
6个稻瘟病抗性基因在浙江省主栽水稻品种中的分布和抗性评价
2019-09-10 07:22何海燕邱海萍柴荣耀毛雪琴王艳丽孙国昌
福建农业学报 2019年2期
何海燕 邱海萍 柴荣耀 毛雪琴 王艳丽 孙国昌




摘 要:【目的】检测浙江省40份水稻栽培品种和6份稻瘟病菌生理小种鉴别品种中6个抗性基因分布情况,同时鉴定品种苗瘟抗性水平,探讨携带抗性基因和抗性水平相關性。【方法】利用功能性分子标记检测品种中Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm、Pita,6个稻瘟病抗性基因的分布情况。用2015-2017年田间采集到的141个稻瘟菌菌株鉴定46个水稻品种苗瘟抗性水平。【结果】6个抗性基因在浙江省栽培品种中分布频率不同,其中Pita基因分布最广,占47.83%;其次是Pikh,占41.30%;抗性基因Pi2/Pizt 和Pikm在栽培品种中分布频率相等,均为34.78%;抗性基因Pi9/Piz在栽培品种中分布较少,仅有21.74%;抗性基因Pi1仅在稻瘟病菌生理小种鉴别品种TTP中检测到。抗性水平鉴定结果显示22个品种的抗性频率在60%以上,两个品种的抗性频率达到80%以上。【结论】浙江40份水稻栽培品种和6份稻瘟病菌生理小种鉴别品种携带抗性基因和其抗性水平部分相关。
关键词:水稻;苗瘟;分子标记;抗病基因
中图分类号:S 435.111.4+1 文献标识码:A文章编号:1008-0384(2019)02-214-09
0 引言
【研究意义】由子囊菌Magnaporthe grisea引起的稻瘟病是水稻三大病害之一,稻瘟病的爆发严重影响稻米的产量及品质。20世纪70年代,稻瘟病是浙江省晚稻主要病害,流行年份发病面积约占栽培面积的20%~30%,2014年浙江省晚稻稻瘟病在杭嘉湖、宁绍平原等地突发流行,发病面积达到2.25万hm2,浙粳88、中浙优1号等推广品种发病严重[1-2]。其原因在于带有某些稻瘟病主效抗性基因的推广品种在多年种植后抗性逐步丧失,并且品种区域布局过于单一化、集中化;在不利天气因素的影响下,田间病菌爆发,导致严重的产量损失。
培育品质良好的抗性品种是水稻育种家孜孜以求的目标,传统育种主要通过田间杂交和回交,再结合田间抗性鉴定和农艺性状的综合选择,经过多年多代筛选,选育抗性品种,这需要育种家有丰富的实践经验[3]。【前人研究进展】Andersen等[4]于2003年提出了基因功能标记(Functional markers,FMs)的概念,指一个分子标记位点代表一个特定的等位基因,通过对分子标记的筛选即能对性状进行筛选。目前,育种家将分子标记结合到传统育种技术中,旨在实现快速、准确、系统地鉴定水稻品种抗瘟基因型,达到培育抗性强及抗谱广的水稻品种的目的。一些已经被定位或克隆的抗性基因,比如Pi9、Pigm、Pita、Pikm等已经在生产上广泛运用[5-8]。范方军等[9]利用水稻稻瘟病抗性基因Pi-b、Pita等4个基因的功能标记检测江苏省64份水稻品系,结合穗颈瘟抗性鉴定,解析4个稻瘟病抗性基因在江苏省粳稻稻瘟病抗性育种中的作用。宋兆强等[10]利用Pita、Pib等4个基因的功能标记,对在稻瘟病菌圃经多年抗性筛选的60份资源材料进行基因型鉴定,并进行抗性基因与穗颈瘟发病相关性分析,发现所有基因的抗病能力都在不断减弱。
【本研究切入点】了解品种本身抗病基因型,可以避免育种上抗病基因利用的盲目性。由此,本研究利用Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm、Pita 6个稻瘟病抗性基因功能性分子标记,对浙江省46个水稻栽培品种和稻瘟病菌生理小种鉴别品种,包括籼型三系杂交稻、粳型三系杂交稻、籼型两系杂交稻、粳型常规稻、籼型常规稻等进行了抗瘟基因型检测,并且对这些品种进行了稻瘟菌苗期接种、抗苗瘟水平分析。【拟解决的关键问题】本研究旨在明确浙江省水稻栽培品种的基因型,初步探讨水稻基因型和抗瘟水平的相关性,为浙江省稻区抗病品种的合理布局提供依据。
1 材料与方法
1.1 水稻材料及种植条件
供试水稻品种包括阳性对照材料丽江新团黑谷单基因系品种8个,为Pi1、Pi9、Piz、 Pi2、 Pizt、 Pikh、 Pikm、Pita基因供体品种IRBL1-CL、IRBL9-W、IRBLz-Fu、IRBLZ5-CA、IRBLZt-T、IRBLkh-k3、IRBLkm-Ts和IRBLta-K1,由粮食作物“基因对基因”病害农业部行业专项提供。阴性感病对照材料丽江新团黑谷1份,浙江省水稻栽培品种 40份,稻瘟病菌生理小种鉴别品种6份;由浙江省农业科学院植物与微生物研究所提供。
将水稻种子置于 28℃培养箱浸种 2 d,32℃催芽 48 h,选取发芽良好的种子穴播于盛有肥沃土壤的育苗盆内,穴间距为 5 cm 左右,每穴播种发芽良好的种子10粒左右,每盆中分别设感病对照 1 份,各重复内品种采用随机排列;2 叶期定苗酌施氮肥,促苗以利于做接种试验。在温室中培养 14 d 后,进行样品采集和接种。
1.2 病原菌材料、培养及接种调查
1.2.1 菌株来源、活化、繁殖和产孢 供试菌株为2016-29-2、2017-58-1、2016-42-1等141个菌株(由浙江省农业科学院植物保护与微生物研究所提供),所有菌株为2015-2017年间分离自浙江各地采集的稻瘟菌标样。菌株在PDA平板上活化后,接种于燕麦固体培养基上, 26℃下 12 h 光暗交替培养 10 d。加入适量无菌水刷洗培养基里的孢子,双层纱布过滤后,观察稻瘟病菌产孢情况,用血球计数板将孢子悬浮液浓度调节至约 2× 105个·mL-1,作为苗期喷雾的接种体。
1.2.2 水稻叶片喷雾接种和发病调查 待水稻生长 14 d 左右,即在幼苗三叶一心期,在悬浮液中加入 0.5%~0.8%的 Tween20 进行喷雾接种,每 100 株苗约喷 20mL(以菌株薄雾均匀沾布全部叶片为度);于 26~28℃下黑暗保湿(相对湿度约95%) 24 h,然后将育苗盘移至 28℃的高湿(用喷雾器进行不定时喷雾)环境下培育。
在水稻接种 7 d 左右进行调查:以感病对照品种发病程度判断试验的有效性。以病斑是否典型,大小及数量记载病情。本试验中丽江新团黑谷单基因系和浙江省栽培品种的抗瘟性等级,以水稻苗期调查9级分级标准[11]。0级,没有症状,免疫;1级,针头状大小褐色小点,高抗;2级,直径≤1 mm的褐色病斑,抗病;3级,灰色病斑,边缘褐色,病斑直径约1~2 mm,中抗;4级,典型纺锤形病斑,长3 mm以上,通常在两条叶脉之间,为害面积不超过叶面积2%,中感;5~7级,典型病斑,为害面积占叶面积10%~50%,感病;8~9级,典型病斑,为害面积51%以上,高感。抗性频率(%)为水稻材料在稻瘟菌接种中表现抗性菌株数与总测定菌株数目之比。1.3 样品采集及DNA提取
每个品种采集1个单株的2 cm长叶片,置于2 mL离心管(提前放置2~3颗钢珠)中,经液氮速冻后,置于-80℃的冰箱里储藏备用。
用TPS法抽提水稻叶片总DNA。在每个放置样品的离心管中加入800 μL的TPS(100 mmol·L -1 Tris-Cl,pH值8.0;10 mmol·L -1 EDTA,pH 值8.0;1 mol·L -1 KCl)抽提液。将离心管放入组织研磨仪,70 Hz、90 s研磨2次。 将磨碎的叶片放入65℃水浴中温浴30 min。12 000 r·min-1离心10 min。 取上清液于1.5 mL的离心管,加入等体积的氯仿/异戊醇(24∶1),混匀,12 000 r·min-1离心5 min。取上清加入等体积的异丙醇,混匀,室温放置10 min, 12 000 r·min-1离心5 min。弃上清,加入800 μL 75%乙醇,洗涤沉淀,12 000 r·min-1离心2 min,弃上清,控干残液,室温(37℃)放置使沉淀干燥,加入100 μL ddH2O(含RNA酶)溶解,37℃温浴40 min,然后-20℃保存。
1.4 Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm、Pita等6个基因特异性分子标记与检测
1.4.1 Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm、Pita等6个基因特异性分子标记
本研究检测Pi1 [12-13]、Pi9/Piz[14-15]、Pi2/Pizt[15-16]、Pikh[17-18]、Pikm[19-20]、Pita[21-22]6个稻瘟病抗性基因,其特异性分子标记的引物信息见表1。引物由上海擎科生物技术公司合成。
1.4.2 Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm、Pita 等6个基因特异性分子标记的检测
采用PCR扩增方法检测。用20 μL PCR反应体系,含1 μL(40 ng)Template DNA,0.5 μLForward Primer(10 μm), 0.5 μL Reverse Primer(10 μm),10 μL 2×TSINGKE Master Mix,8 μL ddH2O。 PCR反应条件:94℃预变性2 min;然后94℃变性30 s,50~60℃退火30 s,72℃延伸1 kb·min-1,将这三步进行35个循环;最后72℃ 5 min。制备 1.0%~2.5%的琼脂糖凝胶,电压 90~140V 下电泳15~45 min。最后,在紫外线凝胶成像系统中观察拍照并记录试验结果。
2 结果与分析
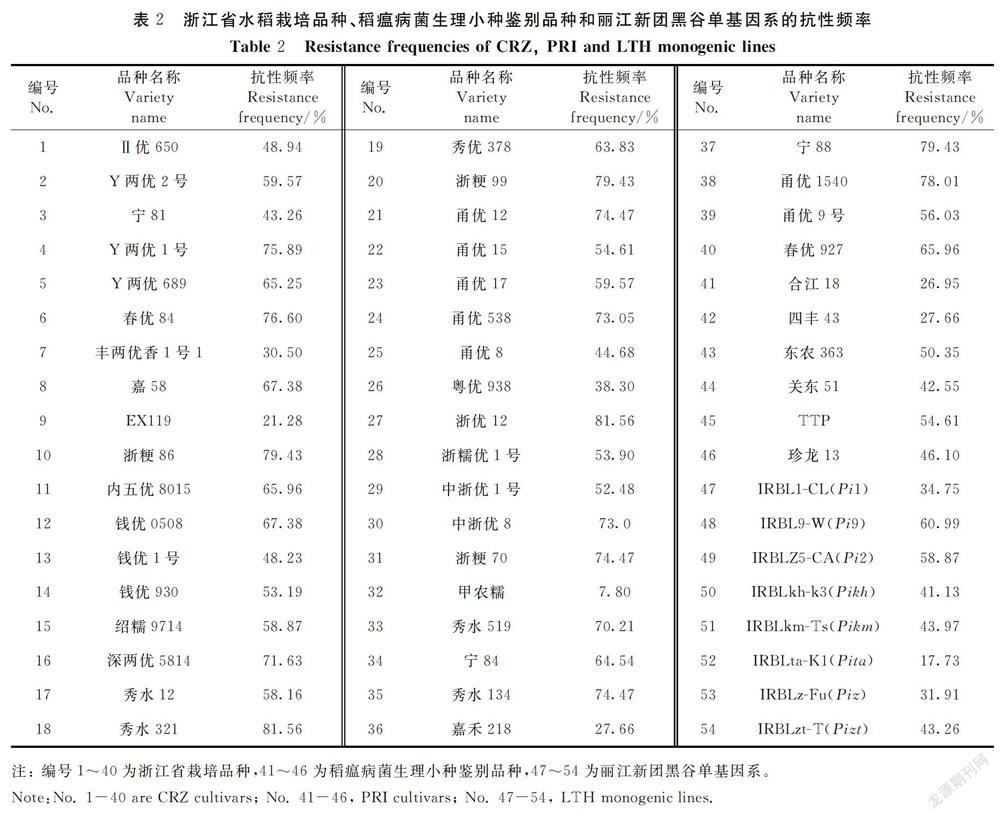
2.1 供试栽培品种、稻瘟病菌生理小种鉴别品种和丽江新团黑谷单基因系的抗性频率
本研究利用田间收集的141个稻瘟病菌对40个浙江省栽培品种、6个稻瘟病菌生理小种鉴别品种,以及8个含有对应抗性基因的丽江新团黑谷单基因系进行了苗期抗性测定。结果显示,在 40个浙江省水稻栽培品种中,有2个水稻品种抗性频率达到80%以上,分别为秀水321和浙优12;有 13个品种抗性频率在70%~80%,分别为Y两优1号、春优84、浙粳86、深两优5814、浙粳99、甬优12、甬优538、中浙优8号、浙粳70、秀水519、秀水134、宁88、甬优1540;有 6个品种抗病频率在60%~70%,分别为Y两优689、嘉58、内五优8015、钱优0508、宁84、春优927。6个稻瘟病鉴别品种中,抗性频率最高的是TTP,为54.61%;抗性频率最低的是合江18,为26.95%。8个丽江黑谷单基因系中,抗性频率最高的是IRBL9-W(Pi9)和IRBLZ5-CA(Pi2),分别为60.99%和58.87%,说明Pi9和Pi2基因的抗谱较广,同前人研究吻合。抗性频率最低的是IRBLta-K1(Pita)和IRBLzt-T(Pizt),均为17.73%,其余几个单基因系的抗性频率在30%~45%。鉴定结果如表2所示。
2.2 供试栽培品种和稻瘟病菌生理小种鉴别品种的抗瘟基因型及其分布
利用已建立的分子鉴别体系对 40个浙江省水稻栽培品种和6个稻瘟病鉴别品种的基因分布情况进行了研究,获得了部分稻瘟病抗性基因的分布情况(表3)。结果发现在这 46个品种中,有1个品种携带 Pi1抗病基因,为稻瘟病菌生理小种鉴别品种TTP。有10个品种(其中两个为稻瘟病菌生理小种鉴别品种)携带Pi9/Piz 抗病基因,其中9个为纯合基因型,1个为杂合型。有16個栽培品种携带 Pi2/Pizt 抗病基因,其中13个为纯合基因型,3个为杂合基因型。有19个品种(其中2个为稻瘟病菌生理小种鉴别品种)携带 Pikh 抗病基因,其中10个为纯合基因型,9个为杂合基因型。有16个品种(其中2个为稻瘟病菌生理小种鉴别品种)携带 Pikm 抗病基因,有22个品种(其中1个为稻瘟病菌生理小种鉴别品种)携带 Pita抗病基因,其中10个为纯合基因型,12个为杂合基因型。在检测的水稻品种中,抗性基因Pita和Pikh分布频率最高,分别为47.83%和41.30%;其次是抗性基因Pikm和Pi2/Pizt,分别为39.13%和34.78%; Pi9/Piz和Pi1分布频率为21.74%和2.17%。部分品种分子标记检测结果如图1所示,各个抗性基因分布频率如图2所示。
2.3 供试品种抗瘟基因数量分析
对40个浙江省水稻栽培品种和6个稻瘟病菌生理小种鉴别品种检出的稻瘟病抗性基因进行数量分析,春优84、浙粳70两个栽培品种同时携带4个抗性基因,占检测品种比率为4.35%;宁81、浙粳86、绍糯9714、秀水12号等15个品种(其中2个为稻瘟病菌生理小种鉴别品种)同时携带3个抗病基因,占检测品种比率为32.61%;Y两优689、嘉58和钱优0508等13个栽培品种同时携带2个抗病基因,占测试品种的23.91%;有12个品种(其中1个为稻瘟病菌生理小种鉴别品种)仅携带1个抗病基因,占测试品种的26.09%;还有6个品种未携带任何抗病基因,其比率为13.04%。
2.4 检出的抗瘟基因数量、类型与品种抗病性的相关性分析
为了探明浙江省栽培水稻品种和稻瘟菌检测品种抗瘟基因类型或数量与品种抗病性之间的相关性,本研究对46个水稻品种进行了稻瘟病苗叶瘟检测,并用6个稻瘟病抗性基因分子标记对46个品种进行了基因型检测。结果表明,其中28个品种(占比为60.87%)含有2个或2个以上抗性基因,这28个品种中18个品种的抗性频率高于60%,仅有1个品种抗性频率低于50%,而含有对应基因的8个丽江新团黑谷单基因系中有6个抗性频率在45%以下,说明聚合了2個或2个以上抗性基因的材料抗性频率明显提高。随着品种中检出的抗性基因数量增多,品种的抗性频率也明显上升。含有4个抗病基因的2个品种,抗性频率都超过70%;在含有3个抗病基因的15个品种中,14个品种抗性频率都在50%以上。春优84聚合了Pi2/Pizt、Pikh、Pikm、Pita 4个抗病基因,其中3个为杂合基因型,抗性频率达76.60%;浙粳86、宁88和甬优1540聚合了Pi2/Pizt、Pikh、Pita等3个抗性基因,抗性频率分别达79.43%、79.43%和78.01%; 秀水321和浙粳99聚合了Pi2/Pizt、Pikm、Pita 等3个抗性基因, 其中2个为纯合型,抗性频率达81.56%和79.43%。说明抗性频率高的品种基本都聚合了多个抗性位点的多个抗性基因。同时没有聚合Pi2/Pizt、Pikh、Pikm、Pita 其4个抗性位点中3个位点,抗性频率却低于70%的品种,说明Pi2/Pizt、Pikh、Pikm、Pita等4个抗病基因在浙江省栽培品种中起到了主要的抗性效用。
3 讨论与结论
了解水稻品种稻瘟病抗病基因的数量和构成情况是利用抗病品种应对稻瘟病的基础。新团丽江黑谷单基因系的构建、稻瘟病抗性基因的克隆和特异性分子标记的开发为人们分析水稻品种中稻瘟病抗性基因提供了技术支持[13,15]。本研究以浙江省水稻栽培品种和稻瘟病菌生理小种鉴别品种为主要检测对象,以期明确上述品种中Pi1、Pi9/Piz、Pi2/Pizt、Pikh、Pikm 和Pita这6个基因位点的分布情况,为本地区广谱抗瘟品种的培育和各栽培品种的合理布局提供科学依据。
有学者研究发现,同时携带Pi1和Pi2两个抗性基因的水稻品种,对稻瘟病的抗性效果明显好于单独携带Pi1和Pi2基因的水稻品种,认为这两个基因存在互补关系[23-26]。于苗苗等[27]在2013年研究指出Pi1和Pi2聚合后不同遗传背景的材料抗性频率均达到90%以上。本研究中发现浙江省水稻栽培品种中有16个材料有Pi2/Pizt基因,然而携带Pi1基因的材料基本没有,因此在今后的育种研究中可以考虑培育聚合Pi1和Pi2基因的品种,提高品种抗病性。
前人研究发现抗病鉴定结果与标记筛选出抗性基因数量多寡之间有一定的差异,分析可能原因认为目前已经定位的抗性基因达到100个以上,而实验中检测的基因数量非常有限,并不能代表品种中实际含有的抗性基因情况[28]。比如品种深两优5814仅含有Pita 1个抗性基因,为杂合型,抗性频率却达到了71.63%;品种中浙优8号没有检测到此6个抗性基因中的任何一个,抗病频率却达到73.05%。说明这2个品种中可能存在其他抗谱广的抗性基因,值得进一步研究利用。
本研究中使用的Pi9和Piz基因、Pi2和Pizt基因的抗感显性分子标记一致,然而分别携带Pi9和Piz基因的丽江新团黑谷单基因系材料抗谱并不一致,分别携带Pi2和Pizt基因的丽江新团黑谷单基因系材料抗谱也不一致,说明Pi9和Piz, Pi2和Pizt基因同源程度比较高,在进化过程中分化比较晚,但是它们对水稻稻瘟病抗性水平有明显不同。文中检测到10个品种携带Pi9/Piz基因,然而它们的抗性频率均不到60%(低于携带Pi9基因的丽江黑谷单基因系,其抗性频率为60.99%,携带Piz基因的丽江黑谷单基因系,其抗性频率为31.91%),可以推测这10个品种携带了Piz基因而非Pi9基因。 在今后的研究中可以进一步开发能区分Pi2和Pizt基因、Pi9和Piz基因的特异性分子标记,进一步明确品种基因型,更有针对性地聚合相应抗性基因的材料。
20世纪末有学者研究指出水稻稻瘟病防治面临的挑战在于稻瘟病菌的变异机制和品种抗性丧失机理并不明朗[29]。不同稻瘟菌在不同品种上寄生适合度存在差异,当一个品种推广种植数年后,常会引起病菌群体小种组成结构的变化,继而导致品种抗性丧失,培育稻瘟病抗性品种需要分析稻瘟病菌群体的地理分布和致病性分化[30-31];因此田间水稻品种的推广应当注意合理布局,适时轮换,避免抗性品种的感病化,延长品种的使用年限。同时因品种中抗性基因分布情况是指导品种合理布局的基础,因此今后仍有必要拓展不同类型抗瘟基因研究分析的深度和广度,加大新的抗稻瘟病基因的利用,以及通过基因聚合等手段培育广谱抗病品种。
参考文献:
[1]郑永利,施德,姚晓明,等.晚稻稻瘟病局部突发流行成因与治理对策探讨[J].浙江农业科学,2015,56(1):92-93.
ZHENG Y L,SHI D,YAO X M,et al. Discussion on the causes of local outbreak of rice blast in late rice and its control countermeasures [J]. Journal of Zhejiang Agricultural Sciences, 2015,56(1):92-93.(in Chinese)
[2]谢子正,许渭根,李仁忠,等. 2014年浙江省水稻稻瘟病流行特点及原因分析[J].中国植保导刊,2015(3):58-60.
XIE Z Z,XU W G,LI R Z,et al. Epidemiological characteristics and causes of rice blast in Zhejiang Province in 2014 [J]. China Plant Protection,2015(3):58-60.(in Chinese)
[3]向聪,雷东阳,任西明,等.水稻抗稻瘟病遗传育种研究进展[J].作物研究,2017(5):547-552.
XIANG C,LEI D Y,REN X M,et al. Research progress in genetic and breeding of rice blast resistance [J].Crop Research,2017(5):547-552.(in Chinese)
[4]ANDERSEN J R,LUBBERSTED T.Functional markers in plants[J].Trends in Plant Science,2003,8(11):554-560.
[5]譚令辞,刘雄伦,杨婷婷,等. 利用Pi9基因分子标记辅助选择培育抗稻瘟病水稻新品系[J]. 作物研究, 2015, 29(4):348-351.
TAN L C,LIU X L,YANG T T,et al. Breeding new rice line with Blast Resistant by molecular marker-assisted selection of the Pi9 Gene [J].Crop Research, 2015, 29(4):348-351.(in Chinese)
[6]行璇,刘雄伦,陈海龙,等. 分子标记辅助选择Pi9基因改良R288的稻瘟病抗性[J]. 作物研究,2016,30(5):487-491.
XING X, LIU X L, CHEN H L, et al. Improving blast resistance of rice restorer R288 by molecular marker-assisted selection of Pi9 Gene [J].Crop Research,2016,30(5):487-491.(in Chinese)
[7]梁毅,杨婷婷,谭令辞,等. 水稻广谱抗瘟基因Pigm紧密连锁分子标记开发及其育种应用[J]. 杂交水稻, 2013, 28(4):63-74.
LIANG Y, YANG T T, TAN L C, et al. Development of the molecular marker tightly-linked with the broad-spectrum blast resistance gene Pigm and its breeding practice in Rice [J]. Hybrid Rice , 2013, 28(4): 63-74.(in Chinese)
[8]杨丰宇,李永聪,刘雄伦,等.分子标记辅助选择改良早籼稻1701的稻瘟病抗性[J]. 分子植物育种, 2017(6):2212-2217.
YANG F Y, LI YONGC, LIU X L, et al. Improving blast resistance of early indica rice 1701 by molecular marker-assisted selection for [J].Molecular Plant Breeding, 2017(6):2212-2217.(in Chinese)
[9]范方军,王芳权,刘永峰,等. Pi-b、Pi-ta、Pikm和Pi54对水稻穗颈瘟的抗性评价[J]. 华北农学报,2014, 29(3):221-226.
FAN F J, WANG F Q, LIU Y F, et al. Evaluation of resistance to rice panicle blast with resistant genes Pi-b, Pi-ta, Pikm and Pi54 [J].Acta Agriculturae Boreali-Sinica,2014, 29(3):221-226.(in Chinese)
[10]宋兆强, 刘艳, 王宝祥,等. 稻瘟病抗性基因Pi-ta、Pi-b、Pi54和Pi-km的育种利用价值评价[J]. 江苏农业学报, 2017,33(5):968-974.
SONG Z Q, LIU Y, WANG B X, et al. [J]Application value of blast resistance genes Pi-ta, Pi-b, Pi54 and Pi-km in rice breeding [J]. Jiangsu Journal of Agricultural sciences, 2017, 33 (5): 968-974.(in Chinese)
[11]International Rice Research Institute. Standard Evalution System for Rice[M]. 4th ed. Manila: IRRI, 1996:17-18.
[12]HUA L X,WU J Z, CHEN C X, et al. The isolation of Pi1, an allele at the Pik locus which confers broad spectrum resistance to rice blast[J]. Theoretical & Applied Genetics, 2012, 125(5):1047-1055.
[13]刘开强,伍豪,颜群,等. 水稻抗稻瘟病基因Pi1的特异性分子标记开发及利用[J]. 西南农业学报, 2016, 29(6):1241-1244.
LIU K Q, WU H, YAN Q, et al. Development and application of specific molecular markers of blast resistance gene Pi1 in rice [J]. Southwest China Journal of Agricultural Sciences, 2016, 29 (6): 1241-1244.(in Chinese)
[14]QU S H, LIU G F, ZHOU B, et al. The broad-spectrum blast resistance gene Pi9/Piz encodes a nucleotide-binding site-leucine-rich repeat protein and is a member of a multigene family in rice[J].Genetics, 2006, 172(3): 1901-1914.
[15]高利军,高汉亮,颜群,等.4个抗稻瘟病基因分子标记的建立及在水稻亲本中的分布[J].杂交水稻,2010,25(S1):294-298.
GAO L J, GAO H L, YAN Q, et al. Establishment of markers for four blast genes and marker distribution in rice parents [J].Hybrid Rice, 2010, 25 (S1): 294-298.(in Chinese)
[16]ZHOU B, Qu S H, LIU G F, et al.The eight amino-acid differences within three leucine-rich repeats between Pi2 and Piz-t resistance proteins determine the resistance specificity to Magnaporthe grisea[J]. Molecular Plant-Microbe Interactions, 2006, 19(11): 1216-1228.
[17]SHARMA T R, MADHAY M S, SINGH B K, et al. High-resolution mapping, cloning and molecular characterization of the Pi-k (h) gene of rice, which confers resistance to Magnaporthe grisea[J]. Molecular genetics and genomics: MGG, 2005, 274(6): 569-578.
[18]王軍,杨杰,朱金燕,等.稻瘟病抗病基因Pi-kh功能标记的开发及江苏粳稻品种中Pi-kh的变异[J]. 中国水稻科学,2014,28(2):141-14.
WANG J, YANG J, ZHU J Y, et al. Development of a functional marker for rice blast resistance gene Pi-kh and natural variation at Pi-kh in Japonica rice in Jiangsu province [J].Chinese Journal of Rice Science, 2014, 28 (2): 141-147.(in Chinese)
[19]ASHIKAWA I, HAYASHI N, YAMANE H, et al. Two adjacent nucleotide-binding site-leucine-rich repeat class genes are required to confer Pikm-specific rice blast resistance[J]. Genetics, 2008, 180(4): 2267-2276.
[20]徐小金. 稻瘟病抗性基因资源的挖掘及其育种利用[D].金华:浙江师范大学,2016.
XU X J. Exploitation and breeding application of rice blast resistance gene resources [D]. Jinhua: Zhejiang Normal University, 2016.(in Chinese)
[21]BRYAN G T, WU K S, FARRALL L, et al. A single amino acid difference distinguishes resistant and susceptible alleles of the rice blast resistance gene Pi-ta[J]. The Plant Cell Online, 2000, 12(11): 2033-2045.
[22]JIA Y L, WANG Z H, SINGH P. Development of Dominant Rice Blast, Pi-ta, Resistance Gene Markers[J]. Crop Science, 2002, 42(6):2145-2149.
[23]JIANG H C, FENG Y T, BAO L, et al. Improving blast resistance of Jin 23B and its hybrid rice by marker-assisted gene pyramiding[J]. Molecular Breeding, 2012, 30(4):1679-1688.
[24]陈红旗,陈宗祥,倪深,等. 利用分子标记技术聚合3个稻瘟病基因改良金23B的稻瘟病抗性[J]. 中国水稻科学, 2008, 22(1):23-27.
CHEN H Q, CHEN Z X, NI S, et al. Pyramiding three genes with resistance to blast by marker-assisted to improve rice blast resistance of Jin23B [J]. Chinese Journal of Rice Science, 2008, 22(1): 23-27.(in Chinese)
[25]柳武革,王丰,金素娟,等. 利用分子标记辅助选择聚合Pi-1和Pi-2基因改良两系不育系稻瘟病抗性[J]. 作物学报, 2008,34(7):1128-1136.
LIU W G, WANG F, JIN S J, et al. Improvement of rice blast resistance of TGMS Line by pyramising of Pi-1 and Pi-2 through molecular marker-assisted selection [J].Acta Agromomica Sinica, 2008, 34(7): 1128-1136.(in Chinese)
[26]DIVYA B, ROBIN S, RABINDRAN R, et al. Marker assisted backcross breeding approach to improve blast resistance in Indian rice ( Oryza sativa) variety ADT43[J]. Euphytica, 2014, 200(1):79.
[27]于苗苗,戴正元,潘存紅,等. 广谱稻瘟病抗性基因Pigm和Pi2的抗谱差异及与Pi1的互作效应[J]. 作物学报, 2013, 39(11):1927-1934.
YU M M, DAI Z Y, PAN C H, et al. Resistance spectrum difference between two broad-spectrum blast resistance genes, Pigm and Pi2 , and their interaction effect on Pi1 [J].Acta Agromomica Sinica, 2013, 39 (11): 1927-1934.(in Chinese)
[28]邢鹏,张幸,李冬梅,等.稻瘟病抗性基因在主要育种亲本中的分布研究[J]. 分子植物育种,2015, 13(3):505-512.
XING P, ZHANG X, LI D M, et al. Research on the distribution of blast resistance genes in main rice breeding parents [J].Molecular Plant Breeding, 2015, 13(3): 505-512.(in Chinese)
[29]孙国昌,杜新法,陶荣祥,等. 水稻稻瘟病防治策略和21世纪研究展望[J].植物病理学报,1998(4):289-292.
SUN G C, DU X F, TAO R X,et al. Control tactics and prospect of rice blast research in 21th century [J]. Acta Phytopathologica Sinica, 1998 (4): 289-292.(in Chinese)
[30]柴荣耀,杜新法,毛雪琴,等.水稻不同类型品种上的稻瘟病菌群体小种的演变[J]. 浙江农业学报,1999, 11(6):297-300.
CHAI H, DU X F, MAO X Q, et al. Race evolution of Magnaporthe grisea on different rice varieties [J].Acta Agriculturae Zhejianggensis, 1999, 11(6): 297-300.(in Chinese)
[31]任鄄胜,肖陪村,陈勇,等. 水稻稻瘟病病菌研究进展[J]. 现代农业科学, 2008(1):19-23.
REN J S,XIAO Y C,CHEN Y, et al. The research progress on rice blast fungus [J]. Modern Agricultural Science, 2008 (1): 19-23.(in Chinese)
(责任编辑:林海清)
猜你喜欢
世界热带农业信息(2021年7期)2021-08-11
好孩子画报(2021年3期)2021-04-02
农民致富之友(2020年8期)2020-05-11
农民致富之友(2019年23期)2019-08-16
农产品市场周刊(2017年35期)2017-10-25
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
江苏农业科学(2016年8期)2017-02-15
山东农业科学(2016年12期)2017-01-21
天津农业科学(2016年12期)2017-01-11
