
日粮因素对反刍动物胰腺外分泌功能的调控
2019-09-05 04:04:38姚军虎曹阳春
饲料工业 2019年16期
■姚军虎 郭 龙 曹阳春
(西北农林科技大学,陕西杨凌712100)
胰腺外分泌对日粮中营养物质在反刍动物小肠内的消化十分重要。胰腺外分泌部由分泌胰液的腺泡和运输胰液的胰腺管组成,能合成和分泌胰淀粉酶、脂肪酶、蛋白酶和糜蛋白酶等消化酶,能够消化过瘤胃的淀粉、脂肪和蛋白质等营养物质。国内外研究中,以胰腺淀粉酶的分泌调控居多,因为反刍动物小肠内过瘤胃淀粉的消化受到胰腺外分泌功能的限制[1-2],约有40%的过瘤胃淀粉不能被消化利用[3],造成能量利用效率偏低。所以提高反刍动物胰腺外分泌能力,尤其是淀粉酶的分泌,是提高能量利用效率的关键[4]。
1 反刍动物胰腺的生理结构
胰腺作为化学性消化腺体,由分泌胰岛素和胰高血糖素等激素的内分泌部和分泌淀粉酶、蛋白酶和脂肪酶等消化酶的外分泌部组成,在动物正常生长发育、繁殖和生产过程中发挥重要的作用。奶牛对淀粉等日粮营养物质的消化主要依赖瘤胃的微生物消化,肠道内的化学性消化则与单胃动物相似,主要依赖胰腺分泌的消化酶。认识反刍动物胰腺的生理结构及分泌消化酶的主要细胞类型——腺泡细胞,是进行胰腺外分泌功能的调控研究的基础。
1.1 胰腺的生理结构
胰腺在奶牛体内位于腹膜后,呈长条状,分为头、颈、体、尾4种结构,但各结构间并无明显界限。胰腺表面被薄层结缔组织所覆盖,被十二指肠环绕。胰腺按照功能分为内分泌部和外分泌部。胰腺外分泌部分呈多分支小叶状,包含分泌胰酶的腺泡和分泌水、电解质并具有运输通道作用的导管。胰液由碳酸氢钠等无机盐、淀粉酶、蛋白酶等酶原颗粒和水构成,通过导管流入十二指肠内,消化食糜中的淀粉、蛋白质和脂肪等物质。胰腺内分泌部分呈紧密球团状,嵌于外分泌部分,由胰岛细胞构成。胰岛细胞分为四类,包括α细胞、β细胞、δ细胞和PP细胞,分别分泌升高血糖的胰高血糖素、降低血糖的胰岛素、生长抑素(以旁分泌的方式抑制α、β细胞)和胰多肽(抑制胃肠运动、胰液分泌和胆囊收缩)。内分泌细胞只占胰腺细胞总数的极少部分[5],但分泌的激素却有十分重要的调节功能,尤其是在血糖平衡方面。
1.2 胰腺腺泡细胞
胰腺组织中,80%的细胞是腺泡细胞。腺泡是胰腺行使外分泌功能的基本功能结构,由锥形的浆液性腺细胞组成,主要用于合成、储存和分泌各种消化酶。腺泡细胞是典型的分泌型细胞,其结构如1 图所示。大量的酶原颗粒分布于细胞基部,苏木精染色呈嗜酸性,颗粒数量随功能和状态而变化。酶原颗粒旁边有许多核糖体和内质网等细胞器,呈嗜碱性,细胞核呈椭圆形,位于细胞中部偏上。内质网核糖体合成蛋白质,运输到高尔基体加工后形成酶原分泌颗粒,腺泡细胞以胞吐的方式将酶原颗粒释放到腺泡腔,再经过胰腺导管进入十二指肠,被肠激酶活化后发挥营养物质消化的作用。本课题组成功分离并培养了奶牛胰腺腺泡细胞,并以此为模型,研究了功能性氨基酸对消化酶分泌的影响和机理。
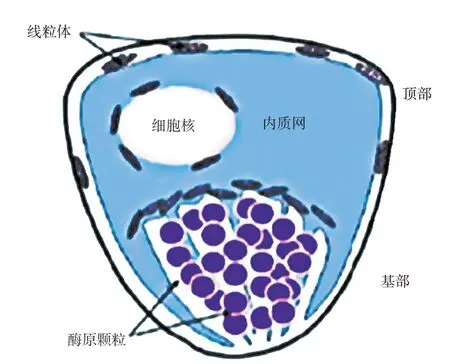
图1 胰腺腺泡细胞的结构示意图[6]
2 反刍动物对碳水化合物的消化过程
反刍动物瘤胃内存在大量的细菌、原虫和厌氧真菌,尽管没有消化腺体,但可依靠这些种类和数量庞大的微生物发酵和降解摄入的饲料。皱胃具有消化腺体,能够分泌消化液,但是消化能力有限。反刍动物,特别是奶牛,瘤胃内碳水化合物的消化代谢调控研究的较为透彻,目前,单纯调控瘤胃的消化代谢并不能继续提高动物的生产性能。小肠内的化学性消化逐渐受到重视,其消化生理的研究围绕胰腺功能、小肠酶分泌、小肠微生物、小肠葡萄糖转运载体以及小肠屏障功能等的作用及其调控。反刍动物可高效的利用蛋白类和脂肪类饲料进行生长、生产、繁殖等[7]。
奶牛日粮中的纤维素和半纤维素在瘤胃内由微生物产生的纤维素酶降解,分别产生葡萄糖和戊糖,进而生成丙酮酸和挥发性脂肪酸(VFA),瘤胃微生物不能消化木质素。淀粉等非结构性碳水化合物被瘤胃微生物发酵产生乙酸、丙酸、丁酸等VFA,经瘤胃壁吸收后为奶牛提供能量,同时被微生物自身利用产生甲烷,损失能量[8],降低能量利用效率。微生物发酵产热时也会消耗能量。研究发现,瘤胃微生物发酵淀粉产热的能量损失约为日粮消化能的3%~12%[9]。瘤胃内保持一定比例的淀粉发酵,有利于反刍动物瘤胃能氮平衡,获得更高的微生物产量。
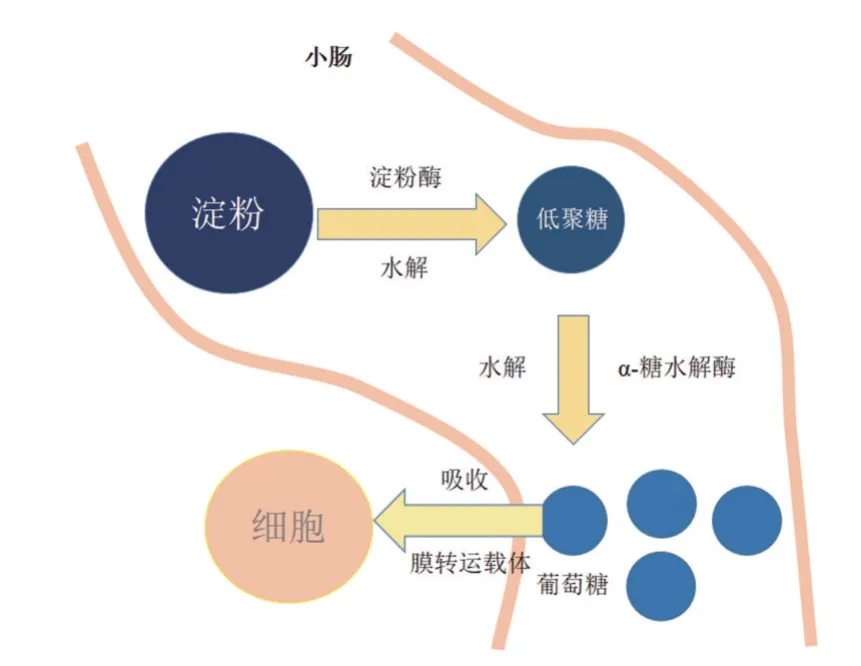
图2 淀粉在反刍动物小肠内的消化吸收过程
淀粉类饲料是日粮中主要为奶牛提供能量的营养成分,大部分在瘤胃中被微生物发酵生成VFA被瘤胃壁吸收,在肝脏内合成葡萄糖或在乳腺内合成乳脂。小肠内的消化过程,反刍动物与单胃动物相似,具体的消化过程如图2 所示,需要经历以下三个过程:①在淀粉酶的作用下淀粉被水解成小分子低聚糖;②在α糖水解酶的作用下,低聚糖被分解为葡萄糖;③葡萄糖转运载体将肠腔中的葡萄糖转运到小肠上皮细胞内[10]。由此看出,淀粉在小肠内的消化起始于胰腺腺泡细胞分泌的淀粉酶[11],淀粉酶在肠腔中被活化,切断α-1,4 糖苷键,将淀粉分解成葡萄糖分子或小分子低聚糖(如麦芽糖、麦芽三糖等)从而被肠上皮细胞吸收,为机体供能。真胃灌注淀粉比瘤胃提供的代谢能转化为组织能量的效率高25%[12]。小肠淀粉消化过程中没有发酵产热,但在消化酶的作用下,糖苷键断裂也释放热能,Baldwina 等[13]估计淀粉在小肠消化中释放的热量相当于日粮消化能的0.6%,远远小于瘤胃中的能量损失。
反刍动物摄入的淀粉中有15%~25%不经瘤胃消化进入小肠[4]。尽管理论上淀粉可在小肠内被消化,并以葡萄糖的形式被高效吸收、直接供能,效率远高于瘤胃内微生物降解产生VFA 吸收到体内被合成葡萄糖等能量物质的效率,但反刍动物小肠淀粉消化能力有限,仅能够消化40%~62%的过瘤胃淀粉[1],限制了淀粉在小肠中的消化利用。提高日粮淀粉水平虽短期内可增加奶牛的有效能供应和生产性能,但过高的瘤胃可发酵淀粉往往诱发各种代谢性疾病,如瘤胃酸中毒。同时,过多的过瘤胃淀粉却因胰腺淀粉酶分泌不足[10]而难以在小肠中被完全消化,降低日粮能量效率甚至引起后消化道酸中毒。因此,改善反刍动物胰腺淀粉酶的分泌能力成为提高小肠淀粉消化的关键。
3 胰腺外分泌功能的影响因素
胰腺消化酶的分泌在营养物质的摄入和消化时启动,同时受到多种因素影响,例如动物年龄、神经和体液、日粮组分等。
3.1 动物年龄
随着日龄的增加,动物胰腺的发育逐渐成熟,分泌胰液的总量及其内含有的消化酶的量提高[14]。新生犊牛小肠内容物中的消化酶包括淀粉酶、蛋白酶和脂肪酶活力都很低,60日龄时,这些酶活力显著提高,淀粉酶活力比初生时约高40倍[15]。胰液总量的分泌情况与之类似,4 日龄犊牛分泌胰液的流量仅为100 日龄犊牛的1/6[16]。
3.2 神经和体液
中枢神经系统通过迷走神经调控胰腺分泌功能。阻断迷走神经后,降低犊牛胰液的分泌量、胰蛋白酶的活性和胰液中蛋白质的浓度[17-18]。此外,刺激胆碱能神经可使胰腺外分泌增加,且提高内源性胰泌素和胆囊收缩素的分泌。这些试验结果说明神经在调节胰腺的外分泌功能中具有十分重要的作用。
胆囊收缩素(cholecystokinin, CCK)已被证实是调控胰酶释放的重要脑-肠肽。CCK 由肠内黏膜I 型细胞在受到氨基酸和脂肪酸的刺激时产生和分泌。CCK通过两种方式调控胰腺外分泌功能:①直接作用于腺泡细胞膜表面受体[19],此过程会受CCK颉颃剂影响(阿托品除外);②与胆碱能神经元调节器互作,调控胰腺腺泡细胞的分泌能力。CCK 受体有CCK1 和CCK2两个亚型,均属于G蛋白偶联受体家族[20-21]。对犊牛进行CCK受体定位,发现这两种受体均在胰腺中表达[22],但成年后胰腺内的CCK受体主要为CCK2,且主要分布于胰岛细胞及导管细胞膜表面,而腺泡细胞膜并没有这两种受体的表达。主要是因为随着犊牛的发育,胰腺功能出现转化和退化,丧失一些功能。此外,免疫荧光试验的抗体选择和成像方法,对试验结果的影响也不容忽视。CCK 还能够促进小鼠胰腺的发育,表现为提高胰腺重量、腺泡细胞体积、胞内核酸和蛋白质的含量[23]。CCK 的体循环水平影响小鼠胰腺酶基因的表达[24]。在反刍动物上的研究发现,当用CCK类似物(如蛙皮素)孵育牛胰腺组织时,会直接影响胰腺外分泌功能,这种影响具有动态性且只出现在经过大量灌注过瘤胃蛋白质的牛的胰腺组织上[25]。在苯丙氨酸和CCK 互作促进奶牛胰腺腺泡细胞消化酶分泌的试验中,我们也观测到CCK对腺泡细胞分泌能力的直接影响。
胰岛素是胰岛β细胞分泌的激素,能调节机体的血糖浓度,并作为信号分子参与机体其他的生理代谢过程,如促进mTOR信号通路调控蛋白质的合成。胰岛素调控血糖浓度是通过促进组织细胞摄取和利用葡萄糖,并抑制糖原分解和糖异生过程来实现的。胰岛素调控单胃动物和反刍动物胰腺外分泌功能的作用可能存在差异。胰岛素能改善糖尿病小鼠胰腺的外分泌功能[26],但是奶山羊十二指肠灌注亮氨酸后胰腺α-淀粉酶mRNA 表达的调控不依赖于血清胰岛素的变化[27]。
3.3 日粮因素
3.3.1 碳水化合物
第三,着力打造农产品物流平台,提升物流配送保鲜能力。河北省逐步建立多层次、宽领域、全方面的网格化物流体系。有效开展农产品第一、二、三方物流,形成单独的河北特色农产品物流运送体系,线上与线下相结合,可通过专业线上互联网平台,搭建新型物流运送体系,进行会员跟踪物流配送制度,以便更好了解用户所需。
日粮碳水化合物的比例和类型影响胰腺的外分泌功能,尤其是淀粉酶的分泌。绵羊饲喂以粉碎玉米为基础的日粮时较饲喂干草提高了胰腺淀粉酶的浓度和活力[28]。同时,研究发现奶山羊小肠干物质和淀粉消化率分别与食糜淀粉酶存在显著的线性关系(图3),说明淀粉酶在碳水化合物和干物质消化中具有重要作用[29]。由表1 可知,奶山羊饲喂淀粉含量分别为25%、35%和45%的等蛋白质日粮时,高淀粉含量组(45%淀粉组)的胰腺淀粉酶活力显著高于25%淀粉组[30]。也有研究表明,单纯增加过瘤胃淀粉并不能够改善反刍动物胰腺的外分泌功能或影响效果会随时间及剂量效应而消失。荷斯坦阉牛皱胃灌注水解淀粉(20、34.2 g/h 和40 g/h)时降低了α-淀粉酶浓度及分泌速率[31-32]。奶山羊胰腺淀粉酶分泌随着过瘤胃淀粉的增加逐渐增加,在102 g/d达到峰值,随后也出现了下降[33]。这些结果说明小肠碳水化合物的含量影响淀粉酶的分泌,但是这些或正向或负向的效应是由肠腔中碳水化合物的增多还是吸收的葡萄糖浓度升高而引起的,目前还不得而知。因此,通过增加过瘤胃淀粉含量的方法提高小肠内淀粉的消化率目前还并不能够取得理想的效果,应该探索其他的营养策略例如调整日粮氨基酸比例和蛋白含量,调节激素(胰岛素)的分泌等。
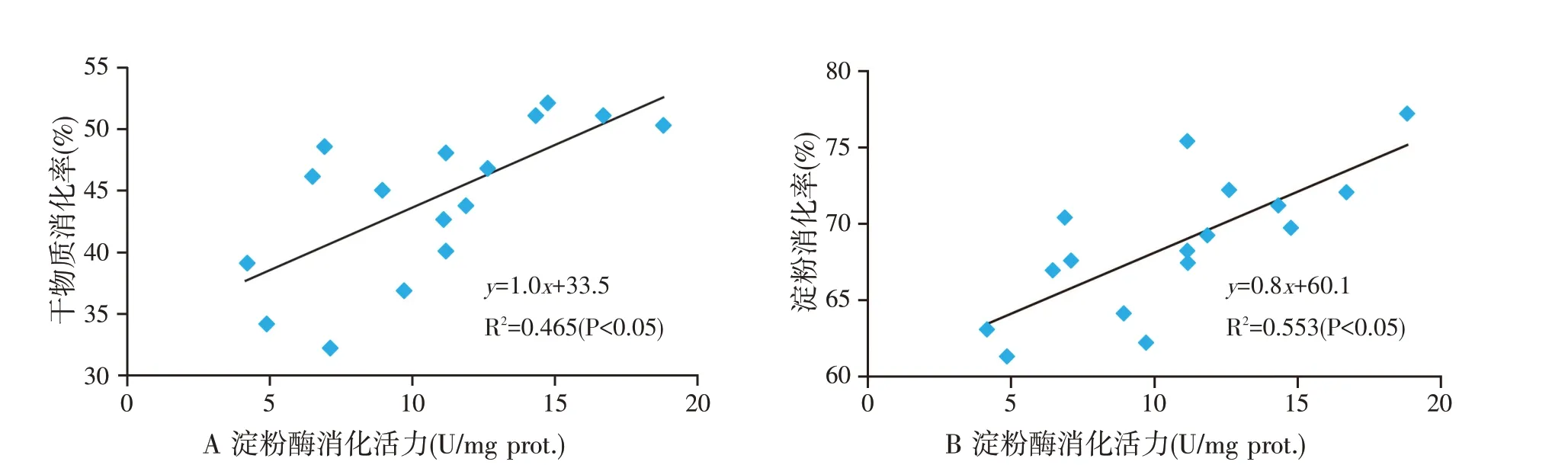
图3 山羊小肠食糜淀粉酶活力与日粮干物质(A)和淀粉(B)小肠消化率间的关系[29]
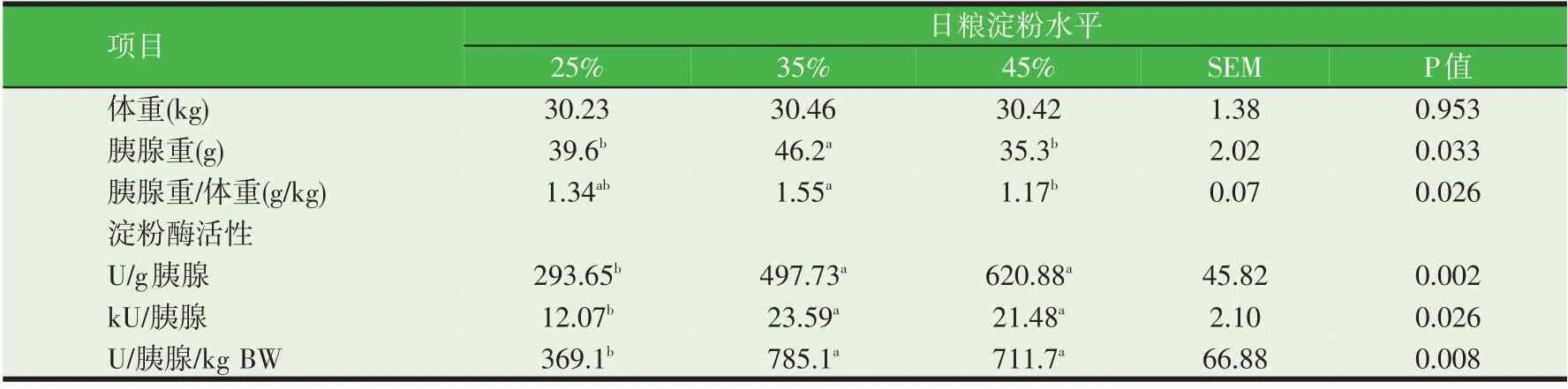
表1 饲喂不同比例淀粉对山羊胰腺淀粉酶活力的影响
3.3.2 氨基酸(蛋白质)
功能性氨基酸尤其是亮氨酸、异亮氨酸和苯丙氨酸,在刺激反刍动物消化酶分泌的试验中已经表现出显著的结果。
奶山羊十二指肠连续灌注亮氨酸14 d 后,线性增加了淀粉酶的活力,但并不影响总蛋白质输出、脂肪酶和胰蛋白酶分泌及血浆胰岛素浓度;胰液和淀粉酶分泌符合二次曲线的规律,最高值出现在亮氨酸灌注3 g/d 和6 g/d;奶山羊十二指肠亮氨酸灌注剂量和时间显著影响了胰酶的分泌,但是不影响血浆胰岛素的浓度[27]。十二指肠灌注亮氨酸也能够显著调控奶山羊胰腺的发育和胰酶基因的表达,对反刍动物肠道内淀粉等营养物质的消化具有重要作用[34]。Liu 等[35]利用4 头荷斯坦青年牛[(215±7)kg]进行胰腺插管及十二指肠瘘管,灌注不同浓度的亮氨酸,结果发现10 g 亮氨酸显著提高了胰液的分泌、淀粉酶的浓度和分泌速率。荷斯坦奶公犊牛饲喂原奶中添加亮氨酸,发现胰腺蛋白质的浓度(mg/kg BDW)显著的增加了,但是并没有影响淀粉酶的分泌[36],血清葡萄糖,必需氨基酸、8 周龄体躯指数和平均日增重均显著提高[37]。亮氨酸可以作为营养信号刺激淀粉酶、胰蛋白酶、胰凝乳蛋白酶、脂肪酶的分泌,且亮氨酸的这些功能有剂量和时间依赖性。
异亮氨酸和亮氨酸同属支链氨基酸,在分子结构和功能上与亮氨酸类似[38]。荷斯坦青年奶牛十二指肠连续灌注试验表明,灌注异亮氨酸10 h 时,胰液的分泌量和pH 值没有受到影响,但是胰液中蛋白质浓度(mg/ml)、胰液分泌速率(mg/h)、淀粉酶活力、胰蛋白酶活力在20 g 和30 g 异亮氨酸灌注组显著提高,且符合线性规律,糜蛋白酶和脂肪酶活力不受异亮氨酸灌注量的影响(表2);灌注异亮氨酸10 d 后,胰腺总蛋白分泌量、胰蛋白酶分泌浓度和分泌速率与异亮氨酸灌注浓度成线性正相关,胰液总蛋白浓度和分泌速率、淀粉酶活性显著提高,糜蛋白酶和脂肪酶活力不受影响(表3)[39]。在体外孵育奶山羊胰腺组织的试验中,我们发现异亮氨酸通过提高4EBP1γ的磷酸化程度,降低eEF2 的磷酸化水平提高淀粉酶、胰蛋白酶和糜蛋白酶的组织浓度、缓冲液释放浓度。
苯丙氨酸属于芳香族氨基酸,有研究表明可以通过CCK 途径调控胰腺的外分泌[40]。奶山羊十二指肠连续灌注苯丙氨酸14 d的试验发现,奶山羊胰胆汁分泌量、淀粉酶分泌量和血清CCK 浓度随苯丙氨酸灌注量的增加呈现先增加后下降的二次曲线变化,在2 g/d 苯丙氨酸时有最大值;胰蛋白酶分泌也呈二次曲线变化,最大值出现在4 g/d;脂肪酶分泌受苯丙氨酸影响显著降低[41]。奶公犊饲喂原奶中添加亮氨酸和苯丙氨酸对小肠食糜α-淀粉酶(表4)活力无显著影响(P>0.05),但小肠各位点食糜内淀粉酶活力有显著差异(P<0.05),各处理和小肠相对位点间存在交互作用的趋势(P=0.054);亮氨酸和苯丙氨酸处理组小肠α-淀粉酶从0%到100%位点,呈先下降再上升的变化趋势;苯丙氨酸对奶公犊小肠食糜胰蛋白酶活力无显著影响(P>0.05),相对位点不同显著影响胰蛋白酶活力(P<0.05)(表5)[36-37]。此外,体外试验表明,苯丙氨酸显著提高奶牛胰腺组织淀粉酶、胰蛋白酶的合成、分泌及mRNA表达[42]。

表2 十二指肠灌注异亮氨酸10 h对奶牛胰腺外分泌的影响
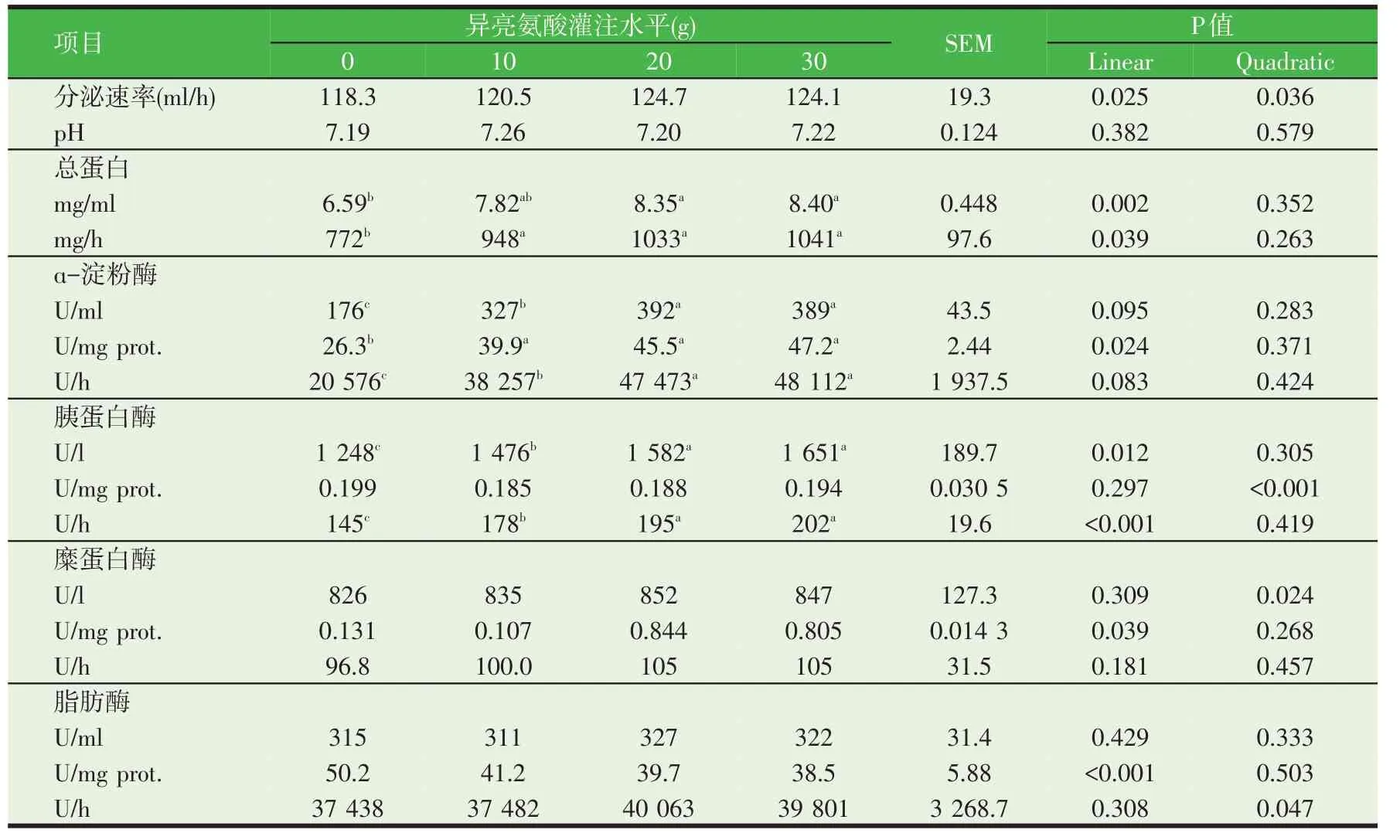
表3 十二指肠灌注异亮氨酸10 d对奶牛胰腺外分泌的影响
4 胰腺外分泌调控的机理
本课题组以奶公犊牛原代胰腺腺泡细胞为试验模型,对其外分泌的调控机理进行了研究,发现胞内钙离子、蛋白酶体及相关信号传导途径等构成的复杂调控网络是胰腺腺泡消化酶合成和分泌的关键。
4.1 胞内钙离子信号
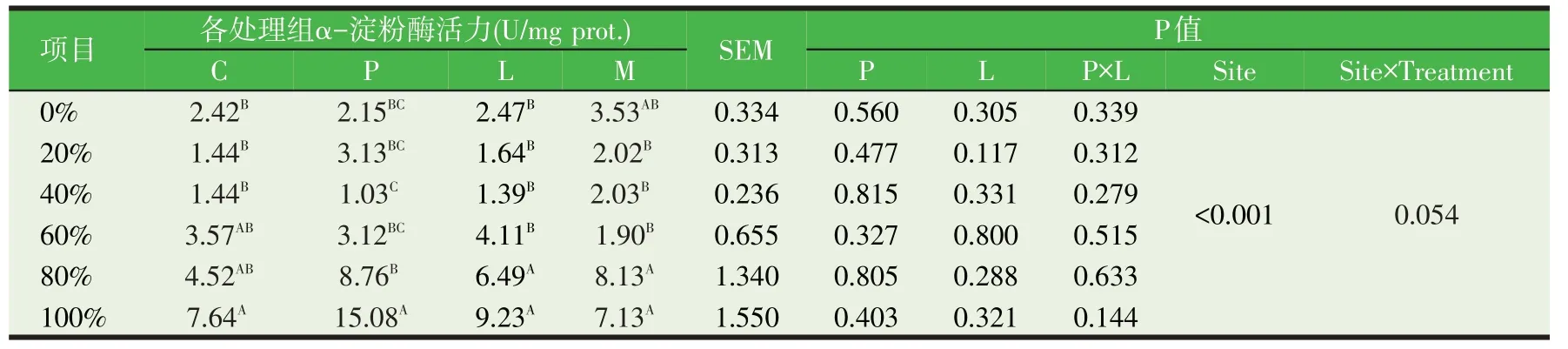
表4 原奶中添加亮氨酸和苯丙氨酸对奶公犊小肠食糜α-淀粉酶活力的影响

表5 原奶中添加亮氨酸和苯丙氨酸对奶公犊小肠食糜胰蛋白酶活力的影响
钙离子在腺泡细胞主要储存于内质网。在细胞质内,钙离子则发挥由刺激到分泌的偶联中心的作用。当细胞内CCK 或乙酰胆碱类激素达到生理刺激浓度时,细胞便会产生IP3和烟酸腺嘌呤二核苷酸磷酸(nicotinic acid adenine dinucleotide Phos Phate,NAADP)这两种第二信使,当它们与位于内质网上的利阿诺定受体(ryanodine recePtors,RyRs)和三磷酸肌醇受体(inositol 1,4,5-tris Phos Phate receptor,IP3R)结合后,会释放储存于内质网的钙离子,使细胞质内的钙离子浓度出现小幅度震荡,从而提高酶原颗粒从胞内到胞外的释放[47-48]。
4.2 蛋白酶体活性
蛋白酶体是细胞内重要的蛋白质复合体,起降解细胞内蛋白质的功能[49]。一些研究表明,在小鼠胰腺中,26S 蛋白酶体的亚基抗分泌因子(anti-factor 4)能够显著调控消化酶的分泌[50]。超生理浓度的亮氨酸(1.35 mmol/l)显著抑制蛋白酶体的活性,降低了奶牛胰腺腺泡细胞淀粉酶的分泌,通过使用蛋白酶体的特异性抑制剂MG132 后,发现蛋白酶体在该腺泡细胞中,对淀粉酶分泌起到重要的作用[51]。此外,本课题组通过Label-free 蛋白质组学分析方法,也发现亮氨酸显著影响了腺泡细胞内蛋白酶体相关蛋白亚基的表达。此外,在胰腺β型细胞中,胰岛素分泌受到蛋白酶体活性的影响[52]。泛素-蛋白酶体是细胞生命进程的重要调节方式,在疾病模型细胞内的功能和作用受到了广泛的关注,但是对于蛋白质代谢活动旺盛的分泌型细胞,还需要进一步研究。
4.3 分子信号途径
哺乳动物雷帕霉素靶蛋白信号(mammals target of rapamycin, mTOR),其上游的磷脂酰肌醇3-激酶(phosphatidylinositol-3,PI3K)信号及一般氨基酸调控阻遏蛋白2(generalamino acid control non-derepressible 2, GCN2)信号通路构成的分子信号网络主要调控腺泡细胞内氨基酸依赖的蛋白质合成过程[53]。mTOR 信号通路是调控细胞内蛋白质合成和降解、细胞生长和增殖的中心[54]。亮氨酸[55]和苯丙氨酸[42]能够分别激活Akt-mTOR 和S6K1,提高翻译起始复合物的形成而增加消化酶的合成。异亮氨酸也能够使mTOR 信号通路因子磷酸化,提高蛋白质合成的效率。氨基酸调控腺泡细胞内蛋白质周转和代谢的信号传导途径,已经有详细综述[56],具体调控网络见图4。
一般分泌型(SEC)信号通路是调控蛋白质运输的信号通路,而且广泛存在于各类型细胞中[57]。调控SEC系统的关键蛋白复合物,原核生物中是SEC YEG通道蛋白,真核生物中则是SEC61复合物[58]。我们发现,奶公犊腺泡细胞内高浓度的ATP,不仅能够供给蛋白质内质网加工的能量,提高蛋白质的合成,同时能够刺激SEC信号通路活化,促进酶蛋白从胞内释放到胞外,提高胰酶的分泌。氨基酸,尤其是亮氨酸介导的ATP增加,主要是依赖三羧酸循环和氧化磷酸化中关键酶活力的增加,如柠檬酸合成酶、异柠檬酸脱氢酶和细胞色素氧化酶等。
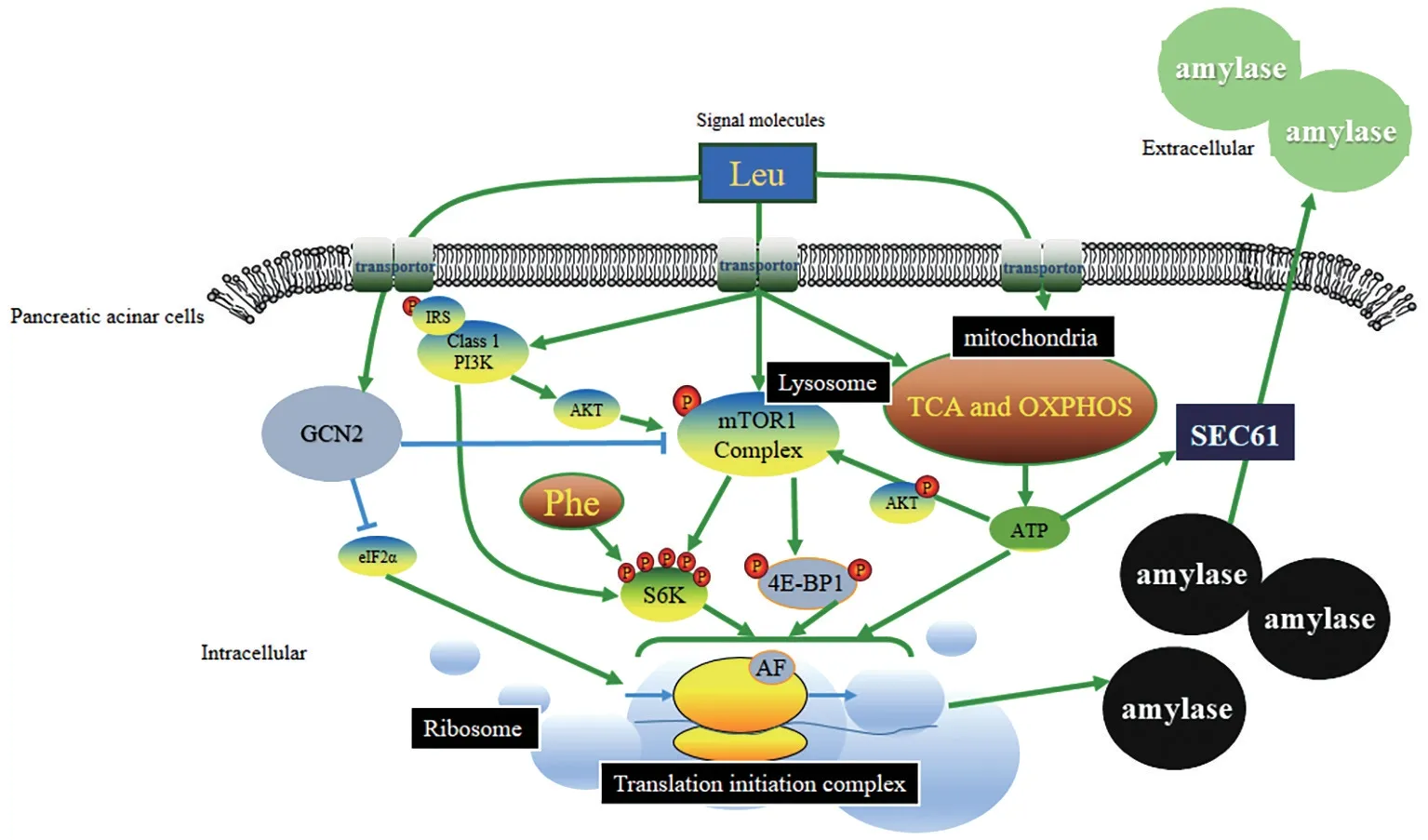
图4 氨基酸调控奶牛胰腺腺泡细胞消化酶分泌的信号传导网络
5 结语
反刍动物,尤其是奶牛,胰腺分泌淀粉酶不足是造成小肠淀粉消化率低的关键因素。理论上小肠内葡萄糖形式的供能效率优于瘤胃内挥发性脂肪酸的供能效率,在奶牛泌乳初期采食量低的情况下,泌乳需要大量能量供应,此时若以小肠供能为主要方式不仅可以提高奶牛的泌乳性能,也能够保证奶牛的健康。因此,通过营养手段提高胰腺外分泌能力,不仅可以解决奶牛能量供应问题,也可提高生产性能,促进机体健康,提高饲料效率,降低养殖过程中对环境的污染,促进奶业的可持续发展。
我们应注意到,奶牛等反刍动物的胰腺外分泌能力受众多因素影响,虽然应用改变日粮碳水化合物结构和单一功能性氨基酸的添加等方法取得了一定的效果,但是,氨基酸的平衡性及功能性氨基酸整合的作用还需要进一步研究。日粮外源消化酶的添加也能够提高肠道营养物质的消化,但酶的种类、数量等基础指标还需要进一步确定。肠道微生物在营养物质的消化过程中也具有重要的作用,我们不能忽视这些微生物的作用,对于微生物在肠道内的定植、代谢需要深入研究。
猜你喜欢
中华胰腺病杂志(2023年5期)2023-10-27 07:32:10
临床与实验病理学杂志(2022年11期)2022-02-15 16:20:11
科学与财富(2018年8期)2018-05-09 01:30:10
中华胰腺病杂志(2015年5期)2015-12-08 12:18:13
中华胰腺病杂志(2015年5期)2015-12-08 12:18:08
动物营养学报(2015年10期)2015-12-01 02:26:21
实用器官移植电子杂志(2015年5期)2015-04-03 03:10:46
大连工业大学学报(2014年2期)2014-09-19 08:52:52
中国中西医结合外科杂志(2013年3期)2013-03-11 20:04:59
食品科学(2013年19期)2013-03-11 18:27:35
