
低温胁迫对不同拔节进程冬小麦光合特性的影响
2019-09-10 08:35:06张溪荷刘荣花
种业导刊 2019年7期
张溪荷,刘荣花,李 祯,成 林
(1.中国气象局 河南气象局农业气象保障与应用技术重点开放实验室,河南 郑州 450003;2.河南省气象科学研究所,河南 郑州 450003;3.河南省气象台,河南 郑州 450003)
影响植物生长的因素有很多,如光照、温度、水分和重金属等,其中,低温是对农作物生产影响最大的逆境伤害之一。当植物遭受低温时,其外部形态和生理功能均会发生变化,形态上主要表现为叶片枯萎、凋零等,生理上主要表现为细胞膜功能失调、呼吸作用减弱等。
冬小麦拔节之前,由于受叶鞘保护,幼穗不易遭受低温危害。拔节期是冬小麦幼穗形成的关键时期,此时植株抗寒性较差,但却是强冷空气活动频繁的阶段,因此,冬小麦霜冻害高发。冬小麦幼穗一旦遭遇低温冻害,分化立即停止而无法结实,最终导致严重减产。大多数冬小麦品种冬季抗寒能力强,从返青期开始恢复生长后,其抗寒力将大幅度下降,不能适应气温突降的环境。拔节期低温是影响冬小麦产量的关键时期,冬小麦拔节前20 d至拔节后40 d均会有低温冻害发生,其中以拔节前20 d至拔节后15 d发生频率为最高。另外,研究发现重霜冻害主要发生在冬小麦拔节5~15 d。而拔节后的冬小麦雌雄蕊分化期抗寒能力较低,若此时出现低温,也会给小麦造成严重危害。
目前,国内外已有许多关于低温胁迫下小麦生理生化的研究,其对小麦的引种、育种以及揭示低温对冬小麦的影响具有重要意义,但上述研究并未对冬小麦拔节关键期进行细化。因此,本研究在全球气候变暖,日最低气温及冬小麦受胁迫生育期不断变化的背景下,通过人工控制环境试验,设计不同水平的日最低温度,以冬小麦优良品种“郑麦366”为试验材料,探讨不同拔节期进程下冬小麦光合特性及干物质转运系统对低温的响应机理,以期为冬小麦抗寒品种的选育以及低温防御提供指导。
1 材料和方法
1.1 试验设计
以河南省主要推广栽培品种郑麦366(半冬性)为试验材料,采用盆栽试验的方法,于2018年在河南省郑州市农业气象试验站进行。试验用土取自大田冬小麦-夏玉米轮作区耕层土,土壤为砂壤土,有机质含量为14.10 g/kg,全氮含量为0.99 g/kg,碱解氮含量为89.20 mg/kg,速效磷含量为29.90 mg/kg,速效钾含量为90.60 mg/kg,pH值为7.8。田间持水量为24.63%。砂壤土自然风干后与肥料充分混匀后装入高28 cm、直径30 cm的聚乙烯塑料桶,每桶装过筛干土7.0 kg,桶底用钻头打洞以利于透气。盆栽,埋于试验大田且保持盆内土壤与大田齐平。11月15日进行播种,于三叶期定苗,每盆留苗30株。试验设置冬小麦拔节初期(拔节5 d)、拔节中期(拔节10 d)、拔节后期(拔节15 d)3个处理时间和日其对应最低气温-3 ℃、-5 ℃和-7 ℃共3个低温处理,模拟自然情况温度下降曲线,逐小时设置温度,处理时间长度均为1个晚上。低温处理结束进行取样后,将被处理的冬小麦移至室外并埋于试验田中,自然状态下直到成熟,留作后期观测冬小麦的存活及再生长情况。
试验小区从东至西两列排开,随冬小麦拔节期不同阶段设置拔节后未遭遇低温(CK)、T1、T2、T3共4个处理,其中T1、T2、T3分别于拔节初期(拔节5 d)、拔节中期(拔节10 d)、拔节后期(拔节15 d)遭遇其对应低温(-3 ℃、-5 ℃和-7 ℃),每个处理设置3个重复。每个小区长1.80 m,宽0.65 m,南北排列。
1.2 测定项目与方法
1.2.1 冬小麦叶片光合作用参数的测定 采用LI-6400便携式光合作用测定系统(LI-COR Biosciences Inc.,USA)测定冬小麦叶片光合特性及气体交换参数。在低温处理完成后的冬小麦各个生育时期(拔节期、孕穗期、扬花期、灌浆期)选取功能叶进行测定,测定内容为净光合 速 率(Net photosynthetic rate,Pn)、 气孔 导度(Stomatal conductance,Gs)、 胞 间CO2浓 度(Intercellular CO2concentration,Ci) 和 蒸 腾 速 率(Transpiration rate,Tr)。
1.2.2 冬小麦产量结构及干物质转运量、转运效率的测定 低温处理后的冬小麦在收获后进行考种。测定项目包括每盆穗数、茎秆整质量、千粒质量、每盆产量。于冬小麦各生育时期取3株完整植株,用自来水洗去根上泥土及叶茎上的杂质,晾干后按根、茎、叶、鞘分割为4个部分,各部分放入烘箱于105 ℃下杀青1 h,75 ℃烘干12 h后称质量并记录,此后每小时称质量1次,当样本前后2次质量差值低于5%时停止烘干,测定冬小麦各营养器官的干质量。冬小麦营养器官干物质转运量为其各生育时期所测干质量的最大值与成熟期干质量的差值,转运效率为转运量占所测干质量最大值的百分比。
1.3 数据处理与分析
试验数据用SPSS软件进行分析,用Duncan's多因素分析检查数据间的显著性,并用Microsoft Excel 2019软件完成数据处理与作图。
2 结果与分析
2.1 拔节期低温对冬小麦叶片光净光合速率的影响
如图1所示,随着生育时期的推进,除-7 ℃处理外,其他低温处理下冬小麦叶片净光合速率变化均呈单峰趋势,即在扬花期达到最大值后逐渐回落。此外,经低温处理的冬小麦各生育时期叶片净光合速率均低于对照处理,且降幅随处理温度降低而加剧,于-7 ℃处理下降至最低,表明低温胁迫越严重,冬小麦叶片净光合速率下降幅度越大。拔节期遭遇低温可使冬小麦叶片净光合速率下降,各拔节期进程中冬小麦净光合速率下降幅度大小依次为拔节中期>拔节后期>拔节初期。
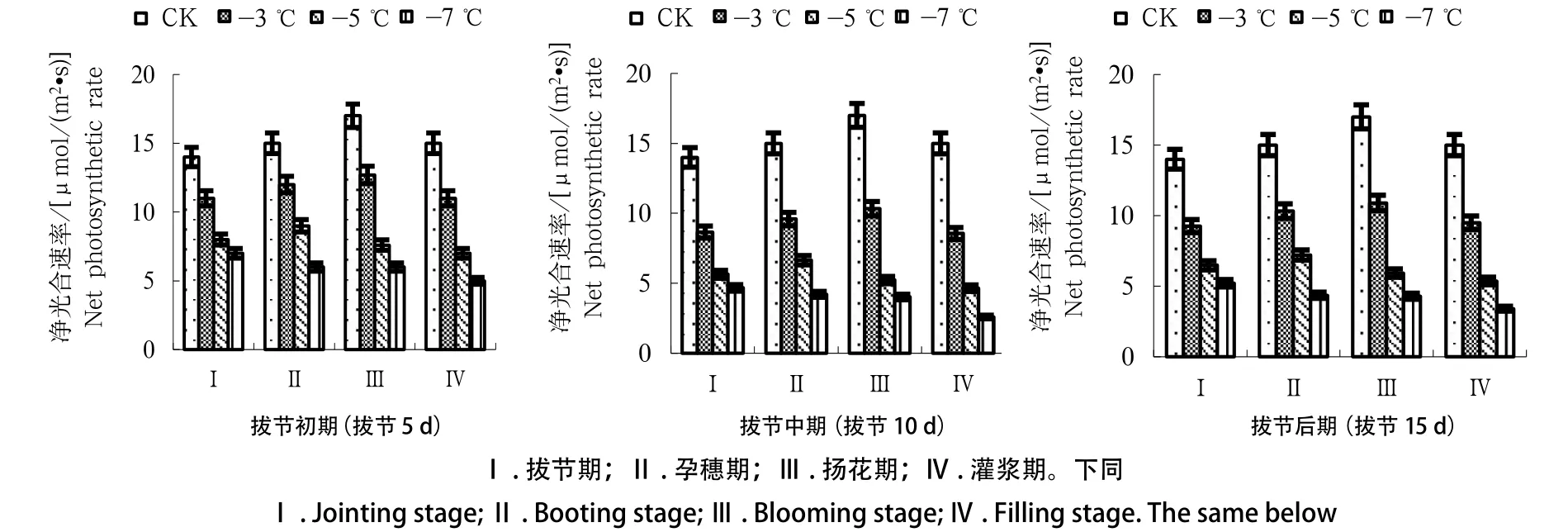
图1 不同拔节进程低温对冬小麦净光合速率的影响Fig.1 Effects of different jointing processes on net photosynthetic rate of winter wheat
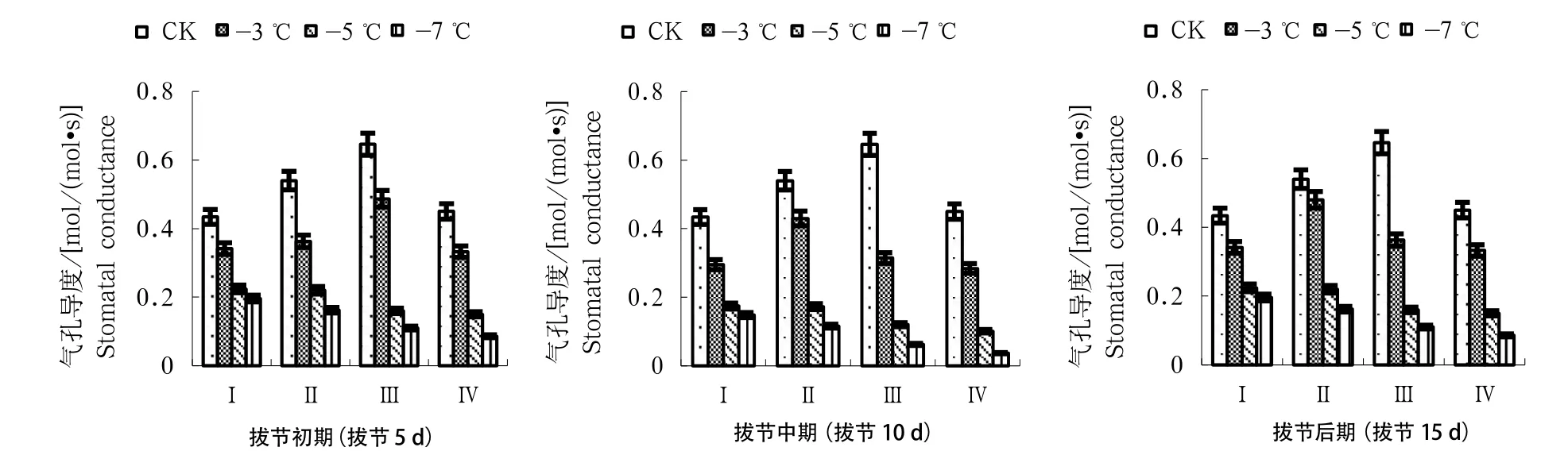
图2 不同拔节进程低温对冬小麦叶片气孔导度的影响Fig.2 Effects of low temperature at different jointing stages on stomatal conductance of winter wheat leaves
2.2 拔节期低温对冬小麦叶片气孔导度的影响
如图2所示,对照处理不同拔节进程中冬小麦叶片气孔导度均在扬花期达到最大值后于灌浆期开始回落。于拔节初期进行-3 ℃处理,冬小麦叶片气孔导度上升至扬花期后下降。当进行-5 ℃处理时,拔节中期各生育阶段冬小麦叶片气孔导度与对照处理相比降幅最大。当进行-7 ℃处理时,拔节中期各生育阶段冬小麦叶片气孔导度呈直线下降趋势,在不同拔节进程中与对照处理相比降幅最大。拔节期遭遇低温导致冬小麦叶片气孔导度不同程度下降,各拔节时期进程中冬小麦叶片气孔导度下降幅度大小依次为拔节中期>拔节后期>拔节初期。
2.3 拔节期低温对冬小麦叶片胞间CO2浓度的影响
如图3所示,不同生育时期中,冬小麦叶片胞间CO2浓度在拔节期达到最大,其他生育时期明显下降,虽于灌浆期略有回升但各生育时期之间差异不显著。在拔节期时,3种低温胁迫均导致叶片胞间CO2浓度不同程度升高。在不同拔节进程中进行-3 ℃处理时,各生育期冬小麦叶片胞间CO2浓度变化趋势与对照处理一致。在不同拔节进程中进行-5 ℃处理时,各生育期冬小麦叶片胞间CO2浓度较-3 ℃处理下升高,而在不同拔节进程中进行-7 ℃处理时叶片胞间CO2浓度进一步升高。拔节期遭遇低温可导致冬小麦叶片胞间CO2浓度上升,各拔节时期进程中CO2浓度上升幅度大小依次为拔节中期>拔节后期>拔节初期。

图3 不同拔节进程低温对冬小麦叶片胞间CO2浓度的影响Fig.3 Effects of different jointing processes on intercellular CO2 concentration in winter wheat leaves
2.4 拔节期低温对冬小麦叶片蒸腾速率的影响
如图4所示,拔节期遭遇低温后,冬小麦叶片蒸腾速率在扬花期达到最大,其他生育时期明显下降。在不同拔节进程中进行-3 ℃处理时,各生育时期冬小麦叶片蒸腾速率变化趋势与对照处理一致,且在同一生育时期内均低于较对照处理。在不同拔节进程中进行-5 ℃和-7 ℃处理时,冬小麦叶片蒸腾速率随着处理温度的降低进一步下降,且温度越低冬小麦叶片蒸腾速率越低。拔节期遭遇低温可使冬小麦叶片蒸腾速率下降,各拔节期进程中叶片蒸腾速率下降幅度大小依次为拔节中期>拔节后期>拔节初期。

图4 不同拔节进程低温对冬小麦蒸腾速率的影响Fig.4 Effects of different jointing processes on transpiration rate of winter wheat
2.5 拔节期低温对冬小麦产量结构的影响
由表1可知,冬小麦受不同程度低温胁迫,对各产量结构因子影响程度不同。随着胁迫温度的降低,冬小麦每盆穗数、茎秆整质量、千粒质量、每盆产量下降。在不同拔节进程中进行-7 ℃处理冬小麦各产量结构因子下降幅最大,其中拔节初期(拔节5 d)、拔节中期(拔节10 d)、拔节后期(拔节15 d)冬小麦每盆穗数分别较对照处理下降了51.41%、79.06%、48.43%(P<0.05),茎秆整质量分别较对照处理下降了22.72%、36.09%、33.97%(P<0.05),千粒质量分别较对照处理下降了15.48%、16.94%、5.89%(P<0.05),每盆产量分别较对照处理下降了54.33%、59.83%、58.66%(P<0.05)。

表1 不同拔节期进程低温对冬小麦产量结构的影响Tab.1 Effects of low temperature in different jointing stages on yield structure of winter wheat
2.6 拔节期低温对冬小麦干物质转运量和转运效率的影响
由表2可知,冬小麦营养器官的干物质转运量随着处理温度的降低而下降。冬小麦干物质转运效率表现为叶片、鞘、根随着温度的降低而下降,但茎的转运效率却随温度下降而上升。相同温度下拔节中期各营养器官的干物质转运量及转运效率最低,拔节后期次之。
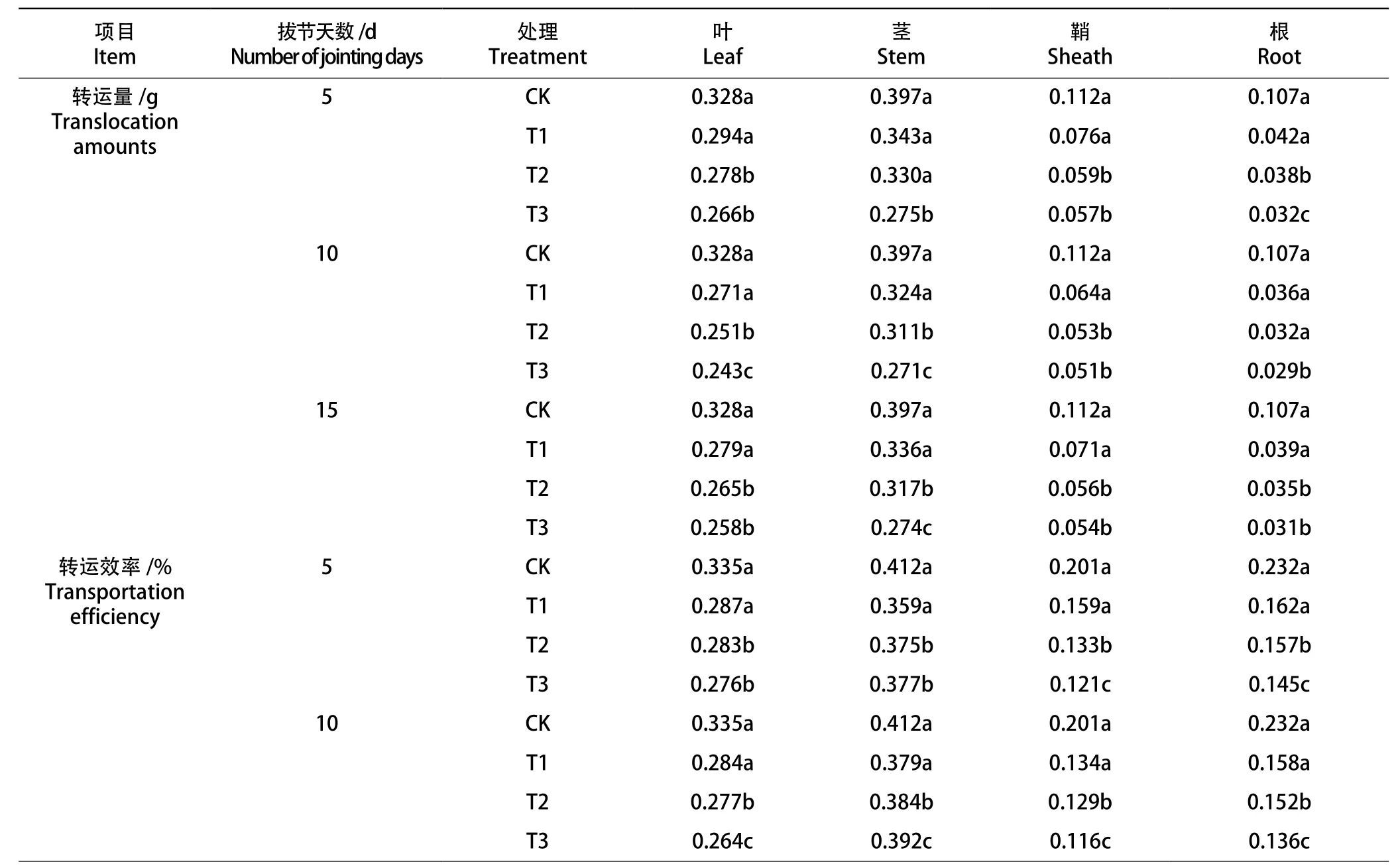
表2 不同拔节期进程低温对冬小麦干物质转运量和转运效率的影响Tab.2 Effects of low temperature in different jointing stages on dry matter translocation and transport efficiency of winter wheat
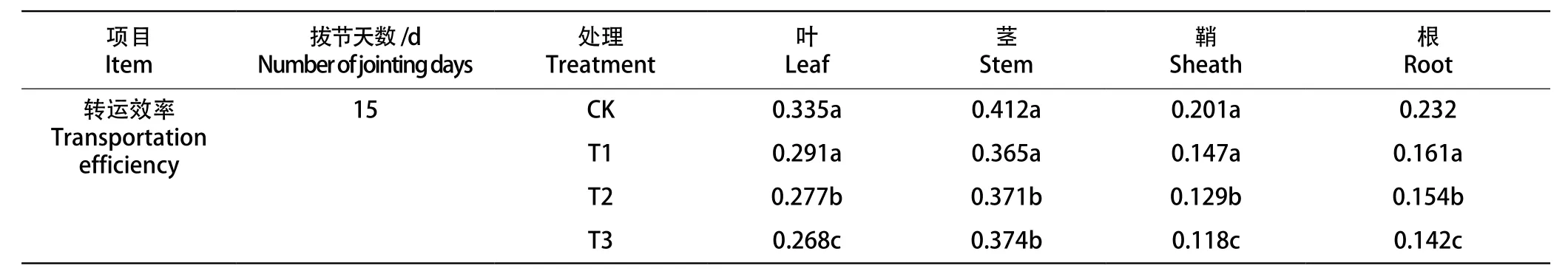
续表2 不同拔节期进程低温对冬小麦干物质转运量和转运效率的影响Tab.2(Continued) Effects of low temperature in different jointing stages on dry matter translocation and transport efficiency of winter wheat
3 结论与讨论
植物的光合器官对温度极为敏感,温度的变化会直接导致光合器官的性能出现变化。本试验中,在拔节进程中的各生育时期遭遇低温均可导致冬小麦叶片光合性能下降,这与前人研究结果一致。冬小麦叶片净光合速率降低的程度不仅取决于低温胁迫程度,也与冬小麦所处的拔节进程密切相关。相同条件下,拔节中期冬小麦叶片光合性能对低温更为敏感,说明拔节中期发生低温灾害更易使冬小麦受灾。光合作用是小麦干物质生产的基础,低温胁迫下小麦叶片的气孔收缩抑制了小麦叶片的光合作用,导致干物质合成转运受阻,最终造成小麦产量降低。
本试验结果表明,低温胁迫下冬小麦叶片的蒸腾速率在拔节与灌浆期下降最明显。蒸腾拉力降低会导致小麦植株内矿物质营养和水分的运输受阻,从而降低了光合底物的传导能力。气孔是水分和CO2进出植株的窗口,气孔导度也会随蒸腾拉力降低而下降。此外,当小麦叶片的净光合速率下降时,若胞间CO2浓度和气孔导度也同时下降,则净光合速率的降低主要是气孔限制引起;若胞间CO2浓度上升,光合速率的降低主要的限制因素为非气孔因素。本试验中,冬小麦胞间CO2浓度除在拔节期明显升高外,其他生育时期均呈现波动变化,说明光合作用的主要限制因素在气孔因素和非气孔因素之间来回转变,也表明气孔导度和蒸腾速率的降低在一定程度上降低了环境对冬小麦的伤害。因此,在不同拔节周期低温胁迫下,冬小麦光合系统的保护机制存在差异。
低温对冬小麦产量结构的影响主要表现在穗数减少、茎秆整质量下降、千粒质量下降等。低温胁迫下,冬小麦产量主要从茎鞘中储存的光合产物中获取,冬小麦茎秆是营养器官向穗部输送水分、矿质元素及干物质的通道,也可以看作“流(同化物从源到库的通道)”的指标,其强弱与冬小麦千粒质量及穗数密切相关。本试验中,光合产物转运量及转运速率随温度降低而下降,最终导致冬小麦干物质积累减少,使小麦产量下降。
综上所述,冬小麦在不同拔节进程中遭遇低温胁迫时,其生理功能及光合能力均受到严重影响,但存在差异,以拔节中期遭遇低温时的影响为最严重,这为进一步研究低温对冬小麦的胁迫机制奠定了基础。
