMW)对恶性黑色素瘤WM3248/vemurafenib细胞株的杀伤效应,进一步探讨NDV/FMW对耐药细胞杀伤效应的可能免疫机制,为恶性黑色素瘤维罗非尼耐药的临床治疗提供依据。
1 材料与方法
1.1 实验材料与仪器
人BRAF突变的黑色素瘤WM3248细胞株和WM3248/vemurafenib细胞株由Joon Kim实验室提供[11];鸡胚成纤维细胞株DF1购自美国ATCC公司;NDV/FMW由大连医科大学孟松树实验室提供;DMEM培养基、0.25%胰酶(Trypsin-EDTA)、RPMI-1640培养基、胎牛血清(FBS)、双抗(Pen Strep)购自美国Gibco公司;兔抗人cleaved Caspase-3抗体、兔抗人poly(ADP-ribose)polymerase(PARP)抗体、兔抗人GAPDH抗体购自美国Cell公司;鼠抗人HN抗体和鼠抗人HSP90抗体购自美国Santa Cruz公司;兔抗人CRT多克隆抗体和兔抗人HMGB1抗体购自英国Abcam公司;羊抗兔Alexa Fluor488抗体和羊抗鼠Alexa Fluor 568抗体购自美国Invitrogen公司;米托蒽醌(MTX)购自美国Selleckchem公司;DAPI购自美国Sigma公司;Cell Counting Kit-8(CCK-8)试剂盒购自美国Med Chem Express公司;ATP检测试剂盒购自上海碧云天生物技术有限公司;Pierce蛋白浓缩管购自美国Thermo Scientific公司;多功能酶标仪-PE(多模式微孔板检测系统)购自美国Perkin Elmer公司;激光(扫描)共聚焦显微镜购自德国Leica公司;凝胶成像(蛋白)仪购自美国Bio-Rad公司;流式细胞分析系统-C6购自美国BD公司。
1.2 细胞培养
人BRAF突变的黑色素瘤WM3248细胞株和WM3248/vemurafenib细胞株用含5%胎牛血清、1%双抗的RPMI-1640培养基培养,鸡胚成纤维细胞株DF1用含10%胎牛血清、1%双抗的DMEM培养基培养。细胞置于37℃、5%CO2饱和湿度培养箱中按照常规方法培养。
1.3 病毒滴度测定
感染复数(multiplicity of infection, MOI)为0.01的NDV/FMW感染WM3248细胞株和WM3248/vemurafenib细胞株,收集上清液进行梯度稀释,用DF-1细胞株进行病毒滴度检测。
1.4 CCK8检测细胞存活率
将黑色素瘤细胞分为4组,WM3248CON组,WM3248NDV/FMW组,WM3248/vemurafenibCON组,WM3248/vemurafenibNDV/FMW组。除对照组外,其他组用MOI=1的NDV/FMW感染黑色素瘤细胞1h后,更换2%RPMI-1640培养基,将96孔板置于培养箱继续培养24、48、72h,每孔设5个复孔。待培养时间结束后向待测孔中加入10μL的CCK-8溶液,将细胞培养板放回培养箱中继续孵育1~4h。用酶标仪测定细胞培养板在450nm处的光密度值(D450),并计算分析。
1.5 Western印迹法检测凋亡及自噬相关蛋白变化
WM3248细胞株和WM3248/vemurafenib细胞株10×105个/皿,分别设为对照组与NDV/FMW组,MOI=1的NDV/FMW感染1h后,每皿更换3mL无血清RPMI-1640后放入培养箱继续培养。NDV/FMW感染24、48h后,收集上清液浓缩至100μL,并收集细胞裂解液,用考马斯亮蓝法测定裂解液中的蛋白浓度。取30μg蛋白用10%的分离胶和4%的浓缩胶进行SDS-PAGE电泳,电泳完毕后,将蛋白转印到NC膜上,5%的脱脂奶粉封闭2h,然后用兔抗人cleaved Caspase-3抗体,兔抗人PARP抗体,兔抗人GAPDH抗体,鼠抗人HN抗体,兔抗人HMGB1抗体,鼠抗人HSP90抗体4℃冰箱孵育过夜,用TBST洗涤2次,每次10min,后加入1∶10000的二抗室温孵育1h,用TBST洗涤2次后显影。
1.6 流式细胞术检测CRT表达
MOI=1的NDV/FMW处理WM3248细胞株和WM3248/vemurafenib细胞株48h,胰酶消化后PBS洗涤2次,加入1∶200稀释的兔抗人CRT多克隆抗体,冰上孵育1~2h,PBS洗涤,加入1∶1000的羊抗兔AlexaFluor 488抗体,冰上避光孵育30min,PBS洗涤,流式细胞术检测细胞表面CRT的表达。
1.7 激光共聚焦检测细胞表面CRT的表达
MOI=1的NDV/FMW感染WM3248细胞株和WM3248/vemurafenib细胞株48h,PBS洗涤2次,4%多聚甲醛固定15min后PBS洗涤2遍,加入1∶75稀释的兔抗人CRT多克隆抗体和鼠抗人HN抗体,孵育2~3h,PBS洗涤,加入1∶1000的羊抗兔Alexa Fluor 488抗体和羊抗鼠Alexa Fluor 568抗体,孵育30min,PBS洗涤后DAPI染色,清洗2次,激光共聚焦显微镜检测细胞表面CRT的表达。
1.8 荧光素酶法检测ATP释放
WM3248细胞株和WM3248/vemurafenib细胞株(1×105/孔)铺于12孔板,MOI=1的NDV/FMW感染48h后,收集上清液。根据ENLITEN ATP试剂盒说明书检测上清液中ATP的含量。
1.9 统计学处理

2 结 果
2.1 NDV/FMW在WM3248/vemurafenib细胞株中有效复制
通过病毒滴度检测考察病毒在细胞内的复制情况。NDV/FMW感染WM3248/vemurafenib细胞株24、48、72h后,病毒滴度逐渐增加。结果表明,NDV/FMW在WM3248/vemurafenib细胞株内有效复制,见图1。
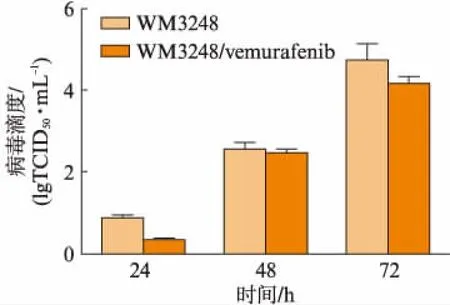
图1 NDV/FMW在WM3248/vemurafenib 细胞株中有效复制Fig.1 NDV/FMW effectively replicated in WM3248/ vemurafenib cell line
2.2 NDV/FMW降低WM3248/vemurafenib细胞株存活率并促进细胞凋亡
NDV/FMW感染WM3248/vemurafenib细胞株24、48、72h后,细胞存活率逐渐降低(P<0.001),见图2A。Western印迹法检测凋亡相关蛋白变化,HN为NDV/FMW感染指示蛋白。NDV/FMW感染WM3248/vemurafenib细胞株后,cleaved-caspase-3和cleaved-PARP蛋白在48h明显增加,见图2B。上述结果表明,NDV/FMW显著降低WM3248/vemurafenib细胞株存活率并促进凋亡。
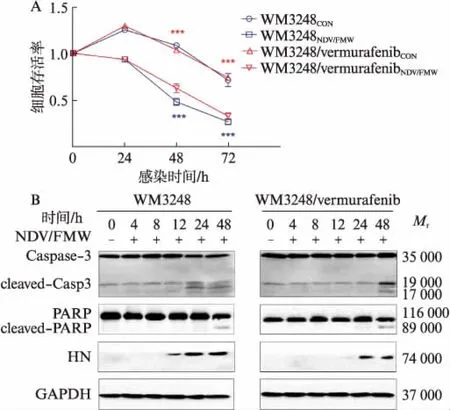
图2 NDV/FMW显著降低WM3248/vemurafenib 细胞株存活率并促进耐药细胞凋亡Fig.2 NDV/FMW significantly reduced cell viability and induced apoptosis in WM3248/vemurafenib cellsA: NDV/FMW感染WM3248/vemurafenib细胞株24、48、72h后,细胞存活率逐渐降低,***P<0.001,亲本细胞用蓝色*代表,耐药细胞用红色*代表;B: Western印迹法检测NDV/FMW对WM3248/vemurafenib凋亡的影响
2.3 NDV/FMW诱导WM3248/vemurafenib细胞株免疫原性死亡相关分子表达或释放
NDV/FMW感染WM3248/vemurafenib细胞株48h后,细胞膜CRT阳性细胞数从1.3%±0.84%增至53.2%±2.56%(P<0.001),见图3A。激光共聚焦结果也显示NDV/FMW感染耐药细胞后,细胞膜表面CRT表达比对照组明显增加,HN也表达增加,见图3B。与对照组相比,NDV/FMW感染WM3248/vemurafenib细胞株后培养液上清液中的HMGB1和HSP90显著释放,ATP分泌增加,差异有统计学意义(P<0.05)。但NDV/FMW感染WM3248细胞48h后ATP比对照组有所消耗(P<0.01),见图3C、D。结果表明NDV/FMW诱导WM3248/vemurafenib细胞株表达或释放免疫原性死亡相关分子。
3 讨 论
新城疫病毒属溶瘤病毒的一种,目前应用于肿瘤治疗的MTH68/H、PV-701、73-T、90D和FMW等毒株均具有良好的溶瘤性[12-13]。既往研究已经证明,NDV/FMW可在神经胶质瘤亲本细胞株U251、肺癌顺铂(DDP)耐药细胞株A549/DDP和肺癌紫杉醇(PTX)耐药细胞株A549/PTX中复制,抑制肿瘤细胞存活率并诱导细胞凋亡或自噬[14-15]。本研究发现,NDV/FMW不仅对恶性黑色素瘤WM3248细胞株有杀伤作用,也对WM3248/vemurafenib细胞株起作用。
本研究进一步发现NDV/FMW对WM3248/vemurafenib细胞株发生ICD相关分子表达的影响,包括细胞膜表面CRT表达增加、ATP分泌、HMGB1和HSP90释放。上述分子的变化与ICD的发生密切相关。肿瘤细胞在死亡早期,CRT从内质网迅速翻转到细胞膜表面,肿瘤细胞膜上的CRT可作为一种“eat me”信号被DC细胞识别和结合,促进肿瘤细胞被DC细胞识别和吞噬,多种肿瘤模型中已证实抑制CRT的表达使肿瘤细胞的免疫原性清除[16]。HSP90同样作为“eat me”信号促进抗肿瘤的免疫原性[17]。HMGB1存在于细胞核和细胞质中,在细胞坏死和凋亡后的细胞均可释放HMGB1,HMGB1同样作为危险信号,有介导细胞免疫原性的潜力[17]。凋亡细胞释放出的ATP被认为是“find-me”信号,由此通过ATP的介导将吞噬细胞聚集在凋亡细胞周围,进而促进凋亡细胞的吞噬和清除[18]。但本研究结果提示NDV/FMW感染WM3248细胞后ATP未见释放增加。既往研究表明,ICD相关的ATP分泌是涉及多种分子的复杂途径,如自噬(ATG 5、ATG 7和BCN 1)、溶酶体外吞(LAMP 1、VAMP 1)、凋亡(Caspases)、膜泡(ROCK 1,肌球蛋白Ⅱ)和质膜透性(Panx 1)等[19]。这些系统中任何分子的缺失都可造成细胞分泌的ATP减少,这意味着细胞自噬、溶酶体功能、caspase活化、膜泡和质膜通透性的改变可能对WM3248细胞ATP的分泌产生影响,需做进一步探讨。
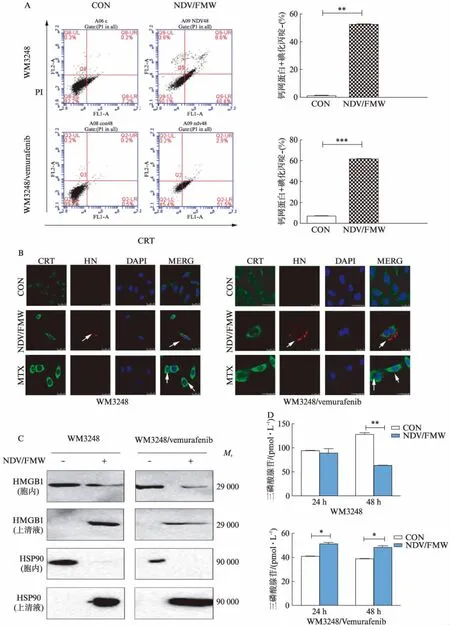
图3 NDV/FMW诱导WM3248/vemurafenib细胞株免疫原性死亡相关分子表达或释放Fig.3 NDV/FMW induced the expression or release of immunogenic cell death related molecules in WM3248/vemurafenib cellsA: 流式细胞术检测耐药细胞表面CRT百分比,**P<0.01,***P<0.001;B: 激光共聚焦检测细胞表面绿色荧光增强,绿色荧光代表CRT,红色荧光代表NDV/FMW特异性HN蛋白,蓝色荧光代表DAPI,米托蒽醌(mitoxantrone,MTX)为阳性对照;C: Western印迹法检测耐药细胞上清液中HMGB1和HSP90的释放;D: 荧光素酶法检测ATP释放,*P<0.05,**P<0.01
本研究不仅在细胞水平上验证NDV/FMW对WM3248/vemurafenib细胞株有细胞毒作用,还进一步证明NDV/FMW可诱导WM3248/vemura-fenib细胞株ICD相关分子表达,进而增强耐药细胞的免疫应答,为NDV/FMW作为恶性黑色素瘤vemurafenib耐药治疗的可行方案提供实验依据。后期实验将进一步研究影响ATP分泌的相关因素,构建恶性黑色素瘤vemurafenib耐药的荷瘤小鼠,进一步探讨NDV/FMW对vemurafenib耐药的体内免疫机制。
猜你喜欢
昆明医科大学学报(2022年8期)2022-07-31
昆明医科大学学报(2021年8期)2021-08-13
昆明医科大学学报(2021年3期)2021-07-22
实用肿瘤学杂志(2020年6期)2020-12-09
中西医结合肝病杂志(2020年2期)2020-10-27
祝您健康·文摘版(2019年1期)2019-05-14
江苏农业科学(2016年10期)2017-02-05
江苏农业科学(2015年11期)2016-01-27
医学研究杂志(2015年9期)2015-07-01
江苏农业科学(2014年10期)2014-11-22
