
桂林岩溶石山不同坡向灌丛植物多样性与土壤环境因子的关系
2019-09-10 07:22:44盘远方李娇凤黄昶吟刘润红姜勇陆志任梁士楚
广西植物 2019年8期
盘远方 李娇凤 黄昶吟 刘润红 姜勇 陆志任 梁士楚


摘要: 該研究以桂林岩溶石山灌丛植物为对象,分析了灌丛群落物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数与土壤环境因子随坡向(阴坡-阳坡)梯度的变化规律。结果表明:(1)土壤含水量、土壤温度、土壤pH和土壤全磷均对植物群落类型及物种分布具有显著的影响,坡向对植物群落类型及物种分布具有极显著的影响;而土壤全氮、土壤有效氮和土壤有机碳对植物群落类型及物种分布则均无显著影响。(2)除Pielou均匀度指数在阴坡与阳坡差异不显著外,物种丰富度指数、Shannon-Wiener指数对不同坡向微生境均存在显著差异。其中,物种丰富度指数和Shannon-Wiener指数在不同坡向上均表现出相同的变化趋势,即阴坡大于阳坡。(3)除土壤pH值和土壤全磷含量差异不显著外,坡向对土壤含水量、土壤温度、土壤全氮含量、土壤有效氮含量、土壤有机碳含量均产生显著的影响。其中,土壤含水量、土壤有效氮含量、土壤有机碳含量则为阴坡大于阳坡,而土壤温度和土壤全氮含量为阴坡小于阳坡。(4)多元逐步回归分析表明,在阴坡上,各土壤环境因子对物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数作用均不显著;在阳坡上,物种丰富度指数与土壤温度呈显著负相关、与土壤有机碳含量呈显著正相关;Shannon-Wiener指数与土壤含水量和土壤有机碳含量呈显著正相关;Pielou均匀度指数与土壤有机碳含量呈显著正相关、与土壤全磷含量呈显著负相关。
关键词: 坡向, 物种多样性, 土壤环境因子, 岩溶石山
中图分类号: Q948文献标识码: A文章编号: 1000-3142(2019)08-1115-11
Abstract: In this study, we took the shrub plants of different slopes in Guilin karst hills as the research object. The relationship between species diversity (species richness index, Shannon-Wiener index and Pielou evenness index) of shrubs and soil environmental factors along with sloped (shady-sunny) gradients in the karst hills of Guilin were analyzed. Our results were as follows: (1) Soil water content, soil temperature, soil pH and soil total phosphorus have significant effects on plant community types and species distribution, and aspect has a very significant impact on plant communities and species distribution. Soil total nitrogen, soil available nitrogen and soil organic carbon had no significant effects on plant community types and species distribution. (2) Except for the Pielou evenness index, the difference between the shady slope and the sunny slope was not significant, species richness index and Shannon-Wiener index had significant differences in different slope microhabitats. Among them, the species richness index and the Shannon-Wiener index showed the same trend in different slopes, that is, the shady slope was larger than the sunny slope. (3) Except for the difference between soil pH and soil total phosphorus content was not significant, the slope aspect had a significant impact on soil water content, soil temperature, soil total nitrogen content, soil available nitrogen content, and soil organic carbon content. Among them, soil water content, soil available nitrogen and soil organic carbon content were shady slope greater than the sunny slope, while soil temperature and soil total nitrogen content were shady slope less than the sunny slope. (4) Multiple stepwise regression analysis showed that on the shady slope, effects of soil environmental factors on species richness index, Shannon-Wiener index and Pielou evenness index were not significant. On the sunny slope, there was a significantly negative correlation between species richness index and soil temperature, and a significantly positive correlation with soil organic carbon content; Shannon-Wiener index was significantly positively correlated with soil water content and soil organic carbon content; Pielou evenness index was significantly positively correlated with soil organic carbon content and negatively correlated with soil total phosphorus content.
Key words: sloped aspect, species diversity, soil environmental factors, karst hills
生物多样性(biodiversity)是指一定时空范围内生物(动物、植物、微生物等)与环境形成的生态复合体以及与此相关的各种生态过程的总和,包括遗传多样性、物种多样性、生态系统多样性和景观多样性4个层次,四者是密不可分、不可或缺的统一体(Zang et al.,2009;吕刚等,2017)。物种多样性是生物多样性在物种水平上的表现形式,是物种丰富度和分布均匀性的综合反映,可以直接或间接地体现群落结构类型、组织水平、发展阶段、稳定程度和生境差异,反映生物群落在组成、结构、功能和动态等方面的异质性(李旭华等,2013;彭羽等,2015)。因此,深入探究物种多样性的变化规律及其影响因子,揭示物种多样性与环境因子的关系,可以更好地评价群落组成、结构及其发展变化,对保护生物多样性具有重要意义(Cadotte et al.,2011;Isbell et al.,2011)。
坡向作为一种重要的地形因素,控制了太阳辐射和降水的空间再分配,往往能够营造出局部的微气候生境,导致光辐射、光照时长、大气温度、土壤水分和土壤养分等环境因子产生生境梯度变化,进而影响着植物群落类型的组成及其物种多样性(段贝贝等,2016)。朱云云等(2016)研究发现坡向对土壤速效养分分布状况具有较强烈的影响,阳坡土壤速效养分含量显著大于阴坡,同时植物在不同坡向环境梯度下作出积极响应,通过选择性的改变根系生长,从而增加阴坡对土壤速效养分的吸收。温璐等(2011)研究发现土壤养分含量为阴坡大于阳坡且在阴坡上土壤全氮、土壤pH值分别与物种丰富度指数呈显著的负相关、与Shannon-Wiener指数呈显著的正相关,土壤含水量与Pieiou均匀度指数呈显著的正相关(刘旻霞和王刚,2013);在阳坡上,光照强度、土壤温度和土壤pH值与物种丰富度指数和Pieiou均匀度指数呈显著的负相关(刘旻霞和王刚,2013)。除坡向对土壤环境因子影响显著外,植被类型对土壤环境因子也产生重要影响。刘海江和郭柯(2003)对浑善达克沙地丘间低地植物群落的分类和排序的研究中得出对其植物分布起主要影响的环境因子为土壤可溶性钠和pH值。因此,研究物种多样性随坡向梯度的变化规律及其与环境因子的关系,对认识不同坡向上植物群落的形成及其对环境的适应机制具有重要意义。
桂林位于广西东北部,是广西岩溶石山地貌集中分布的地区之一(刘彦随等,2006),也是我国石漠化区水土治理和植被恢复的重点区域(李阳兵等,2004)。其特征为地形破碎、岩石裸露率高、成土条件差、土层浅薄、土被不连续、渗漏严重和地表水缺乏(李先琨等,2003)。长期以来,该地区由于受人为活动干扰和破坏,导致桂林岩溶石山植被生态系统发生逆行演替,除了少数村庄后山残存小片落叶阔叶常绿阔叶混交林以外,大多为灌丛或藤刺灌丛(马姜明等,2009)。其中,灌丛作为岩溶植被恢复过程中的一种典型的植被类型,具有萌生力强、旱生、岩生、嗜钙等特点,是该地区植被从草丛恢复到乔木林的关键过渡阶段,在群落演替过程中扮演着重要的角色(屠玉麟,1995;He et al.,2008)。然而,目前关于岩溶石山灌丛植物多样性及其与土壤环境因子关系的研究尚鲜见报道。鉴于此,本研究以桂林岩溶石山灌丛植物为研究对象,分析了桂林岩溶石山灌丛群落物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数与土壤环境因子随坡向(阴坡-阳坡)梯度的变化规律及其与环境因子的关系,以期揭示不同坡向上植物群落的形成及其对环境的适应机制,进而为岩溶退化生态系统的植被恢复与重建等提供科学依据。
1研究区域概况
研究区位于广西壮族自治区桂林市临桂区西南部,为典型的岩溶地貌区,地理坐标为110°12′—110°48′ E、24°21′—25°05′ N。该区域属中亚热带湿润季风气候区,全年光照充足,气候温和,四季分明,雨量充沛,雨热基本同期,年均气温为19 ℃,最热的8月份平均气温为28 ℃,最冷的1月份平均气温为8 ℃,全年无霜期为309 d,年均降雨量为1 856.7 mm,降雨量年分配不均,秋、冬季干燥少雨,年均蒸发量为1 458.4 mm,无霜期长达300余天。该地区地表形态复杂多样,生态环境差异明显,主要以石灰岩、白云岩等碳酸盐类岩石为主,成土速度慢,形成的土壤浅薄,土壤钙含量高,且土被不连续,岩石裸露率极高,地形破碎,土壤蓄水能力弱,土壤含水量低,受这些特殊的环境条件影响,植物生长缓慢,其适生植物具有嗜钙性、耐旱性、石生性等特点。研究区植被类型以灌丛为主,主要的优势种有檵木(Loropetalum chinense)、红背山麻杆(Alchornea trewioides)、山麻桿(A. davidii)、牡荆(Vitex negundo)、紫薇(Lagerstroemia indica)、南方荚蒾(Viburnum fordiae)、铜钱树(Paliurus hemsleyanus)和千里香(Murraya paniculata)等。
2研究方法
2.1 群落学调查
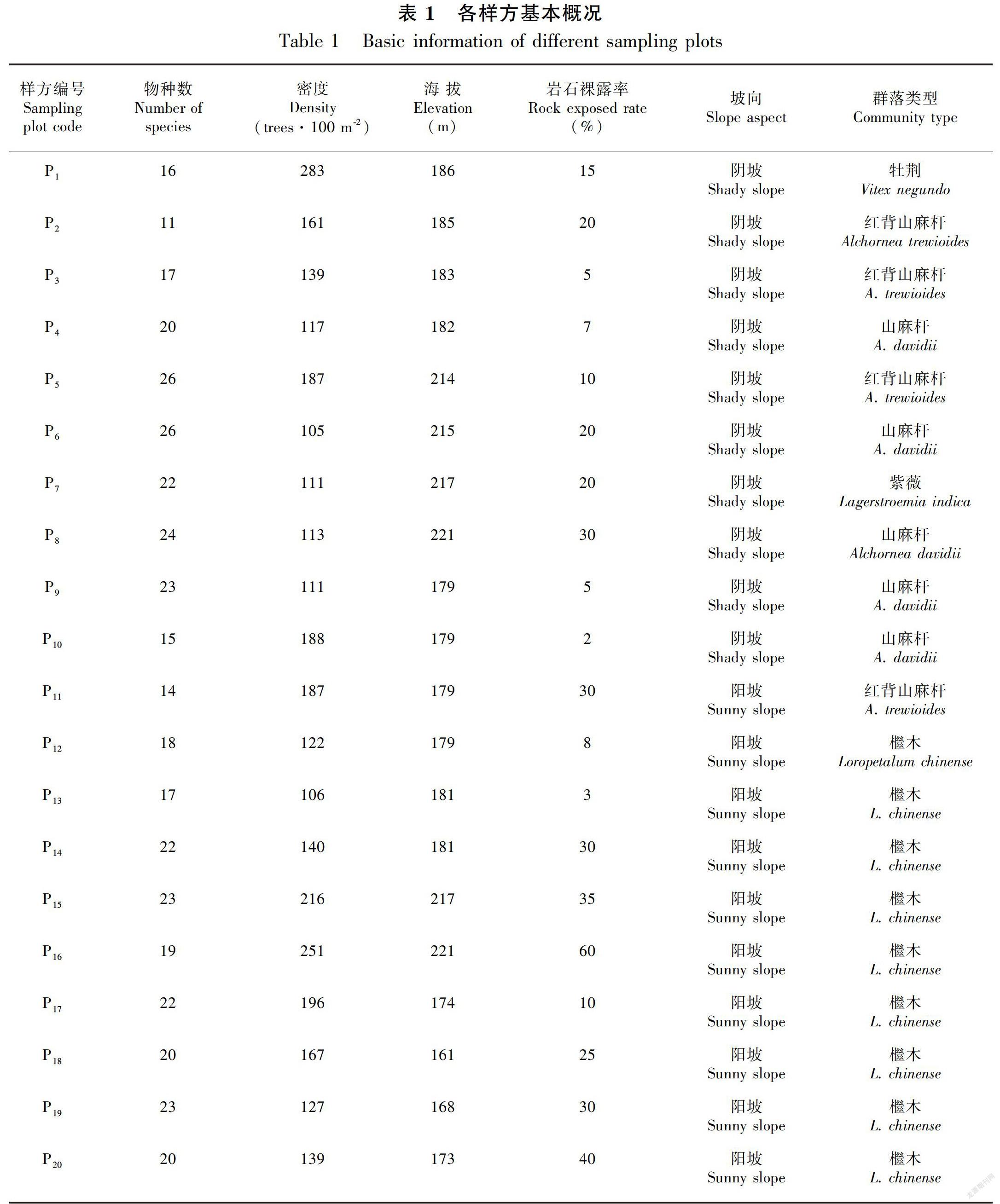
经过实地勘察,2016年7—11月分别在桂林市临桂区会仙镇的阳家村和李家村,以坡向为主导因子,在阴坡和阳坡各设置10个大小为10 m×10 m的样方,合计20个样方。采用每木调查法对样方内灌丛植物进行全面调查,记录样方内基径≥1 cm的所有木本植物的种名、株高、胸径、基径、冠幅、空间坐标等。同时,记录各样方的经纬度、海拔、坡向、岩石裸露率等生境特征,各样方基本情况如表1所示。本次共调查了3 166株植物个体,隶属98种。
2.2 土壤样品的采集及测定
2.2.1 土壤样品采集土壤化学性质采用“梅花五点法” 采样(卜文圣等,2013),在每个10 m×10 m样方的四个顶点及中心位置,清除土壤表层枯枝落叶和腐殖质层后,用土钻钻取0~20 cm的表层土壤1 kg,混合均匀后装入密封袋带回实验室, 置于室内阴凉通风处自然干燥,待风干后,剔除其中的根系、石块、钙核及动植物残体等杂物后用粉碎机粉碎,过孔径为2 mm的筛后用于土壤有效氮的测定。另取风干土壤样品粉碎,过孔径为0.25 mm的筛后用于土壤全氮含量、土壤全磷含量和土壤有机碳含量的测定。
2.2.2 土壤样品的测定将每个10 m×10 m样方划分成4个5 m×5 m的小样方并在其中心位置处用土壤三参数仪(W.E.T Sensor Kit)测定土壤含水量(SWC,%)和土壤温度(Tem,℃)。土壤pH值采用电导法测定(水土比为 2.5∶1);有机碳含量(TOC,g·kg-1)采用重铬酸钾容量法测定;土壤全氮含量(TN,g·kg-1)采用全自动凯氏定氮仪(KJELTECTM8400)测定;土壤有效氮含量(AN,mg·kg-1)采用碱解扩散法测定;土壤全磷含量(TP,g·kg-1)采用钼锑抗比色法测定。每个土壤样品化学及物理性质均重复测定3次后取其平均值作为本研究分析的数据。
2.3 数据分析
计算每个样方灌丛植物物种的重要值[IV灌木=(相对密度+相对频度+相对盖度)/3](习新强等,2011),基于样方内物种重要值计算的结果进行大小排序确定样方的植物群落类型种类(徐建霞和王建柱,2018),并用CCA对植物群落类型和环境因子进行了排序。物种多样性指数主要采用物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数。各指数计算公式如下:
采用Wilcoxon加符秩检验方法对不同坡向(阴坡-阳坡)梯度上物种多样性指数(物种丰富度指数、Shannon-Wiener指数、Pielou均匀度指数)和土壤环境因子进行差异显著性检验。为进一步探究哪些环境因子对物种多样性起重要影响,以7个土壤环境因子作为自变量,3个物种多样性指数(物种丰富度指数、Shannon-Wiener指数、Pielou均匀度指数)作为因变量进行多元逐步回归分析,根据AIC(赤池信息量准则)、R2(决定系数)和对模型判断P的统计检验得出在不同坡向上显著的物种多样性指数与对应环境因子的回归关系。其中,AIC表示用于衡量回归模型拟合优劣的一种标准,AIC值越小说明其拟合度越高;R2表示回归模型拟合优度判定系数,反映因变量的全部变异能通过回归关系被自变量解释的比例;P值即概率,是用来判定假设检验结果的一个参数,P值越小表明结果越显著。所有数据分析及作图均在R 3.3.1软件(R Development Core Team,2016)中完成。
3结果与分析
3.1 植物群落分布与土壤环境因子的分析
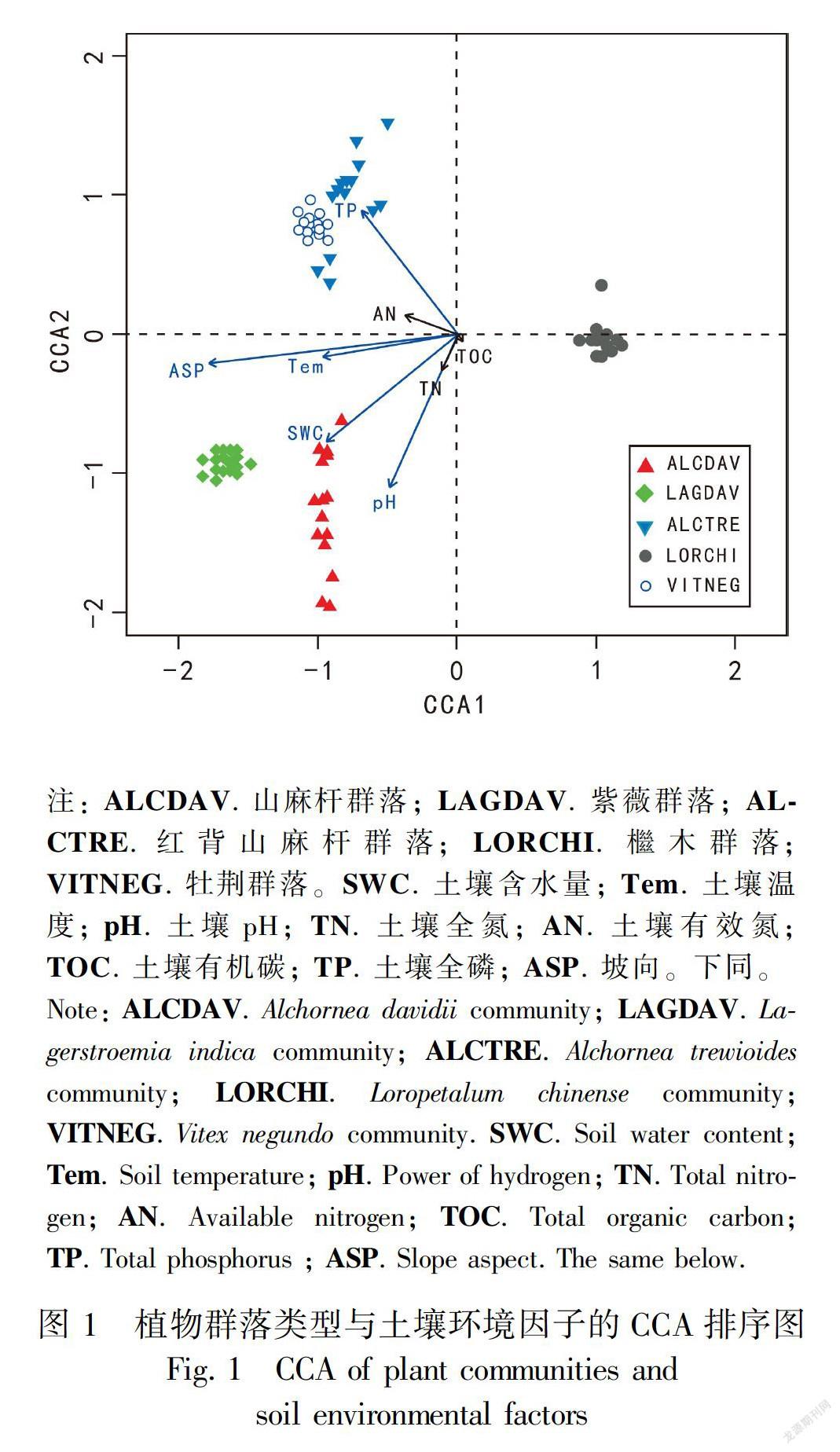
CCA排序结果表明,土壤含水量、土壤温度、土壤pH和土壤全磷均对植物群落分布产生显著影响(P<0.05),坡向对植物群落分布产生极显著影响(P<0.01)。土壤全氮、土壤有效氮和土壤有机碳对植物群落分布均无显著影响(P>0.05)(表2,图1)。
3.2 不同坡向物种多样性指数的比较
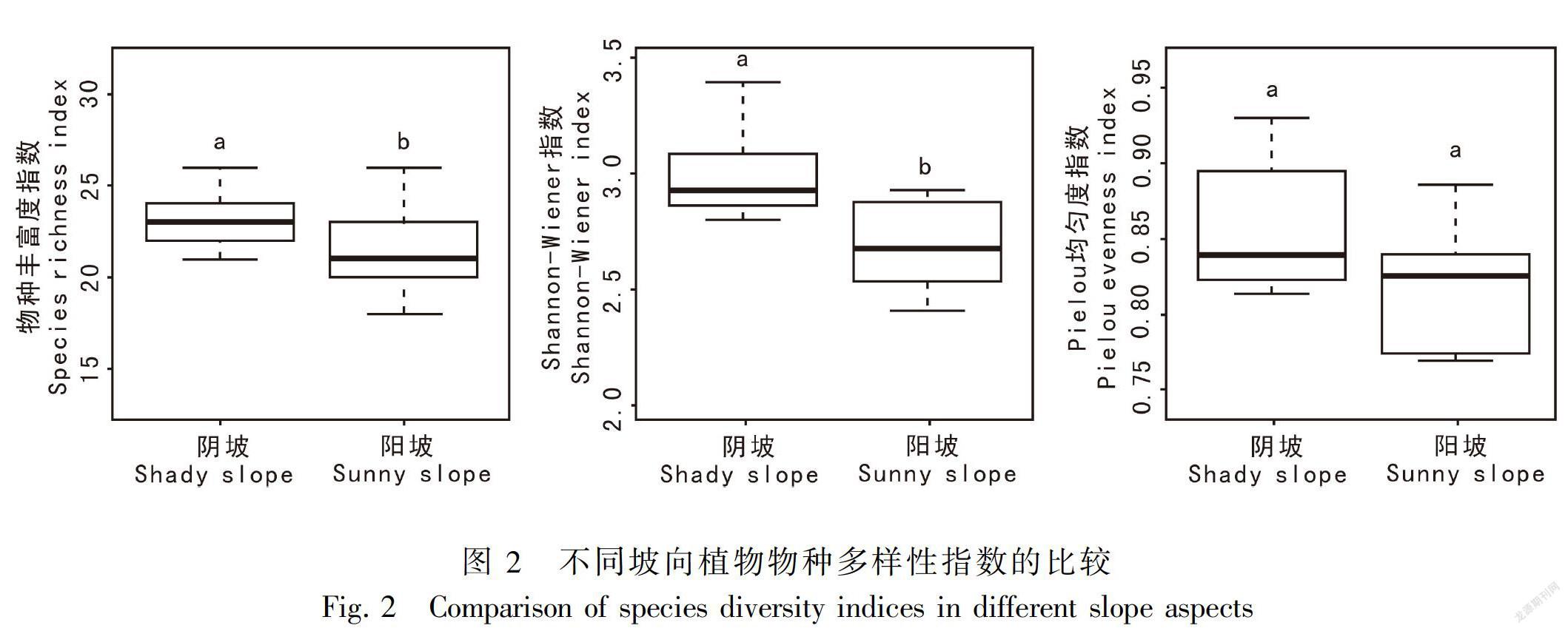
如图2所示,除Pielou均匀度指数在不同坡向上差异不显著外(P>0.05),物种丰富度指数、Shannon-Wiener指数对不同坡向微生境均产生显注: ALCDAV. 山麻杆群落; LAGDAV. 紫薇群落; ALCTRE. 红背山麻杆群落; LORCHI. 檵木群落; VITNEG. 牡荆群落。SWC. 土壤含水量; Tem. 土壤温度; pH. 土壤pH; TN. 土壤全氮; AN. 土壤有效氮; TOC. 土壤有机碳; TP. 土壤全磷; ASP. 坡向。下同。
3.3 不同坡向土壤环境因子的比较
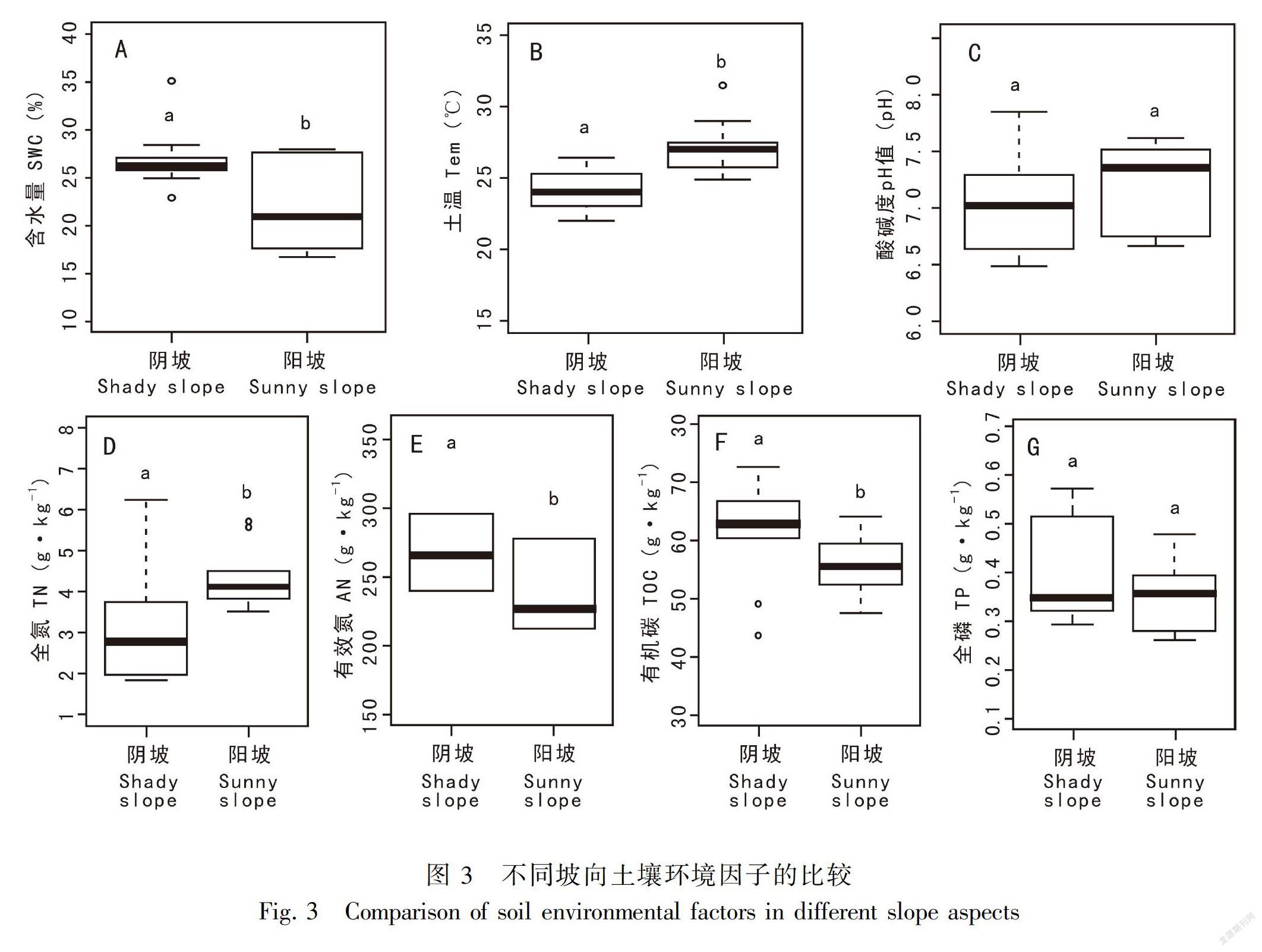
如圖3所示,除了土壤pH值和土壤全磷含量差异不显著外(P>0.05),坡向(阴坡-阳坡)对其它土壤环境因子均产生显著的影响(P<0.05)。其中,土壤含水量、土壤有效氮含量、土壤有机碳含量为阴坡大于阳坡,而土壤温度和土壤全氮含量为阴坡小于阳坡。
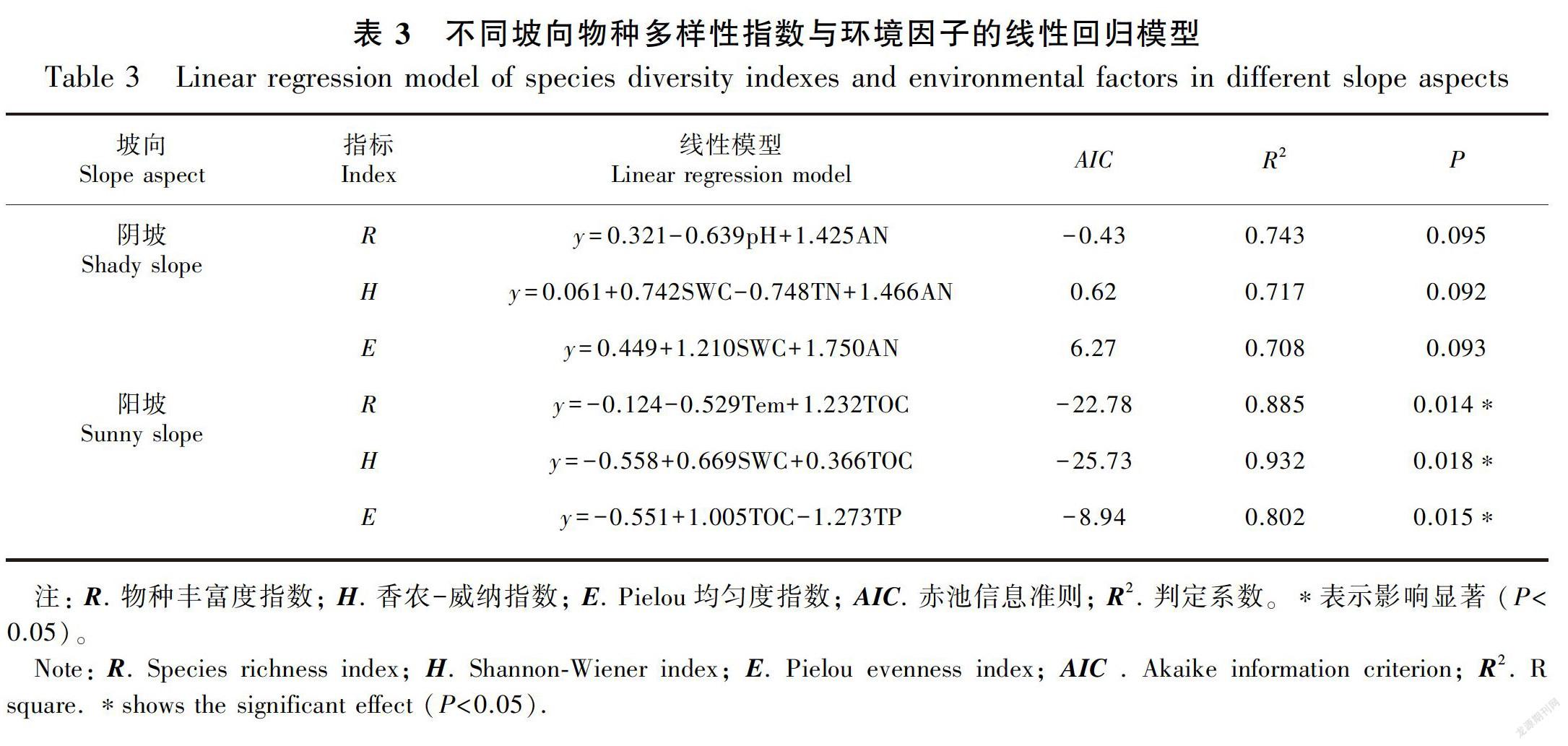
3.4 不同坡向物种多样性指数与土壤环境因子的回归分析
多元逐步回归分析结果表明,在阴坡上,土壤环境因子对物种丰富度指数、Shannon-Wiener指数指数和Pielou均匀度指数影响均不显著(P>0.05)(表3)。在阳坡上,对物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数存在显著影响的土壤环境因子是土壤含水量、土壤温度、土壤有机碳含量和土壤全磷含量(P<0.05)(表3)。
4讨论
4.1 不同植物群落类型与土壤环境因子的关系
植物生长过程是植物对土壤环境因子的不断适应和改善的过程, 而土壤环境因子的不断改善和恢复又促使植物群落类型及结构多样化。CCA排序结果表明,坡向、土壤含水量、土壤温度、土壤pH和土壤全磷均对植物群落分布产生显著的影响。坡向是重要的地形因子,影响着地表接受的太阳辐射量,而太阳辐射则是决定近地面气温、土壤湿度、土壤温度和养分循环的主要因子。其中,土壤含水量是植物生命活动的直接水源,对植物生长发育有着重要作用,植物所吸收的养分元素必须溶于溶液中才能被植物吸收利用,而土壤温度作为土壤水资源形成、转化、消耗和能量交换的重要参数,则控制着种子的萌发及幼苗的生长(刘旻霞和王刚,2013)。因此,坡向的空间变化不仅是植物群落组成及分布格局的重要决定因素,而且还使其分布格局表现出一定的变化规律。土壤pH值是土壤重要的化学性质,直接影响植物生长、土壤微生物的活动、酶的活性和土壤的肥力状况。朱媛君等(2016)研究表明,随着土壤pH值变化土壤多酚氧化酶活性受到抑制,土壤多酚氧化酶活性高低与动植物凋落物腐解释放土壤营养元素速率呈正相关。全磷是衡量土壤中各种形态磷素总和的一个指标,也是植物生长发育不可缺少的元素(刘淑娟等,2011)。磷素是植物体内参与碳水化合物的合成,提供植物生长及代谢的能量,同时能促进根系发育,扎根至深层的土壤吸收水分和养分以适应岩溶石山干旱缺水的环境(Zhang et al., 2012)。因此,土壤磷素对植物群落的分布具有重要的影响。综合CCA排序结果可知,决定群落分布的土壤环境因子是以某种复杂的方式综合作用该地区植物群落类型及分布。
4.2 物种多样性指数和土壤环境因子对不同坡向的响应
坡向是影响土壤表层接受太阳光照强度的关键因子,导致不同坡面接受太阳的辐射量不同,促使光照、土壤温度和土壤水分产生高度异质性,对植物生长发育、物种多样性和生态系统功能产生重要影响(刘旻霞和王刚,2013)。本研究发现,物种丰富度指数为阴坡大于阳坡,土壤含水量为阴坡大于阳坡,而土壤温度则为阴坡小于阳坡。受到土壤含水量和土壤温度等的影响,土壤营养的矿化、土壤团聚体稳定性、土壤孔隙度、土壤渗透系数等在不同坡向上会产生巨大的差异(Wang et al.,2008;Huang et al.,2015;朱云云等,2016)。刘旻霞和王刚(2013)研究表明,从阴坡到阳坡,太阳辐射量增加,导致阳坡土壤表层温度较高、土壤水分蒸发量大,土壤含水量减少。同时,阳坡土壤风化严重,增加了土壤表层的孔隙度,加之阳坡光照强度大,地表温度高,水分易于蒸发和漏失,使阳坡土壤水分不易保持,植物对土壤的保水能力下降,土壤的渗透阻力增大,从而导致阳坡土壤保水和持水能力下降(李胜平和王克林,2016)。土壤含水量是限制不同坡向群落构成的最主要因素之一,阴坡水分相对充足,温度较适宜,以及丰富的营养物质能够保障更多的物种在阴坡上生存繁衍。在本研究中,从阴阳坡主要植物群落分布来看,阴坡主要以山麻杆、紫薇和红背山麻杆等耐阴植物为主,而阳坡则主要以檵木等喜阳植物为主,体现了植物对生境的一种适应。Shannon-Wiener指数阴坡显著高于阳坡,土壤有机碳含量和土壤有效氮含量为阴坡大于阳坡。可能原因是由于不同坡向光照强度不同,使植物初级生产力、凋落物量及分解速率、根系分泌物、土壤微生物等方面产生差异。其中,凋落物由易分解的脂肪、糖类、淀粉等成分和难分解的木质素、多酚等成分组成,其分解一方面能增加土壤有机碳的来源及改变土壤理化性质,另一方面能增加土壤微生物的数量及调控微生物对凋落物的分解(温璐等,2011;宋媛等,2013)。土壤温度的升高,土壤微生物活性增加,加速动植物凋落物的分解,有利于土壤有机碳的积累(Kirschbaum,2000)。阳坡的立地条件较为恶劣,即土壤风化和水土流失严重、土壤养分难以富集、光照强度大及土壤水分蒸发较快,加之动植物凋落物有机质输入较少、土壤容重增加及土壤退化,抑制了耐阴植物的正常生长,使阳坡仅能生长一些耐旱耐强光的植物,导致阳坡群落总体物种多样性降低(吴昊等,2012)。本研究调查发现,阴坡灌丛不仅郁闭度较高,而且阴坡土壤含水量和有机碳等养分含量也较高,灌丛林下植物可利用的光资源异质性高,能给耐阴植物提供更多的生长机会,从而使物种Shannon-Wiener指数阴坡高于阳坡,这说明阴坡物种相对阳坡较为丰富,阴坡生态系统更稳定以及物种多样性更高。Pielou均匀度指数阴坡与阳坡差异不明显,这可能是岩溶石山整体立地条件比较恶劣导致阴阳坡各物种组成及个体数无明显的差异,而土壤全氮含量则是阴坡小于阳坡。Cadotte et al.(2011)在加拿大亚伯达省中部利用氮-16同位素示踪技术研究发现,土壤pH值增加促使土壤腐蚀质向氮素转化;而本研究结果则显示阳坡土壤pH值较高,因此阳坡具有较高的全氮含量。阴坡与阳坡的土壤全磷含量无显著差异,主要是因为磷素是一种沉积性元素,由土壤母质类型和成土条件决定,在岩溶石山土壤中的磷素主要与钙质成分作用形成难溶性磷酸钙和磷灰石,其形式比较稳定,不易流失(Allison et al.,2007;刘淑娟等,2011)。
4.3 不同坡向物种多样性指数与土壤环境因子的回归关系
植物群落的物种多样性及生长发育状况不仅取决于自身的生理特性,而且也会受到土壤环境因子的显著影响(李东升和王克林,2016)。多元逐步回归分析表明,在阴坡上,土壤含水量、土壤pH值、土壤有效氮含量和土壤全氮含量对物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数均有影响,但影响均未达到显著的水平。这表明影响物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数的因素较为复杂,除了生境中的环境因子以外,还可能存在一些其他因子影响其变化,如受到光照、气候、降雨量、石漠化程度及群落中植物自身的生物学特性等综合影响。在阳坡上,物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数均与土壤有机碳含量呈显著的正相关。这可能是由于阳坡光照充足,造成阳坡坡面温度较高和土壤水分蒸发快,使阳坡植物凋落物腐解受到阻碍(朱云云等,2016)。土壤有机碳能间接地影响植物群落的结构、功能和生产力,是影响植物群落多样性的关键因子,同时土壤有机碳能提高土壤的粘性,改善土壤的透气性、保水性、抗蚀性等,增加群落的多样性(郭曼等,2010;徐建霞和王建柱,2018)。因此,在阳坡上,物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数均与土壤有机碳含量呈显著的正相关。本研究中,物种丰富度指数与土壤温度呈显著的负相关、Shannon-Wiener指数与土壤含水量呈显著的正相关,这可能是因为阳坡土壤风化严重、表层土壤水分蒸发快,为了适应石生性和耐旱性的特点,植物必须具有发达而强壮的根系才能扎根和生长。植物根系与土壤形成的根系-土壤复合体以及植物根系间的相互缠绕、团聚形成的根系网,能有效地固结、黏聚土壤颗粒促进土壤团聚体的形成(吕刚等,2017)。团聚体的形成有利于土壤对水分吸附增强,形成充满团聚体的吸附水膜,借助土壤团聚体毛管作用保持少量水,促使土壤保水能力变强,从而维持植物的生长(葛晓改等,2012)。本研究中,物种Pielou均匀度指数与土壤全磷含量呈显著的负相关,这可能是由于阳坡土壤风化严重,水土流失严重,岩石裸露率较高、石漠化严重,雨季土壤磷流失明显(吕刚等,2017)。刘淑娟等(2011)研究表明,土壤酸性磷酸酶能促进有机磷的脱磷速度,从而提高磷的有效性,土壤酸性磷酸酶的最适pH值为4.8;而本研究结果则显示阴坡与陽坡土壤pH值均大于4.8,土壤偏中性或者碱性抑制了土壤酸性磷酸酶的活性,降低了磷的有效性。此外,磷素是一种沉积性元素,在岩溶石山土壤中的磷素主要与钙质成分作用形成难溶性磷酸钙和磷灰石,其形式比较稳定,不易被植物吸收利用。因此,在阳坡上Pielou均匀度指数与土壤全磷含量呈显著的负相关。
5结论
通过对岩溶石山不同坡向(阴坡-阳坡)梯度灌丛植物多样性与土壤环境因子关系进行研究。结果显示,影响植物群落分布是土壤环境因子坡向、土壤含水量、土壤温度、土壤pH和土壤全磷共同作用的结果。物种丰富度指数、Shannon-Wiener指数和Pielou均匀度指数均表现为阴坡大于阳坡,表明阴坡的物种更丰富,生态系统更稳定,其维持、繁衍和保护物种多样性的能力也相对较强。土壤含水量、土壤有效氮含量、土壤有机碳含量均为阴坡大于阳坡,土壤温度、土壤全氮含量为阴坡小于阳坡,表明阳坡生境较为恶劣,土壤资源较贫瘠。尽管不同坡向梯度土壤养分含量不同,但不同坡向植物采取不同的生存策略来适应不同坡向梯度的微生境,揭示了微气候生境条件下土壤环境因子对岩溶石山灌丛植物物种多样性的影响,反映了环境筛选对植物群落形成的作用,对岩溶退化生态系统的植被恢复与重建等具有重要的指导意义。
参考文献:
BU WS, ZANG RG, DING Y, et al., 2013. Relationships between plant functional traits at the community level and environmental factors during succession in a tropical lowland rainforest on Hainan Island, South China [J]. Biodivers Sci, 21(3):278-287. [卜文圣, 臧润国, 丁易, 等, 2013. 海南岛热带低地雨林群落水平植物功能性状与环境因子相关性随演替阶段的变化 [J]. 生物多样性, 21(3):278-287.]
CADOTTE MW, CARSCADDEN K, MIROTCHNICK N, 2011. Beyond species: Functional diversity and the maintenance of ecological processes and services [J]. J Appl Ecol, 48: 1079-1087.
DUAN BB, ZHAO CZ, XU T, et al., 2016.Correlation analysis between vein density and stomatal traits of Robinia pseudoacacia in different aspects of Beishan Mountain in Lanzhou [J]. Chin J Plant Ecol, 40(12): 1289-1297. [段貝贝, 赵成章, 徐婷, 等, 2016. 兰州北山不同坡向刺槐叶脉密度与气孔性状的关联性分析 [J]. 植物生态学报, 40(12): 1289-1297.]
GUO M, ZHENG FL, HE WX, et al., 2010. Variation of vegetaon diversity and its relationship with soil nutrient and enzyme activity in lands of different abandoned years in the loess hilly-gully region [J]. Acta Pedol Sin, 47(5): 979-986. [郭曼, 郑粉莉, 和文祥, 等, 2010. 黄土丘陵区不同退耕年限植被多样性变化及其与土壤养分和酶活性的关系 [J]. 土壤学报, 47(5): 979-986.]
GE XG, XIAO WF, ZENG LX, et al., 2012.Relationships between litter substrate quality and soil nutrients in different-aged Pinus massoniana stands [J]. Acta Ecol Sin, 32(3): 852-862. [葛晓改, 肖文发, 曾立雄, 等, 2012. 不同林龄马尾松凋落物基质质量与土壤养分的关系 [J]. 生态学报, 32(3): 852-862. ]
HU YS,YAO XY, LIU YH, 2014. The functional traits of forests at different succession stages and their relationship to terrain factors in Changbai Mountains [J]. Acta Ecol Sin, 34(20): 5915-5924. [胡耀升, 么旭阳, 刘艳红, 2014. 长白山不同演替阶段森林植物功能性状及其与地形因子间的关系 [J]. 生态学报, 34(20): 5915-5924.]
HE XY, WANG KL, ZHANG W, et al., 2008. Positive correlation between soil bacterial metabolic and plant species diversity and bacterial and fungal diversity in a vegetation succession on Karst [J]. Plant and Soil, 307(1-2): 123-134.
ISBELL F, CALCAGNO V, HECTOR A, et al., 2011. High plant diversity is needed to maintain ecosystem services [J]. Nature, 477: 199-202.
KIRSCHBAUM UF, 2000. Will changes in soil organic carbon act as a positive or negative feedback on global warming? [J]. Biogeochemistry, 48(1):21-51.
LI SP, WANG KL, 2016. Seasonal distribution of soil nutrients and their response to the plant diversity of karst mountain grassland [J]. J Soil Water Conserv, 30(4): 199-205. [李勝平, 王克林, 2016. 桂西北喀斯特山地草地土壤养分季节变化规律及其对植被多样性的响应 [J]. 水土保持学报, 30(4): 199-205.]
LI SP, WANG KL, 2016. Effect ofhuman disturbance on siol nutrients and plant diversity of grassland in karst mountain [J]. Res Soil Water Conserv, 23(5): 20-27. [李胜平, 王克林, 2010. 人为干扰对桂西北喀斯特山地植被多样性及土壤养分分布的影响 [J]. 水土保持研究, 23(5): 20-27.]
LI XK, SU ZM, L SH, et al., 2003. The spatial pattern of natural vegetation in the karst regions of Guangxi and the ecological signality for ecosystem rehabilitation and reconstructio [J]. Mt Res, 21(2): 129-130. [李先琨, 苏宗明, 吕仕洪, 等, 2003. 广西岩溶植被自然分布规律及对岩溶生态恢复重建的意义 [J]. 山地学报, 21(2): 129-130.]
LI XH, DENG YL, ZHANG F, et al., 2013. Species diversity of forest communities in Pangquangou Nature Reserve, Shanxi of China [J]. Chin J Ecol, 32(7): 1667-1673. [李旭华, 邓永利, 张峰, 等, 2013. 山西庞泉沟自然保护区森林群落物种多样性 [J]. 生态学杂志, 32(7): 1667-1673.]
LI YB, WANG SJ, RONG L, 2004. Prospect of the study on rock desertification and its restoration in Southwest Karst mountains [J]. Chin J Ecol, 23(6): 84-88. [李阳兵, 王世杰, 容丽, 2004. 西南岩溶山地石漠化及生态恢复研究展望 [J]. 生态学杂志, 23(6): 84-88.]
LIU MX, WANG G, 2013. Responses of plant community diversity and soil factors to slope aspect in alpine meadow [J]. Chin J Ecol, 32(2): 259-265. [刘旻霞, 王刚, 2013. 高寒草甸植物群落多样性及土壤因子对坡向的响应 [J]. 生态学杂志, 32(2): 259-265.]
LIU SJ, ZHANG W, WANG KL, et al., 2011. Spatiotemporal heterogeneity of topsoil nutrients in karst peak-cluster depression area of Northwest Guangxi, China [J]. Acta Ecol Sin, 31(11): 3036-3043. [刘淑娟, 张伟, 王克林, 等, 2011. 桂西北喀斯特峰丛洼地表层土壤养分时空分异特征 [J]. 生态学报, 31(11): 3036-3043.]
LIU YS, DENG XS, HU YC, 2006. Rocky land degradation and poverty alleviation strategy in Guangxi karst mountainous area [J]. Mt Res, 24(4): 228-233. [刘彦随, 邓旭升, 胡业翠, 2006. 广西喀斯特山区土地石漠化与扶贫开发探析 [J]. 山地学报, 24(4): 228-233.]
LIU HJ, GUO K, 2003. Classification and ordination analysis of plant communities in Inter-dune lowland in Hunshandak Sandy Land [J]. Acta Ecol Sin, 23(10): 2163-2169. [刘海江, 郭柯, 2003. 浑善达克沙地丘间低地植物群落的分类与排序 [J]. 生态学报, 23(10): 2163-2169.]
L G, WANG T, LI YX, et al., 2017. Herbaceous plant diversity and soil physicochemical properties on the regeneration slash of Pinus sylvestris var. mongolica [J]. Acta Ecol Sin, 37(24): 8294-8303. [吕刚, 王婷, 李叶鑫, 等, 2017. 樟子松固沙林更新迹地草本植物多样性及其对土壤理化性质的影响 [J]. 生态学报, 37(24): 8294-8303.]
MA JM, LIANG SC, LIANG YM, et al., 2009. Aboveground biomass and its allocation of main shrub types in karst hills of Guilin, China [J]. J Guangxi Norm Univ (Nat Sci Ed), 27(4): 96-98. [马姜明, 梁士楚, 梁月明, 等, 2009. 桂林岩溶石山主要灌丛类型地上生物量及分配特征 [J]. 广西师范大学学报 (自然科学版), 27(4): 96-98.]
PENG Y, QING FT, MI K, et al., 2015. Study progress on spatial scale effects and coupling relationships of different levels in biodiversity [J]. Acta Ecol Sin, 35(2): 577-583. [彭羽, 卿凤婷, 米凯, 等, 2015. 生物多样性不同层次尺度效应及其耦合关系研究进展 [J]. 生态学报, 35(2): 577-583.]
RU HL, ZHANG HD, JIAO F, et al., 2016. Impact of micro-landform on grassland plant community structure and function in the hilly Loess Plateau region, China [J]. Chin J Appl Ecol, 27(1): 25-32. [汝海丽, 张海东, 焦峰, 等, 2015. 黄土丘陵区微地形对草地植物群落结构组成和功能特征的影响 [J]. 应用生态学报, 27(1): 25-32.]
SONG Y, ZHAO XZ, MAO ZJ, 2013.SOC decomposition of four typical broad-leaved Korean pine communities in Xiaoxingan Mountain [J]. Acta Ecol Sin, 33(2): 443-453. [宋媛, 赵溪竹, 毛子军, 等, 2013. 小兴安岭4种典型阔叶红松林土壤有机碳分解特性 [J]. 生态学报, 33(2): 443-453.]
TU YL, 1995. A analisis of flora and ecological characteristics of karst scrubs in Guizhou Province [J]. J Guizhou Norm Univ(Nat Sci Ed), 13(3): 1-8. [屠玉麟, 1995. 贵州喀斯特灌丛区系与生态特征分析 [J]. 贵州师范大学学报(自然科学版), 13(3): 1-8.]
XU JX, WANG JZ, 2018.Correlation analysis between vegetation and soil physical-chemical factors in the Xiangxi River water-level fluctuation zone of the Three Gorges Reservoir area [J]. Chin J Ecol, 37(12): 3661-3669. [徐建霞, 王建柱, 2018. 三峡库区香溪河消落带植被群落特征与土壤环境相关性 [J]. 生态学杂志, 37(12): 3661-3669.]
VESTERDAL L, CLARKE N, SIGURDSSON BD, 2013. Do tree species influence soil carbon stocks in temperate and boreal forests? [J]. For Ecol Manag, 3(9): 4-18.
WU H, LIU H, ZHANG Y, et al., 2012. Coupling relationship between species diversity of planted Chinese Pine (Pinus tabulae formis Carr.) communities and environment factors in mountain area of Shangluo [J]. Acta Bot Boreal-Occident Sin, 32(2): 377-383. [吴昊, 刘华, 张洋, 等, 2012. 商洛山区人工油松群落物种多样性与环境因子关系研究 [J]. 西北植物学报, 32(2): 377-383.]
ZHU YY, WANG XA, WANG X, et al., 2016. Effect of slope aspect on the functional diversity of grass communities in the Loess Plateau [J]. Acta Ecol Sin, 36(21): 6823-6833. [朱云云, 王孝安, 王贤, 等, 2016. 坡向因子对黄土高原草地群落功能多样性的影响 [J]. 生态学报, 36(21): 6823-6833.]
ZHU YJ, ZHANG PJ, NIU ML, et al., 2016. Soil enzyme activities of the main plant communities in inter-dune lowland of Mu Us sandy land [J]. Chin J Ecol, 35(8): 2014-2021. [朱媛君, 張璞进, 牛明丽, 等, 2016. 毛乌素沙地丘间低地主要植物群落土壤酶活性 [J]. 生态学杂志, 35(8): 2014-2021.]
ZHANG HY, QIAN YB, WU ZU, et al, 2012. Vegetation-environment relationships between northern slope of Karlik mountain and naomaohu basin, east tianshan mountains [J]. Chin Geogr Sci, 22(3): 288-301.
猜你喜欢
测绘学报(2019年11期)2019-11-20 01:31:42
科技创新与应用(2018年36期)2018-01-29 10:30:36
安徽农学通报(2017年11期)2017-06-23 22:21:10
绿色科技(2016年21期)2016-12-27 10:53:26
现代农业科技(2016年20期)2016-12-20 09:17:04
科技创新导报(2016年21期)2016-12-17 12:53:50
现代经济信息(2016年13期)2016-06-17 21:16:45
土壤与作物(2015年3期)2015-12-08 00:46:55
绿色科技(2015年6期)2015-08-05 18:12:16
安徽农学通报(2015年3期)2015-07-30 00:02:28
