
重金属镍胁迫对向日葵幼苗生理生化特性的影响
2019-09-10 07:22:44曾小飚唐健民朱成豪邹蓉,史艳财韦霄蔡丽华
广西植物 2019年12期
关键词:生长
曾小飚 唐健民 朱成豪 邹蓉, 史艳财 韦霄 蔡丽华


摘要: 为揭示向日葵对镍胁迫的响应机制,该文以向日葵幼苗为材料,采用营养液培养法探索了重金属镍胁迫对其生理生化指标的影响。结果表明:低浓度的镍胁迫(≤10 mg·L-1)有利于幼苗的生长,高浓度的镍胁迫(≥50 mg·L-1)对幼苗具有明显的抑制作用。随着镍胁迫浓度的逐渐增加,向日葵幼苗各项生理生化指标的变化较大,其中叶绿素、可溶性糖(SS)、可溶性蛋白(SP)、过氧化物酶(POD)活性、游离脯氨酸(Pro)的含量均在1~10 mg·L-1时呈现上升趋势,在50~100 mg·L-1时呈下降的趋势,丙二醛(MDA)含量则呈现持续上升趋势。这表明向日葵幼苗对低浓度(1~10 mg·L-1)镍胁迫能够通过自身调节,增加POD、Pro和MDA等物质的含量来提高对生态环境的抗逆能力,说明向日葵幼苗对重金属镍具有一定抗性;而高浓度(50~100 mg·L-1)镍胁迫会破坏其自身防御系统,从而影响幼苗的生长发育。
关键词: 向日葵幼苗, 镍胁迫, 生长, 生理生化特性, 响应机制
中图分类号: Q945.78文献标识码: A文章编号: 1000-3142(2019)12-1702-08
Abstract: In order to reveal the response mechanism of Helianthus annuus nickel stress, nutrient solution culture experiments were carried out to probe the affection of heavy metal nickel on physiological and biochemical characteristics by using Helianthus annuus seedlings as materials. The results showed that low nickel concentration (≤10 mg·L-1) was beneficial to the seedlings growth, and high nickel concentration (≥50 mg·L-1) had obvious inhibitory effects on seedlings. With the increasing of nickel concentration, the physiological and biochemical indexes changed greatly, in which chlorophyll, soluble sugar (SS), soluble protein (SP), peroxidase (POD) activity, free proline (Pro), all their contents increased when the nickel concentration was 1-10 mg·L-1, and decreased when the nickel concentration was 50-100 mg·L-1; but the content of malondialdehyde (MDA) continued to rise. This indicates that the seedlings can used autoregulation to improve resistibility by increasing POD, Pro and MDA in low nickel concentration (1-10 mg·L-1), and that seedlings have certain resistance to heavy metal nickel, while high concentration (50-100 mg·L-1) nickel stress will destroy its own defense system so as to affect the growth and development of seedlings.
Key words: Helianthus annuus seedlings, nickel stress, growth, physiological and biochemical characteristics, response mechanism
土壤是人類生存的基础,随着我国社会的进步和工业的不断发展,土壤生态环境状况十分严峻,土壤重金属污染问题日趋严重。重金属借助风力、水力以及化学迁移等方式进入到土壤中(白玉杰等,2018),在土壤中富集无法被微生物降解,造成植物吸附后导致生理功能紊乱,影响植物生长发育和生存环境的变化。由于重金属污染的长期性和不可逆性(吕学研等,2014),研究重金属污染的防治和修复显得十分迫切;镍是土壤中广泛分布的金属污染物之一,根据《全国土壤污染状况调查公报》显示,我国 Ni 污染超标率为 4.8%,镍的污染研究已成为人们关注的热点。随着对重金属镍的研究不断开展,有大量的文献报道,主要研究集中在对玉米(王丽娜,2014)和板蓝根(王兵和马芳,2014)等作物的生理生化影响,但关于观赏性的重金属镍富集植物的研究报道较少,研究观赏性植物对重金属镍的富集性种植和土壤修复具有重要意义。
向日葵(Helianthus annuus)别名朝阳花、望日莲、太阳花,一年生草本,是菊科向日葵属的植物。高1~3.5 m,花期7—9月,花色有金黄色、红色及复色等,具有非常高的观赏价值,广泛用于公园花境营造、盆花、染色花及庭院美化等领域(徐惠风等,2003)。向日葵茎干直、根系较发达、喜光照、生长周期短、生物量大、对重金属有较强的富集性(郭平等,2007)。
该研究利用镍元素在不同浓度下所配成的混合营养液对向日葵进行培养,研究不同浓度镍胁迫对向日葵生长发育和生理生化指标的影响;从形态指标和生理生化指标两个方面分析镍胁迫对其的影响,旨在探索镍胁迫对幼苗生长的影响程度和向日葵幼苗对镍胁迫的响应机制;为确定镍元素对观赏向日葵生长发育的影响和镍污染环境修复提供一定的科学依据。
1材料与方法
1.1 材料
向日葵种子为金星1号,购于百色城西蔬菜种子店。Hoagland营养液的配方:硝酸钙945 mg·L-1;硝酸钾607 mg·L-1;磷酸铵115 mg·L-1;硫酸镁493 mg·L-1;铁盐溶液2.5 mL·L-1;微量元素5 mL·L-1;pH=6.0。
1.2 方法
1.2.1 种子萌发及幼苗培育精选饱满的向日葵种子,用1/1 000的高锰酸钾溶液进行表面消毒30 min,蒸馏水清洗3次,置于垫有纱布的平底托盘中,在(25±1)℃下萌发。待四分之三的种子萌芽后,将其植入装有洁净细砂的塑料杯中,用新配置的Hoagland培養液浇灌,自然条件下培养一段时间后,挑选长势一致(高15 cm)的幼苗分为6组,每组30棵,供镍胁迫实验用。
1.2.2 重金属镍胁迫分别用浓度为0、1、5、10、50、100 mg·L-1的镍胁迫溶液(分别取一定质量的NiSO4· 6H2O溶于1/2Hoagland培养液中)浇灌向日葵幼苗,于阳光充足的条件下培养,并统一在植株高于土壤表面约2 cm处用记号笔做好标记,便于以后从此标记处测量株高,胁迫12 d后对向日葵幼苗做各项生理生化指标测定。
1.3 向日葵幼苗叶片生理生化指标的测定
分别对每个胁迫浓度的向日葵幼苗叶片进行采集、提取测定;相同指标所取叶片的形态学位置相同。叶绿素含量测定采用80%丙酮萃取法(赵世杰等,1998);丙二醛(MDA)与可溶性糖(soluble sugar, SS)含量测定分别采用硫代巴吡妥酸(TBA)法和蒽酮法(李合生,2000);过氧化物酶(POD)活性测定采用愈创木酚法(张治安等,2004);可溶性蛋白(soluble protein, SP)含量测定采用考马斯亮蓝染色法;游离脯氨酸(Pro)含量测定采用酸性茚三酮法等(邹琦等,2000)。以上实验做3次重复,株高用游标卡尺测量。
1.4 数据处理分析
利用 Excel 和 Origin 2015 软件对所得数据进行处理分析及制图。
2结果与分析
2.1 镍胁迫对向日葵幼苗株高及形态的影响
株高作为衡量植物生长特征的重要指标之一,株高的变化反应植物的生长状态。由图1可知,在低浓度区间,1~10 mg·L-1向日葵幼苗株高逐渐增高,叶片直立绿色,说明低浓度的镍胁迫有利于植株的生长;随着浓度升高,50 mg·L-1株高的增幅显著低于对照组,叶片逐渐出现弯曲、萎蔫的现象;在100 mg·L-1镍胁迫下,向日葵幼苗叶片已发黄、出现斑点,并伴随着萎蔫现象,说明此阶段向日葵幼苗叶片的生长受到了严重抑制。
2.2 镍胁迫对向日葵幼苗叶绿素含量的影响
植物叶绿素主要包括叶绿素a与叶绿素b,植物光合作用离不开叶绿素,光合作用的强弱直接影响植物体内有机物的合成。叶绿素含量减少,叶片失绿,重者导致死亡。从图2可以看出,在1~5 mg·L-1浓度胁迫下,镍浓度升高,叶绿素a含量随之增加,但上升幅度不大,分别增加了2.4%、8.1%;但浓度10 mg·L-1胁迫下,增加了17.8%,上升显著;当镍胁迫浓度在50~100 mg·L-1时,叶绿素a含量则下降明显,分别降低了5.9%、19.9%。最高胁迫浓度导致叶绿素a减少的量远高于最低胁迫浓度促进增长的量。可见,1~10 mg·L-1镍浓度促进叶绿素a的合成,增强光合作用效率,50~100 mg·L-1镍浓度对向日葵幼苗毒害较深。而叶绿素b含量分别比对照组增加了10.4%、11.6%、15.9%,呈持续上升趋势。叶绿素b含量达到峰值之后,开始大幅度下降,受镍胁迫浓度影响较大,相对于只用营养液培养组(比对照)降低了19.2%、42.3%。叶绿素b在重金属镍的影响下,其含量下降幅度远远大于上升幅度。由此可见,在向日葵幼苗生长发育过程中,低浓度镍对叶绿素b的合成有促进作用,高浓度下光合作用效率降低。综合来看,叶绿素a与叶绿素b对镍毒害的敏感程度有很大差别,镍对叶绿素b的作用力更大,毒害更深。
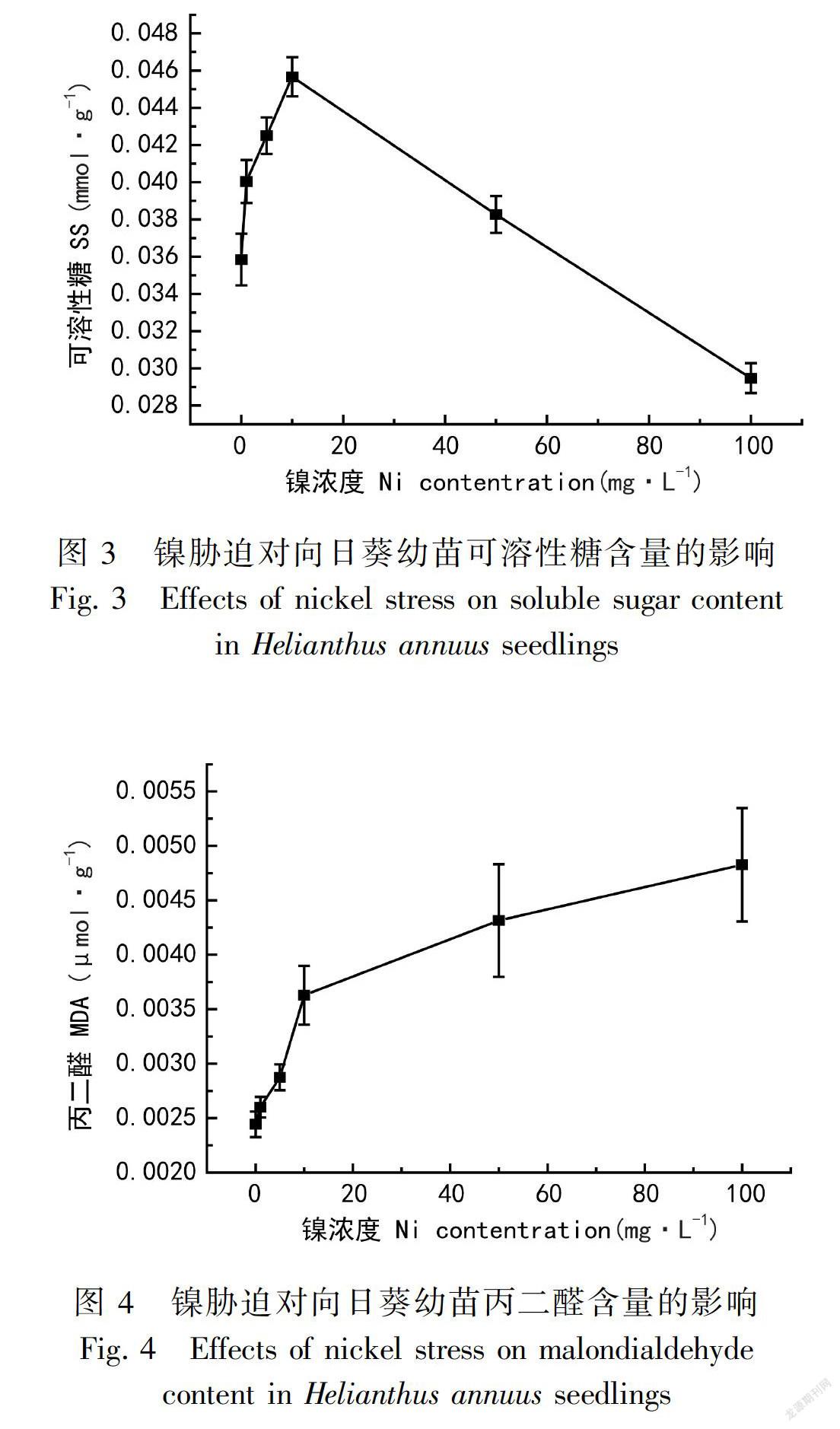
2.3 镍胁迫对向日葵可溶性糖含量的影响
可溶性糖是植物体光合作用的产物,以单糖为主,为生命活动提供能量,逆环境下,这类小分子化合物可以作为渗透调节剂维持渗透平衡。其含量增加可以降低重金属毒害机体,提高植物体的抗逆能力。
由图3可知,当镍胁迫浓度≤10 mg·L-1时,可溶性糖在向日葵叶片中的含量明显增加。随着不同浓度胁迫溶液加入12 d后,可溶性糖迅速增加14.2%,之后逐渐增加了18.2%、29.3%,可溶性糖最大增加量是10 mg·L-1时的2.07倍;当胁迫浓度增加到50 mg·L-1时,可溶性糖含量锐减,增加量达到最低。1~10 mg·L-1镍处理后,可溶性糖含量持续增加;50 ~100 mg·L-1镍处理后,含量明显下降,经100 mg·L-1镍处理后与对照组相比,可溶性糖含量减少了16.1%。可以看出向日葵叶片中可溶性糖含量的变化趋势呈先上升后下降。1~10 mg·L-1浓度的镍胁迫对向日葵影响不大,可溶性糖的增加可以起到渗透调节的作用,减轻重金属带来的伤害;高浓度时,植株已经受到较深的毒害,可溶性糖的含量明显降低。
2.4 镍胁迫对向日葵丙二醛含量的影响
植物在逆环境下受伤害,其组织或器官膜脂质发生过氧化反应产生丙二醛(MDA),使质膜平衡体系受损。因此MDA含量的多少可以反映出植物体受伤程度。
从图4可以看出,MDA的含量随着镍胁迫浓度的增加小幅度上升且都高于对照组。在浓度为1、5、10、50、100 mg·L-1的镍胁迫下,MDA含量一直处于上升趋势,增加率为0.4%、11.8%、44.8%、62.3%、71.4%。在低浓度镍胁迫下,向日葵幼苗MDA含量变化较小,当浓度超出5 mg·L-1时,含量显著地增加;到10 mg·L-1时,丙二醛含量相对5 mg·L-1时高出33%,增加非常显著。说明在低浓度(≤5 mg·L-1)镍胁迫的逆环境下,镍胁迫对细胞膜的伤害不大,而随着镍浓度的逐渐增大,促使丙二醛含量增多,反映向日葵幼苗膜质损伤的程度逐渐加深。
2.5 镍胁迫对向日葵可溶性蛋白含量的影响
生物体生命活动离不开蛋白质,植物体内的可溶性蛋白几乎都是参与各种代谢的酶类,催化各种化学反应。在重金属胁迫下,其含量变化是反应植物抗逆性的重要指标。
在重金属镍1~100 mg·L-1的胁迫下,向日葵叶片中的可溶性蛋白含量变化(图5),经1~50 mg·L-1的镍处理,向日葵叶片中可溶性蛋白含量均高于对照组,分别高出12.0%、15.9%、20.7%、7.6%,100 mg·L-1的镍处理后,比对照组低了3.8%。这说明低浓度的镍处理刺激可溶性蛋白的合成,以减少镍胁迫带来的毒害;经高浓度的镍处理后细胞代谢活动减慢,可溶性蛋白的合成受到抑制,植物的生长发育受到严重影响。从整体看来,镍胁迫明显抑制可溶性蛋白的合成。
2.6 镍胁迫对向日葵过氧化物酶活性的影响
过氧化物酶(POD)广泛存在于植物体内,能有效清除过量的活性氧,提高植株的抗逆性,是植物重要的保护酶之一。过氧化物酶活性在植物生长发育过程中随胁迫浓度的增加不断发生变化,在抵制外界逆境毒害过程中扮演着重要的角色。
结果图6显示,镍胁迫12 d后,向日葵叶片的POD活性先上升,至最高点后持续下降。经1 ~10 mg·L-1 镍处理,过氧化物酶活性持续上升,实验组超过对照组2.1%、10.6%、16.1%,活性最大出现在镍胁迫浓度为10 mg·L-1。这说明低浓度镍有增强植物叶片中POD活性的作用。50 mg·L-1的镍处理,会导致过氧化物酶活性显著下降,相对10 mg·L-1而言,降低了14.2%。虽然活性明显降低,但仍超出对照组1.8%。在100 mg·L-1的胁迫下,过氧化物酶活性低于对照组,说明镍胁迫浓度过高,会导致向日葵叶片中的过氧化物酶活性明显降低。
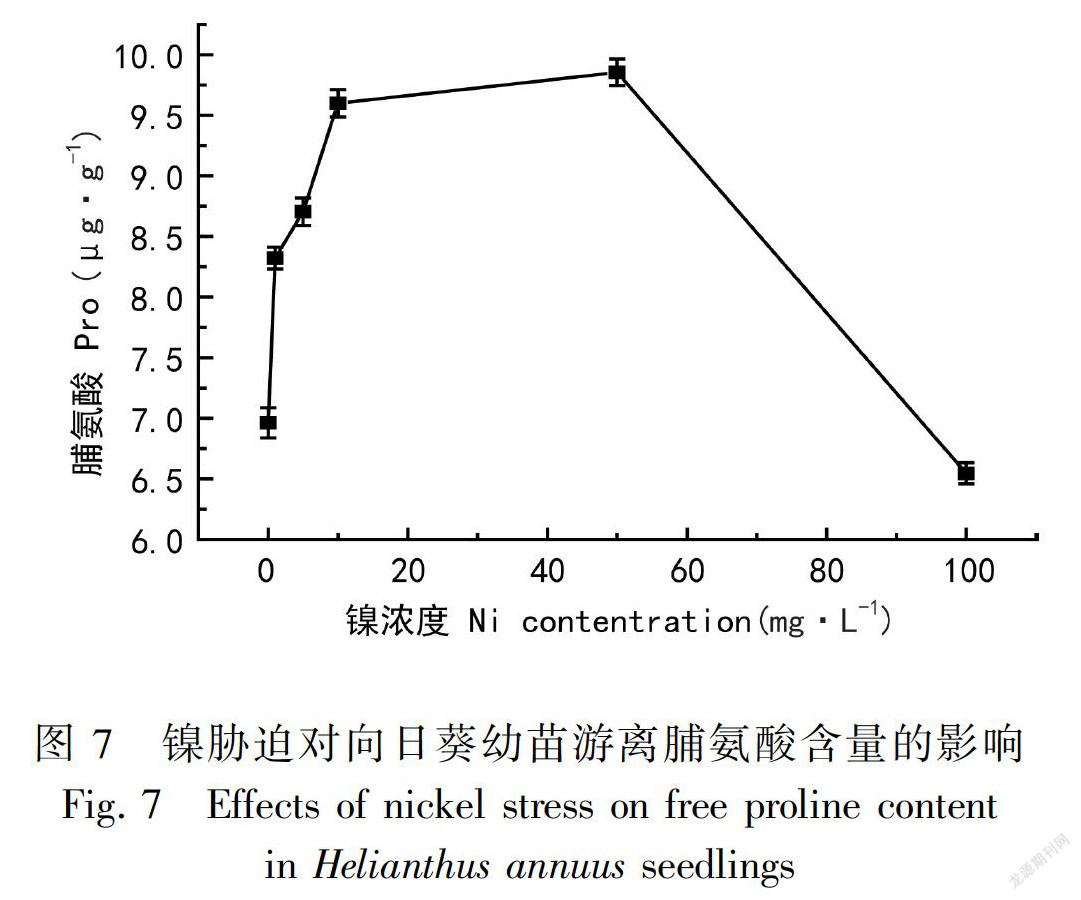
2.7 镍胁迫对向日葵游离脯氨酸含量的影响
游离脯氨酸(Pro)具有较强的溶解性,可以起到防止植物组织脱水与减小渗透压的作用。游离脯氨酸在细胞中大量的积累,作为自身的防脱水剂和渗透压剂,对于保证组织的持水力与降低细胞内溶质的渗透势有重大意义。
由图7可知,向日葵在镍胁迫下,向日葵叶片中的游离脯氨酸含量随着镍胁迫浓度的增加呈先上升后降低的趋势。当胁迫浓度为1、5 mg·L-1时,游离脯氨酸含量的增加值比较接近,分别增加了19.6%、22.1%。当胁迫浓度为10、50 mg·L-1时,比对照高出了38.0%、41.7%,两组增加量明显比前面的大,游离脯氨酸含量最高是在镍胁迫浓度为50 mg·L-1时。经100 mg·L-1的镍胁迫,游离脯氨酸含量骤降,比对照组降低了5.9%。由此可说明,低浓度的镍处理,向日葵叶片中脯氨酸仍不断增加,起到保持细胞渗透势的作用,是适应逆环境的一种表现;100 mg·L-1时向日葵受镍毒害严重,游离脯氨酸的积累减少,导致向日葵的渗透平衡破坏。
3讨论
植物在多种因素的共同作用下表现出抗逆性,其生理生化指标是研究植物抗逆能力的基础。镍具有潜在性,无论是低浓度还是高浓度都会对植物的外观形状、生物量和生理生化过程具有一定的影响(刘金华等,2012;赵娜和周米平,2011;鲁艳等,2012)。
在逆境中,植物的POD、MDA会发生协调性变化。过氧化物酶在逆境中被激活,有研究表明,过氧化物酶活性随镍胁迫浓度的增加先升后降(赵娜,2011)。本研究结果表明,有镍溶液加入时,POD活性几乎无变化,5 mg·L-1时活性明显上升。当POD活性到达一定程度后就开始下降,且下降幅度大,直到低于对照组。这表明向日葵幼苗在一定镍胁迫条件下,加强了POD活性并激发植株自身防御机制,以清除镍胁迫带来的膜脂过氧化伤害。胁迫浓度过高,过氧化物酶则受到破坏,活性降低。活性降低可能是由于胁迫致使向日葵幼苗细胞膜系统受损,破坏POD分子结构,使植物不能耐受高浓度胁迫。研究结果表明MDA含量随镍浓度的增加而增加。在高浓度的镍胁迫下,活性自由基大量生成,机体的清除能力有限,MDA含量逐漸增加,导致向日葵幼苗膜脂过氧化作用增强,质膜透过性增加(郑爱珍,2005),影响细胞正常代谢,POD活性降低,说明POD这类保护酶对植物膜系统的保护具有限制性。
本研究结果表明,镍在低浓度时,可以促进向日葵叶绿素合成。高浓度胁迫严重破坏叶绿素合成机制,使机体无法抵制镍毒害。这说明向日葵对低浓度镍胁迫有较强的抗性,能正常进行光合作用,叶绿体是较敏感的细胞器之一,在高浓度胁迫下,叶绿体结构遭到破坏,使叶绿素含量下降,光合作用减弱,影响植物体的生长发育。本研究发现,镍对叶绿素b的毒害作用比叶绿素a强,50 mg·L-1的胁迫下,叶绿素b含量下降幅度更大,大概是两者结构存在差异的原因。在100 ml·L-1的镍胁迫下,向日葵叶片失绿,出现斑点并伴有灼烧现象,表明此时向日葵幼苗耐受镍毒害达到最大限度,自身防御系统严重破坏。
可溶性糖是光合作用的产物,既是植物体的能量来源,也是渗透调节不可或缺的部分。在黄栌和紫荆的研究中,随胁迫浓度增加,可溶性糖含量呈上升趋势(李国雷等,2004)。有研究表明低浓度胁迫下,可溶性糖含量随镍胁迫浓度增加逐渐上升,可溶性糖的积累可能是机体在逆环境下的一种应急机制,随着胁迫浓度的持续增加,可溶性糖含量开始降低(常云霞等,2018)。这说明向日葵幼苗在高浓度金属胁迫下,叶绿素合成途径受阻,光合作用效率下降,呼吸作用加强,有机物合成减少,运输途径受到阻碍,可溶性糖含量逐渐减少,在植物体内的渗透调节能力受到限制,影响植株的生长发育。有研究表明向日葵幼苗在镍胁迫下,可溶性蛋白的含量先是逐渐上升,随后缓慢下降(张牡丹等,2018)。可溶性蛋白合成增强并参与渗透调节,提高其抗逆性,保护自身机制免受伤害;当胁迫浓度超出一定范围,蛋白质合成受阻,植物的这种自我调节机制受到破坏,导致向日葵幼苗生命活动受到影响。
游离脯氨酸是植物在逆环境中重要渗透调节剂之一,可以起到保持机体渗透平衡的作用。研究表明随着外界胁迫浓度的增加,植物体内的游离脯氨酸含量升高(张云起等,2003;杨树军等,2008;武春霞等,2008)。本研究结果表明,在镍浓度≤50 mg·L-1时,游离脯氨酸在向日葵叶片中含量与镍浓度呈正相关;当镍浓度达到100 mg·L-1时,游离脯氨酸含量急剧下降,低于对照组。这说明一定浓度的镍胁迫溶液会使植物体内游离脯氨酸含量不断增加,维持体内渗透势平衡,是植物自身采取的一种保护措施;当镍浓度超过一定范围时则会导致脯氨酸含量下降,破坏了植物的自我保护。可见,游离脯氨酸在机体内的调节作用受镍胁迫浓度影响较大。
4结论
通过营养液培养法研究不同浓度的镍胁迫向日葵幼苗后叶片各项生理生化指标变化规律,得出以下结论:低浓度的镍胁迫对向日葵幼苗叶绿素、可溶性糖、可溶性蛋白、过氧化物酶、游离脯氨酸的合成影响不大,植株可以通过自身调节,抵制镍毒害,提高抗逆能力,各生理生化指标不仅表明向日葵幼苗对重金属镍有一定的抗性;也侧面反应镍和向日葵的生长发育状态有一定的相关性,表明向日葵作为观赏性植物可在轻度镍污染的土壤中进行生态修复栽培。
参考文献:
BAI YJ, SHEN GX, CHEN XH, et al., 2018. Accumulation and transport of nickel in three vegetable crops and their edible safety [J]. J Agro-Environ Sci, 37(8): 1619-1625. [白玉杰, 沈根祥, 陈小华, 等, 2018. 三种蔬菜对镍累积转运规律及食用安全研究 [J]. 农业环境科学学报, 37(8):1619-1625.]
CHANG YX, LI ZL, LI FR, 2018. Effect of IAA on the osmotic adjustment substance and antioxidant properties in Arabidopsis thaliana seedling under Cd2+ stress [J].J Zhoukou Norm Univ, 35(5):79-82. [常云霞, 李姿琳, 李芙蓉, 2018. IAA 對Cd2+胁迫下拟南芥渗透调节物质及抗氧化特性的影响 [J]. 周口师范学院学报, 35(5):79-82.]
GOMES-JUNIOR RA, MOIDES CA, DELITEFS, et al., 2006. Niekel elicits a fast antioxidant response in Coffea arabiea cells [J]. Plant Physiol Biochem, 44:420-429.
GUO P, LIU C, ZHANG HB, et al., 2007. Study on the enrichment ability and tolerance of Pb and Cu in sunflower seedlings [J]. J Soil Water Conserv, 21(6): 92-95, 113. [郭平, 刘畅, 张海博, 等, 2007. 向日葵幼苗对Pb、Cu富集能力与耐受性的研究 [J]. 水土保持学报, 21(6):92-95,113.]
LI GL, SUN MG, XIA Y, et al., 2004. Sail stress treatments affect the phychological responses of seedling leaves of Pistacia chinensis Bunge. and Cercis chinensis Bunge. [J]. J Shandong Agric Univ(Nat Sci Ed), 35(2):173-176. [李国雷, 孙明高, 夏阳, 等, 2004. NaCl胁迫下黄栌、紫荆的部分生理生化反应动态变化规律的研究 [J]. 山东农业大学学报(自然科学版), 35(2):173-176.]
LI HS, 2000. Principles and techniques of plant physiology and biochemistry experiment [M]. Beijing: High Education Press. [李合生, 2000. 植物生理生化实验原理和技术 [M]. 北京: 高等教育出版社.]
LIU JH, YANG JM, ZHANG ZQ, et al., 2012. Effects of nickel application on the growth of soybean seedlings and nickel poisoning [J]. J Jilin Agric Univ, 34(3):311-315. [刘金华, 杨靖民, 张忠庆, 等, 2012. 黑土施镍对大豆幼苗生长的影响及镍中毒研究 [J]. 吉林农业大学学报, 34(3):311-315.]
LU Y, LI XR, HE MZ, et al., 2012. Effects of Ni and Cu stress on the activity of antioxidant enzymes in Peganum harmala [J]. Hay J, 21(3): 147-155. [鲁艳, 李欣荣, 何明珠, 等, 2012. Ni和Cu胁迫对骆驼蓬抗氧化酶活性的影响 [J]. 草业学报, 21(3):147-155.]
L XY, WU SQ, ZHANG Y, et al., 2014. Analysis on variation of main indicators of eutrophication and mutrition level in Taihu Lake [J]. J Water Resour Water Eng, 25(4):1-6. [吕学研, 吴时强, 张咏, 等, 2014. 太湖富营养化主要指标及营养水平变化分析 [J]. 水资源与水工程学报, 25(4) : 1-6.]
MOLAS J, 1997. Changes in morphological and anatomical structure of cabbage (Brassicea L.) outer leaves and in ultrastructure of their chloroplast caused by an in vitro excess of nickel [J]. Photosynthetica, 34(4):513-522.
WANG B, MA F, 2014. Effects of nickel stress on growth and physiological characteristics of 5 species of Cruciferae [J]. Agro-Technol Comm, 12:87-89. [王兵, 马芳, 2014. 镍胁迫对十字花科5种植物生长及生理特性的影响 [J]. 农业科技通讯, 12:87-89.]
WANG LN, 2014. Effects of nickel stress on physiological cha-racteristics, NPK nutrient absorption and yield of maize [D]. Changchun: Jilin Agricultural University. [王丽娜, 2014. 镍胁迫对玉米生理特性、氮磷钾营养元素吸收和产量的影响 [D]. 长春: 吉林农业大学.]
WU CX, WU HY, ZHU WB, et al., 2008. Physiological response and salt tolerance of halophytes in different saline soils [J]. J Anhui Agric Sci, 36(20):8450-8452. [武春霞, 吴海燕, 朱文碧, 等, 2008. 盐生植物在不同盐碱土壤中的生理反应及耐盐性 [J]. 安徽农业科学, 36(20):8450-8452.]
XU HF, LIU XT, JIN YM, et al., 2003. Study on sunflower chlorophyll and the specific leaf weight [J]. Agric Syst Sci Integr Res, 19(2): 97-100. [徐惠风, 刘兴土, 金研铭, 等, 2003. 向日葵叶片叶绿素和比叶重及其产量研究 [J]. 农业系统科学与综合研究, 19(2):97-100.]
YANG SJ, ZHANG BX, ZHANG XL, 2008. Evaluation and screening of salt and alkali resistance of different seed sources of American Gleditsia [J]. Prot For Sci Technol, (2):7-8,14. [杨树军, 张柏习, 张学利, 2008. 美国皂角不同种源耐盐碱评价与筛选 [J]. 防护林科技, (2):7-8, 14.]
ZHANG MD, SAINAO WQ, RAN RL, et al., 2018. Protective effect of attapulgite clay on Angelica sinensis seedlings under Cu stress [J]. Guihaia, 39(4): 453-463. [张牡丹, 赛闹汪青, 冉瑞兰, 等, 2018. Cu胁迫下基质中凹凸棒石粘土对当归幼苗的保护作用 [J]. 广西植物, 39(4): 453-463.]
ZHANG YQ, LIU SQ, YANG FJ, et al., 2003. Study on screening and salt tolerance mechanism of salt-tolerant watermelon rootstocks [J]. NW Agric J,12(4):105-108. [张云起, 刘世琦, 杨凤娟, 等, 2003. 耐盐西瓜砧木筛选及其耐盐机理的研究 [J]. 西北农业学报, 12(4):105-108.]
ZHANG ZA, ZHANG MS, WEI RH, 2004. Experimental gui-dance of plant physiology [M]. Beijing: China Agricultural Technology Press: 32-139. [张治安, 张美善, 蔚荣海, 2004. 植物生理学实验指导 [M]. 北京: 中国农业技术出版社: 32-139.]
ZHAO N, 2011. Effects of nickel stress on growth and physiological characteristics of maize and soybean seedlings [D]. Changchun: Jilin Agricultural University. [赵娜, 2011. 镍胁迫对玉米、大豆苗期生长及生理特性的影响 [D]. 长春: 吉林农业大学.]
ZHAO N, ZHOU MP, 2011. Effect of nickel stress on root growth and membrane protection system of maize seedlings [J]. J Anhui Agric Sci, 39(10):5821-5823. [赵娜, 周米平, 2011. 镍胁迫对玉米苗根系生长及膜保护系统的影响 [J]. 安徽农业科学, 39(10):5821-5823.]
ZHAO SJ, LIU HS, DONG XC, 1998. Experimental guidance of plant physiology [M]. Beijing: China Agricultural Science and Technology Press. [赵世杰, 刘华山, 董新纯, 1998. 植物生理学实验指导 [M]. 北京: 中国农业科技出版社.]
ZHENG AZ, 2005. Effects of nickel treatment with different nickel concentrations on physiological and biochemical indexes of maize seedlings [J]. J Anhui Agric Sci, 33(1): 2109-2145. [郑爱珍, 2005. 不同镍浓度镍处理对玉米幼苗生理生化指标的影响 [J]. 安徽农业科学, 33(1):2109-2145.]
ZOU Q, 2000. Experimental guidance on plant physiology [M]. Beijing: China Agricultural Press. [鄒琦, 2000. 植物生理学实验指导 [M]. 北京: 中国农业出版社.]
猜你喜欢
小读者(2021年2期)2021-03-29 05:03:48
少儿美术(2020年3期)2020-12-06 07:32:54
现代装饰(2020年11期)2020-11-27 01:47:48
中学生天地(A版)(2020年3期)2020-04-10 10:57:45
故事作文·高年级(2020年3期)2020-03-17 09:24:33
疯狂英语·新悦读(2019年11期)2019-12-18 05:14:16
华人时刊(2019年13期)2019-11-17 14:59:54
NBA特刊(2018年21期)2018-11-24 02:48:04
文苑(2018年22期)2018-11-19 02:54:14
商周刊(2017年6期)2017-08-22 03:42:35
