
不同退化程度下的高寒草甸主要温室气体通量
2019-09-05 08:57:38郭小伟戴黎聪李以康张法伟钱大文朋措吉杜岩功曹广民
水土保持研究 2019年5期
郭小伟, 戴黎聪,2, 李以康, 张法伟, 林 丽, 李 茜,钱大文, 樊 博, 柯 浔,2, 舒 铠,2, 朋措吉,2, 杜岩功, 曹广民
(1.青海省寒区恢复生态学重点实验室, 中国科学院 西北高原生物研究所, 西宁810001; 2.中国科学院大学, 北京100039)
工业革命以来人类活动使用大量的化石燃料,导致大气中温室气体(CO2、CH4和N2O)浓度迅速增加,三种温室气体年增长速率分别为1.5 ml/m3,4 μl/m3,0.8 μl/m3[1],改变了陆地生态系统C、N平衡,导致全球变暖、氮沉降增加,由此而引起的全球环境变化改变了生态系统生产力[2],碳、氮循环[3]。土壤呼吸是陆地生态系统向大气输出碳素的主要形式[4],全球每年因土壤呼吸产生大约80.4 PgCO2-C,大约是燃烧化石燃料和砍伐森林释放碳的10倍[5],因此,土壤呼吸很小的变化都可能影响全球大气碳和热量平衡[6]。温度的升高,促进土壤微生物活性,进一步激发CH4和N2O气体释放[7],温室气体排放与全球变暖之间的正反馈效应对陆地生态系统稳定性以及可持续发展造成重要影响,近年来成为科学界关注的热点问题之一[8-9]
青藏高原高寒草地约为1.28亿km2,涵养着我国五大水系:黄河、长江、澜沧江、怒江、雅鲁藏布江,是我国巨大的草地畜牧业生产基地、生态安全的重要屏障和多民族生存的重要家园,同时,高寒草地是“世界第三极”重要的碳库,对该地区生态系统的碳库的平衡具有重要的调节作用[10]。据政府间气候变化专门委员会(IPCC)最新预测,到2100年全球气温将上升1.8~4.0℃[11]。青藏高原高寒草地生态系统温室气体排放的研究已经有很多重要结论,高寒草地是大气CO2的弱汇,温度是控制土壤呼吸的主要因子,能够解释其44%的变异[12],青藏高原高寒草地年CH4年吸收量约为0.2 Tg[13],高寒草地是重要的大气N2O源,年N2O贡献量达0.3 Tg[14],而CO2的年贡献量达46.5 Tg[15]。但是这些研究大多着重于某一类型高寒草地的温室气体排放或几种草地的温室气体排放研究,对于草地退化过程中主要温室气体通量研究较少,退化高寒草地面积占青藏高原高寒草地总面积的80%,因此对退化高寒草地温室气体通量研究是非常有必要的。关于青藏高原高寒草甸温室气体通量的研究助于理解本地区草地生态系统对于全球气候变化的响应,尤其是关于草地退化演替过程的研究,更加有利于明晰草地温室气体产生与消耗机制。
本研究利用2014年建立青海省海北州皇城乡建立的高寒草甸放牧演替平台,研究高寒草甸退化过程中的环境要素关键变化和主要温室气体通量特征,并且同期收集各类环境因子和植被特征变化,采用逐步回归的手段,明确高寒草甸主要温室气体通量的关键影响因子,以期为草地周转和草地温室气体减排提供理论支撑。
1 材料与方法
1.1 研究区地理位置
试验设布置于中科院海北高寒草甸生态系统试验观测站附近,其坐落在青藏高原东北隅,祁连山北支冷龙岭东段南麓的大通河河谷,位于101°19′E,37°37′N,站区平均海拔3 280 m。年平均温度-1.7℃,最暖月为7月(9.8 ℃)和最冷月1月左右(-14.8 ℃),年均降水量618 mm,主要集中在生长季(数据来自中科院高寒草甸海北定位研究站)。
1.2 样地设置与样品采集
样地设置于青海省海北州皇城乡,根据植被生长状况,设施原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸,本地区草地退化使由于放牧家畜种类的改变,草地退化年限为第6年,不同退化阶段高寒草甸按照青海省地方标准《高寒嵩草草甸退化状态评估》DB63/T1414—2015进行划分(表1)。于植物分布比较均匀,地形平坦地段进行试验,设置地框进行静态箱温室气体采集。其中:(1) 原生草地放牧数量为3.6羊单位/hm2,植被高度5~12 cm,盖度90%~100%,具有明显的上下层结构,上层以针茅(Stipaspp)和羊茅(Festucaspp.)等禾本科植物为优势种,下层以矮嵩草(Kobresiahumilis)等莎草为优势种,禾本科植物出现的频率和盖度高于莎草科植物,草毡表层厚度不高于5 cm;放牧时间为每年3—5 月;(2) 轻度退化高寒草甸放牧数量为7.5羊单位/hm2,草地植被高度3~7 cm,盖度80%~95%,上层以针茅(Stipaspp)和羊茅(Festucaspp.)等禾本科植物为优势种,下层以矮嵩草(K.humilis)等莎草科植物为优势种,矮嵩草植物的频度及盖度高于禾本科植物,草毡表层厚度5 cm 左右,放牧时间每年的2 月至5 月;(3) 中度退化草甸,演替所处阶段为小嵩草草甸草毡表层加厚期,草地为单片层结构,禾本科植物出现的频率小,小嵩草(K.pygmaea)等莎草科植物为优势种,并呈明显的斑块化分布,小嵩草斑块边缘出现一定深度的裂缝,植被高度2~4 cm,植被盖度40%~50%;(4) 重度退化草甸,演替所处阶段为小嵩草草甸草毡表层开裂期,草地为单片层结构,禾本科植物出现的频率小,小嵩草(K.pygmaea)等莎草科植物为优势种,并呈明显的斑块化分布,小嵩草斑块中心区域存在大量空地,植被高度1~4 cm,植被盖度20%~40%。
静态箱法测定温室气体通量,采用50 cm ×50 cm×50 cm的箱体,箱体外包有白色塑料泡沫和白色纱布,不透明,由于采用用的静态箱法为暗箱,所以这里的草地二氧化碳通量仅表示草地的呼吸能力。于2017年10月到2018年9月采集温室气体样品,生长季和休眠季每月分别进行2次和1次气体样品采集。
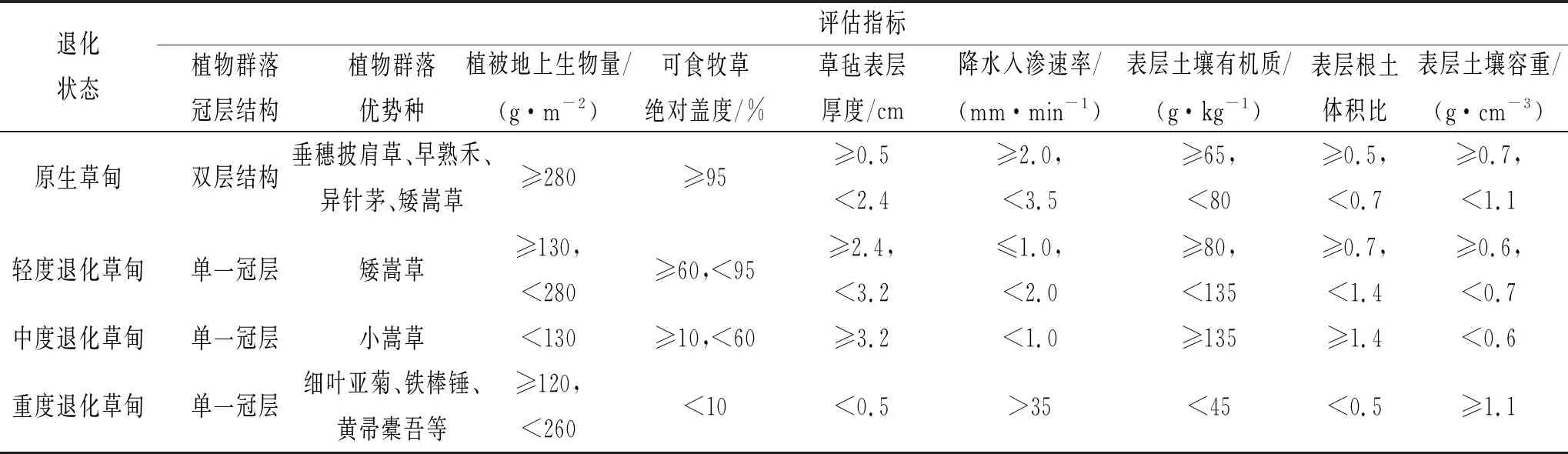
表1 高寒嵩草草甸退化状态评估指标
1.3 样品分析
气体测定采用气相色谱法(HP4890D,Agilent)。检测器为FID离子火焰化检测器,分离柱为SS-2 m×2 mm×13 XMS(60/80),检测器温度为230℃,分离柱温度为55℃。标准气中CH4浓度为2.24 μl/L,最小检测限0.08 μl/L[16]。
1.4 计算公式
通量计算公式:
式中:F为温室气体通量;V是静态箱内空气体积;A是静态箱覆盖面积;Ct是t时刻箱内被测气体的体积混合比浓度;t是时间;ρ是标准状态下的被测气体密度;T0和P0分别为标准状况下的空气绝对摄氏温度和气压;P为采样地点的气压;T为采样时的绝对温度。
1.5 数据处理
温室气体通量和土壤温度以及湿度的关系采用相关性分析、不同草地温室气体通量间差异比较采用单因素方差分析使用SPSS 20.0统计软件。
2 研究结果
2.1 退化演替过程中甲烷通量特征
自2016年10月至2017年9月进行的高寒草甸甲烷通量观测显示,在原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸甲烷通量均表现为大气甲烷汇,4种草地甲烷通量存在明显季节动态趋势,即生长季甲烷吸收更多,而休眠季甲烷吸收较少(图1),甲烷吸收速率最大值均出现在6月,休眠季和生长旺季(7月、8月)重度退化草甸显著高于其余3个阶段的草甸(p<0.05),而在生长季初期(5月、6月)不同退化阶段草甸甲烷通量差异不明显。
原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸甲烷通量分别为-40.09,-37.78,-45.06 ,56.66 μg/(m2·h)。其中中度退化和重度退化草甸甲烷通量显著高于原生草甸和轻度退化草甸(p<0.05)。高寒草甸在退化过程中甲烷吸收能力增加,其中重度退化阶段草甸甲烷通量比原生草甸阶段甲烷吸收能力增加41.33%。

注:图中误差线代表标准误。NM代表原生草甸,LM代表轻度退化草甸,MM代表中度退化草甸,HM代表重度退化草甸,下同。
图1 高寒草甸不同退化阶段甲烷通量特征
2.2 退化演替过程中二氧化碳通量特征
同时期的二氧化碳通量观测结果显示,生长季通量高,而休眠季低,4种草甸生长季二氧化碳通量均显著高于休眠季通量(p<0.05)。原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸二氧化碳年平均通量分别为294.50,400.03,513.374,521.17 mg/(m2·h),四者差别较大(图2),但这种差异主要出现与生长季,休眠季4种不同退化阶段的草甸二氧化碳通量差异不显著(p>0.05)。
草甸二氧化碳排放能力随草甸退化程度加重而逐渐减少,其中重度退化高寒草甸相比原生高寒草甸二氧化碳通量减少76.9%。单因素方差分析显示,中度退化草甸和重度退化草甸二氧化碳通量显著低于原生草甸(p<0.05),但二者与轻度退化草甸相比差异不显著;原生草甸与轻度退化草甸甲烷排放能力无差异(p>0.05);中度退化草甸与重度退化草甸相比差异也不显著(p>0.05)。
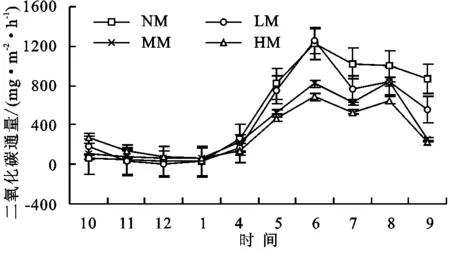
图2 高寒草甸不同退化阶段二氧化碳通量特征
2.3退化演替过程中氧化亚氮通量特征
同时期氧化亚氮通量观测值都为正值,说明高寒草甸是大气氧化亚氮的源,相比甲烷和二氧化碳其季节特征不明显(图3),而仅表现为在生长季末有一个峰值,对不同退化阶段草甸休眠季初期(10月、11月、12月)氧化亚氮通量进行配对T检验,结果显示此时间段重度退化高寒草甸氧化亚氮通量显著高于其余3种草甸(p<0.01)。休眠季氧化亚氮的通量对于草地退化的响应更加明显,总体来看草甸退化程度越严重其氧化亚氮排放速率越大。
原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸氧化亚氮通量分别为47.20,35.17,53.40,87.07 μg/( m2·h)。方差分析显示重度退化高寒草甸氧化亚氮排放能力显著高于原生草甸(p<0.05),而其余3种退化阶段草甸氧化亚氮通量差异不显著,重度退化草甸相比原生草甸氧化亚氮排放能力增加了84%。
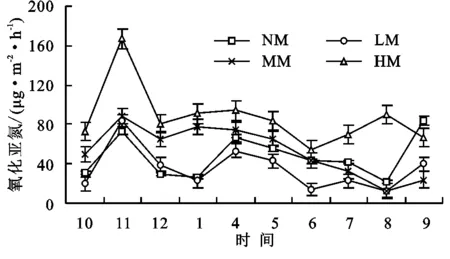
图3 高寒草甸不同退化阶段氧化亚氮通量特征
2.4 草甸生态系统要素对退化的响应
随着放牧强度的增强,高寒草甸主要植被特征参数(有机质、活根生物量、死根生物量、地上生物量、植被盖度)发生显著变化,土壤有机质含量、活根生物量、死根生物量总的变化趋势为先增加后减少,有机质含量与死根生物量有较好的对应关系,但是死根生物量在轻度退化阶段就表现出增加趋势,而活根生物量直到中度退化阶段才表现出增加(图4);高寒草甸生物量和植被盖度随着放牧强度的增加而减少,由原生草甸演)变到重度退化阶段其盖度减少幅度(52%±8.2%)大于生物量减少幅度(11.2%±4.3%)。
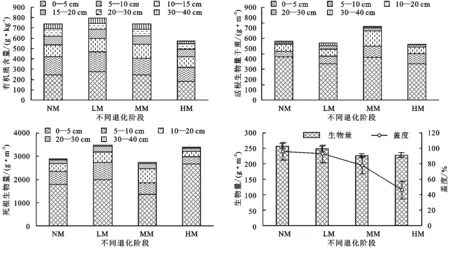
图4 植被特征对于退化的响应
随放牧强度的增高,高寒草甸环境因子也发生了明显变化,但变化幅度弱于植被特征参数变化幅度,pH变化较小,但表层pH值有所增大;紧实度随放牧大而显著增加(p<0.05),相比原生草甸重度退化草甸土壤紧实度增加26.7±3.6%;土壤营养状况也随着放牧强度增加而恶化,总氮和总磷含量显著下降(p<0.05),相比原生草甸重度退化草甸土壤总氮下降幅度(25.6%±3.2%)大于土壤总磷下降幅度(13.6%±1.3%)(图5)。

图5 土壤环境特征对于退化的响应
2.5 高寒草甸温室气体通量的关键要素筛选
通过相关分析,从环境要素中选取了有机质、5—10 cm活根、5—10 cm死根、0—5 cm全氮、10—15 cm全磷、生物量、植被盖度、20—30 cm紧实度和pH十个环境要素作为主要的影响因子,与温室气体进行逐步回归分析,结果显示不同的温室气体受控的关键影响因子不同,草地甲烷吸收量主要由土壤紧实度、有机质和盖度控制;草地二氧化碳通量主要受全磷、植被盖度和全氮控制,而草地氧化亚氮主要受土壤有机质、紧实度和死根生物量控制(表2)。

表2 高寒草甸温室气体通量的关键要素筛选
3 讨 论
3.1 放牧作用对草地植被与环境的影响
放牧活动显著改变草地植被、草地C,N元素周转活动。高寒草甸的土壤有机质C含量高,潜在肥力高,速效养分匮乏,草地碳周转对于草地生物多样性和稳定性具有重要影响,同时草地结构和功能的发挥也依赖于草地有机碳[6]。放牧活动显著减少土壤总C库、植物地下部分C库、土壤微生物C库和凋落物C库,分别减少10.28%,13.72%,21.62%和8.93%[17],土壤微生物C只占土壤有机C的1%~5%,但却是土壤有机质和养分的重要驱动者,同时也是综合评价土壤C动态、土壤质量和土壤肥力的重要指标[18]。放牧活动显著增加草原生态系统土壤呼吸通量、土壤净氮矿化和硝化速率,增幅分别为4.25%,30.63%和12.88%。土壤N素的固定和矿化、植物对N素的吸收及内部转移、分配、凋落物N素返还土壤、土壤N素硝化返还大气是草地生态系统中N素循环的主要途径。放牧家畜主要通过排泄、对凋落物的践踏作用及改变植物生理活动或群落结构3种途径影响着土壤的N素循环[19],但其具体的作用结果仍然存在争议。草地生态系统N 素99%以上分布在0—30 cm 的土层中,只有不到0.4%的N 存在于植物体中,并且重牧使牧草的再生能力降低,地上部分的N 素分配减少[20]。长时间的放牧活动不仅改变群落结构的同时也能显著减慢N素的循环速率[21],放牧作用显著减少土壤N库、植物地下部分N库、土壤微生物量N库和凋落物N库,减幅分别为13.38%,4.40%,24.40%和10.39%[17]。随着高寒草甸的退化,0—100 cm 土壤的有机C、全N、全P和全K含量以及碳氮比、碳磷比、碳钾比、氮磷比、氮钾比和磷钾比均呈降低趋势,速效养分(速效N、速效P、速效K)下降尤为明显[22],相对其他化学计量特征,土壤N素是放牧的第二大敏感要素[23],在中等放牧强度下草地系统N素输出就表现为大于系统的输入[3]。随着放牧强度增大凋落物逐渐减少,过度放牧严重阻碍草地物质平衡,重度放牧下以凋落物和家畜粪便返还草地仅只有20%~50%[24]。本研究与先前研究较为一致,放牧活动引起的草地退化使草地大量的C,N流失,其中速效养分损失最为严重,本研究还发现草甸生态系统要素变化与草地退化年限关系密切,通过对不同退化阶段植被要素和土壤要素的对比发现,草地土壤环境具有较大的稳定性,放牧活动引起的草地退化首先表现在植被特征参数的下降,随着退化年限的增加,才会影响到土壤环境,因此关于退化草甸温室气体通量研究应明确其退化年限。
3.2 草地温室气体通量与环境因子
本研究中不同退化阶段的高寒草甸甲烷吸收速率差异较大,原生高寒草甸、轻度退化高寒草甸、中度退化高寒草甸和重度退化高寒草甸甲烷通量分别为-40.09,-37.78,-45.06,-56.66 μg/(m2·h),这与先前关于高寒草甸、高寒草原、羊草草原、大针茅草原差别不大[16],但是本研究中重度退化草甸相比原生草甸甲烷通量增加41.33%,且中度和重度退化草甸甲烷通量要显著高于先前大多数类型生态系统,说明在草地退化过程中草地甲烷氧化能力是逐渐增加的,草地的甲烷吸收能力主要是来自元甲烷氧化菌的作用,草地退化过程中一些环境条件的改变,根据逐步回归分析可知,草甸甲烷吸收能力影响最为重要的因素是:土壤紧实度、有机质和盖度,而先前的研究对于草地影响因子的分析大多都是温度、水分、土壤孔隙[25],草地土壤在退化过程中温度变化不大,但水分逐渐降低,水分下降后通气性增加,因此可理解为先前大多数研究认为草地甲烷吸收能力随通气性的增加而增加[13,16,25],而本研究却发现紧实度、有机质、盖度对于草地甲烷通量影响最大,是对草地甲烷通量影响研究理论的一种补充。从原生草地退化为重度退化草甸,此过程有利于甲烷氧化菌的生长,但本研究未涉及极度退化高寒草甸,有研究显示草甸地上植被对于草地甲烷氧化菌具有重要保护作用,裸地甲烷氧化菌数量骤降其甲烷吸收能力也大大降低[26],有关研究还需要进一步进行。
CO2在大气中的寿命为5~200年,贡献了50%~60%的温室效应[27],工业革命以来的大气CO2浓度上升引起人们的强烈关注。生态系统呼吸包括自养呼吸和异养呼吸两部分,其中自然呼吸指的是植物本身的呼吸作用,异养呼吸是微生物的分解释放CO2。放牧对于草地植被和土壤的影响是显而易见的,总得来说其既有积极作用,又有消极作用[28],放牧通过家畜的选择采食、践踏和粪便归还而直接影响草地植物群落结构和土壤理化性质[29],由于草地植物超补偿生长的存在,轻度放牧能促进草地生长[30],随放牧强度的增大,优势种比例显著下降,植被生产力降低、群落地上、地下生物量显著下降,群落结构趋于简单化,引起一系列的草地生态问题[31]。本研究试验样地设置于2014年,不同的放牧活动已经对草地植被和草地土壤环境造成很大影响,但由于放牧时间较短,放牧活动对于草地产生的影响较多的反应在草地植被上(生物量、盖度大幅减少),而草地土壤环境的变化是由上而下,需要长期的放牧活动体现。本研究设置的4个处理样地(原生、轻度退化、中度退化、重度退化)的二氧化碳通量,只有重度退化草甸显著低于原生草甸,并未发现轻度退化草甸、中度退化草甸和重度退化草甸间二氧化碳通量差异,所以关于草地退化温室气体通量的研究,草地退化年限是很重要的,将短时期形成的退化草地代表整个青藏高原高寒草甸温室气体通量的计算方法,将会高估本底温室气体排放量。
本研究中所有退化阶段草地均排放N2O,但也有研究表明非生长季湿地土壤是N2O“汇”[32],本研究原生草甸、轻度退化草甸、中度退化草甸和重度退化草甸氧化亚氮通量分别为47.20,35.17,53.40,87.07 μg/(m2·h),其中重度退化草甸氧化亚氮通量显著高于其余3种草甸(p<0.05)。草地的N2O通量主要依赖于微生物的硝化和反硝化作用[33],草甸的退化演替过程中,由原生退化为轻度再到中度的过程草甸N2O通量变化并不明显,这种不明显的变化更加体现了草地退化过程中土壤环境相对于植被变化的迟滞性,而草地在持续的过渡放牧压力下,到达重度退化阶段后,N2O通量大幅增加,氧化亚氮排放能力相比原生草甸增加了84%。高寒草地生态系统土壤中有较高的根系密度和有机碳含量,其缓慢的分解周速率[34],形成了一个大气CO2汇的功能。但是近年来在人类活动和气候变化的双重干扰下,天然草地的碳汇作用被减弱甚至逆转为碳源[35]。本研究目前最有意义的发现就在于草地退化后期大量的草地封存碳通过温室气体排放,但草地碳流失具体格局还有待于进一步研究,虽然草地退化增强了草地大气甲烷汇功能,这种增汇依赖于甲烷氧化菌活性增加,当草地退化到一定年先后,失去植被保护的土壤系统中甲烷氧化菌数量和活性也会大大降低,而引起加完汇的丧失,草地退化使二氧化碳排放能力大大降低,其原因是由于植物生物量降低,植物的呼吸作用大大减少所致,但草地退化同时也伴随着土壤封存碳的大量流失,所以草地退化使其由大气碳汇变为大气碳源。因此现阶段探索高寒草地合理利用模式,是发挥其气候变化缓冲功能的重要途径。
4 结 论
在高寒草甸的退化演替过程中,3种主要的温室气体通量都大幅增加,相比原生草甸,重度退化草甸甲烷吸收能力增加41.33%,二氧化碳排放减少76.9%、氧化亚氮排放能力增加84%。放牧活动对于高寒草甸的影响首先表现在植被上,而土壤环境的变化比较迟滞,因此退化年限对于草地温室气体通量至关重要。通过逐步回归分析得知,草甸甲烷通量影响较大的环境因素为:土壤紧实度、有机质、盖度,二氧化碳通量影响较大的环境因素为:全磷、盖度、全氮,氧化亚氮通量影响较大的环境要素为:有机质、紧实度、死根,高寒草甸退化演替发展到重度退化阶段时释放大量温室气体。
猜你喜欢
儿童故事画报·自然探秘(2024年5期)2024-05-22 22:26:58
疯狂英语·新读写(2023年4期)2023-05-10 10:44:22
青海草业(2022年2期)2022-07-23 09:34:58
农业灾害研究(2022年1期)2022-05-07 01:31:04
天津农林科技(2022年2期)2022-04-19 10:48:20
中国特种设备安全(2021年11期)2021-05-05 06:13:20
中国化肥信息(2019年7期)2019-08-26 09:47:00
活力(2019年21期)2019-04-01 12:17:10
化工进展(2015年6期)2015-11-13 00:26:29
中国海洋大学学报(自然科学版)(2014年12期)2014-02-28 12:22:06
