
亚热带人工针叶林叶绿素荧光参数日变化及其与植被生产力的关系
2019-09-04 09:30陈敬华王绍强王辉民杨风亭居为民
生态学报 2019年15期
陈敬华,王绍强,*,李 悦,王辉民,杨风亭,居为民,张 乾,梁 琛
1 中国科学院地理科学与资源研究所,生态系统网络观测与模拟重点实验室,北京 100101 2 中国科学院大学资源与环境学院,北京 100049 3 南京大学国际地球系统科学研究所,南京 210023 4 中国科学院地理科学与资源研究所,陆地表层格局与模拟重点实验室,北京 100101
植物光合作用是生物圈最基础的物质和能量来源,是决定植物生产力的重要过程,也是全球碳循环研究中的关键。被植物叶片中叶绿素分子吸收的光能以3种形式转化利用:1)驱动光合作用(光化学);2)以热量的形式重新发射;3)以荧光的形式重新发射[1]。这三种形式存在此消彼长的竞争关系,三者在被吸收的光能总和中所占比例之和为1,因此叶绿素荧光产量的变化包含了光合作用和热耗散的有效信息。由光合作用引起的叶绿素荧光产量的下降(淬灭)称之为光化学淬灭(Photochemical quenching),而由热耗散引起的则为非光化学淬灭(Non-photochemical quenching),二者均可以通过相应的叶绿素荧光参数反映。与"表观性"的气体交换指标相比,叶绿素荧光参数更具有"内在性"特点,被称为测定植物光合功能响应环境变化快速、灵敏和无损伤的探针[2-3],已经成为当前植物光合能力测定的重要技术手段之一[1,4]。
利用调制技术和饱和脉冲技术,脉冲振幅调制式(Pulse Amplitude Modulation, PAM)叶绿素荧光仪实现了叶绿素荧光的原位测量,使得光化学淬灭和非光化学淬灭的测量成为可能[5]。叶绿素荧光仪通过直接测量暗适应下的最小荧光Fo、暗适应下的最大荧光Fm以及光适应下的最小荧光Fo′、光适应下的最大荧光Fm′和瞬时荧光Ft,进而推导出与光化学反应相关的光系统Ⅱ(Photosystem Ⅱ,PSⅡ)最大光化学量子产量Fv/Fm、实际光化学效率ΦPSⅡ、光化学淬灭系数qP以及表征非光化学淬灭的荧光参数NPQ和qN等[6]。在众多荧光参数中,ΦPSⅡ可以衡量与PSⅡ反应中心相关的叶绿素将吸收的光能用于光化学反应(光合作用)的比例,能更准确反映实际的PSⅡ中心进行光化学反应的效率情况,可以作为光合作用的指标;NPQ表示吸收的光能用于热耗散的比例,是表征非光化学淬灭的一种量化形式,能更准确反映植物非光化学淬灭的情况,是植物耗散过剩光能为热的能力(即光保护的能力)的指标[3,7]。通过分析关键荧光参数的动态变化,可以检测植物光合能力的状态和变化,揭示植物光合作用机理。
由于叶绿素荧光技术在叶片光合作用过程中光系统对光能的吸收、传递、耗散、分配等方面的研究具有独特的作用,已经广泛应用于植物光合机制、逆境生理和生态适应性评价等诸多领域[8-11]。但是,我国当前开展的叶绿素荧光研究多是利用控制实验探讨植被对环境胁迫的响应和抗逆性[12-14],少量自然条件下的植被叶绿素荧光动态变化研究也多针对某类植物的特定生长时期[15-17],而利用叶绿素荧光开展针叶树种生理生态特征变化及其受环境影响的研究相对较少。
植物通过光合作用吸收大气CO2并固定在植被与土壤中形成碳汇,植被总固碳速率可以由总初级生产力(Gross primary productivity,GPP)表征[18]。GPP代表植物在单位时间内通过光合作用吸收太阳能产生有机物的总量[19],而光能利用率(Light use efficiency,LUE)被定义为植物将吸收的光合有效辐射通过光合作用转化有机干物质的效率[20],二者都是碳循环研究中的关键参数。我国南方亚热带人工林是东亚季风区碳汇的重要组成部分[21],位于江西泰和的千烟洲人工针叶林是我国典型的亚热带人工林生态系统,其年净碳吸收可达387.2 g C m-2a-1,是重要的碳汇[22]。但当前多是基于涡度相关通量数据开展的生态系统水平的碳循环研究,缺乏涉及植被叶片水平光合机制的研究,叶绿素荧光技术可以弥补这一空缺。引进种湿地松和本地种马尾松是我国两种重要的人工林类型,占我国中部和南部造林面积的59%[23]。因此,本研究以千烟洲亚热带人工针叶林为研究对象,利用2016年不同季节的叶绿素荧光测量和涡度相关通量观测数据,结合增强回归树(Boosted regression trees, BRT)和相关分析的方法,明确亚热带人工针叶林湿地松和马尾松叶片关键荧光参数不同季节的日变化特征,并分析和量化其受环境因子的影响,同时揭示关键荧光参数与植被总初级生产力、光能利用率的关系,为深入认识亚热带人工针叶林光合作用的过程机理提供支持。
1 材料与方法
1.1 研究区概况
研究区位于中国科学院江西省泰和县千烟洲生态试验站内,紧邻通量观测塔(26°44′48′′N,115°04′13′′E,110.8 m)。研究区气候属于典型的亚热带季风气候,太阳年总辐射量4661 MJ /m2,年均气温为17.9℃,年均降雨量为1485.1 mm,年均相对湿度84%[22,24]。降雨季节分配不均,雨季主要集中在3—6月份,夏季炎热少雨[25]。研究区内通量塔附近森林覆盖率高达90%,林分为林龄30年左右的人工针叶林,优势树种为湿地松(Pinuselliotii)、马尾松(Pinusmassoniana)以及杉木(Cunninghamialanceolata),树高约15.5 m[26]。
1.2 数据观测与处理
1.2.1叶绿素荧光参数的测定
选取研究区通量塔附近的优势树种湿地松和马尾松作为观测对象,分别于2016年春夏秋冬四个季节晴朗的天气(春:3月29日—4月1日;夏:6月4—9日;秋:9月21—26日;冬:11月10—15日),利用超便携式调制叶绿素荧光仪MINI-PAM-II(德国WALZ)测定湿地松和马尾松叶片叶绿素荧光参数日变化。在测量叶绿素荧光参数的同时,同步观测光合有效辐射(Photosynthetically active radiation,PAR)、叶片温度(Leaf temperature,Tleaf)以及相对湿度(Relative humidity,RH)。观测期间每天在通量塔附近选取湿地松或马尾松冠层高度自然生长的叶片3—5片,先对叶片进行遮光处理30 min,测定暗适应条件下的荧光参数(Fo、Fm),然后自然光条件下进行完全的光适应后,从8:00至17:00,每隔30 min测定光适应条件下的荧光参数(Fo′、Fm′和Ft)日变化,每个树种连续观测3天作为重复。为了量化吸收光能中的光化学利用和非光化学耗散,本研究选用实际光化学效率ΦPSⅡ和非光化学淬灭NPQ。二者的计算公式如下:
(1)
(2)
1.2.2通量数据采集与处理
研究区内针叶林的GPP和LUE由开路涡度相关通量观测系统(Open Path Eddy Covariance, OPEC)提供的通量观测数据计算获得。该系统安装于观测样地紧邻的通量塔上高度23 m处,由三维超声风速仪(CAST3, Campell Scientific Ltd., USA)和快速响应红外CO2/H2O分析仪(Li- 7500, LiCor Inc., USA)组成,数据采集器(CR5000,Campbell Scientific,USA)以10 Hz的频率进行数据采集,并在线计算为30 min的平均通量数据存储。此外,通量塔上安装有常规气象观测仪器,可以提供包括总辐射、光合有效辐射、空气温湿度和降水量等常规气象要素,同样为半小时平均数据。
通量观测获得时间步长为30 min的净生态系统CO2交换量(Net ecosystem exchange, NEE),再将NEE 拆分为生态系统总呼吸(Ecosystem respiration,Re)和总初级生产力:
GPP=-NEE+Re
(3)
式中,Re为白天的生态系统呼吸,利用夜间NEE数据和5 cm处土壤温度之间非线性模型拟合获得。
光能利用率为总初级生产力与植被吸收的光合有效辐射(Absorbed photosynthetic active radiation, APAR)的比值,LUE的计算方式如式(4):
(4)
式中,PAR为气象观测中获取的光合有效辐射,fPAR为光合有效辐射的吸收比率(Fraction of absorbed photosynthetically active radiation),可以利用光谱观测获得的归一化植被指数(Normalized difference vegetation index, NDVI)来估算[27- 28]。
在通量塔上搭建的自动多角度光谱观测系统(Automated Multi-angular Spectro- radiometer, AMSPEC)采用的双通道光谱仪(UniSpec-DC; PP Systems),观测波段范围300—1100 nm,光谱分辨率为3.3 nm,每2—3s进行一次光谱采集,15 min 存储一次数据。将存储文件中的数据进行观测角度插值和异常值剔除,再经过白色标准板参考辐射校正后计算光谱反射率。通过公式(5)和(6)计算得到fPAR:
(5)
(6)
式中,R850和R680分别代表光谱观测到的850nm和680nm波长下的反射率;NDVImin和NDVImax分别是植被覆盖为5%和98%时的NDVI,在我国常绿针叶林分别取值0.023和0.647[29],fPARmin和fPARmax分别表示最小和最大的fPAR,分别假定为0.001和0.95[30- 31]。选择开展荧光观测日期的光谱数据,计算出NDVI后利用算数平均法计算得到与通量数据匹配的30 min时间步长的NDVI,进而获得相应的LUE日变化。
1.3 分析方法
本研究主要通过增强回归树和相关性分析来进行变量的关联及影响的分析。增强回归树(Boosted regression tree, BRT)是基于分类回归树算法(Classification and regression tree, CART)的一种自学习方法,通过随机选择和自学习方法产生多重回归树,提高模型的稳定性和预测精度[32]。BRT综合了两种算法:1)回归树算法,通过递归将数据集分成多组易建模的数据,线性回归建模;2)Boosting方法,构造预测函数序列,以一定的方式将它们组合成一个预测函数,以提高弱分类算法的准确度。在运算过程中多次随机抽取一定量的数据,分析自变量对因变量的影响程度,剩余数据用来对拟合结果进行检验,最后对生成的多重回归取均值并输出。BRT方法可以计算在其他自变量取均值或不变的情况下,某自变量与因变量的相互关系,从而得出自变量对因变量的影响载荷。本研究中利用RStudio调用BRT程序包,以环境因子为自变量,叶绿素荧光参数为因变量进行BRT分析。设置交叉验证次数为5次,学习速率为0.005,每次选取50%的数据进行分析,剩余数据进行训练。
相关性分析主要借助Pearson相关系数来反映两个变量之间线性相关程度。Pearson相关系数也称Pearson积矩相关系数,文中用R表示,其计算公式如下:
(7)
式中,n为样本量,R的取值在-1与+1之间,R的绝对值越大表明相关性越强。若R>0,表明两个变量是正相关;若R<0,表明两个变量是负相关。
2 结果与分析
2.1 湿地松和马尾松叶片荧光参数及环境因子日变化特征
ΦPSⅡ代表光适应下PSⅡ的实际光化学效率,反映叶片用于光化学反应占所吸收的光能的比例。ΦPSⅡ日变化总体上呈现“U”型,早晚高午间低(图1)。ΦPSⅡ的春季日最低值出现在午间12:00—14:00时段,夏季日最低值出现较春季有所提前,秋季日最低值出现在12:00前后,而冬季的ΦPSⅡ波动小,均维持在较高水平。NPQ反映了叶片吸收的光能以热耗散形式散失的程度,热耗散可以防御强光带来的破坏, 是植物保护PSⅡ反应中心免受伤害的重要机制。NPQ日变化总体表现为先升后降,午间达到峰值,呈抛物线型,与ΦPSⅡ日变化趋势相反。相关分析结果表明,ΦPSⅡ和NPQ有显著的线性负相关关系,在湿地松和马尾松叶片二者的相关系数R分别为-0.926(P<0.001)和-0.863(P<0.001)。
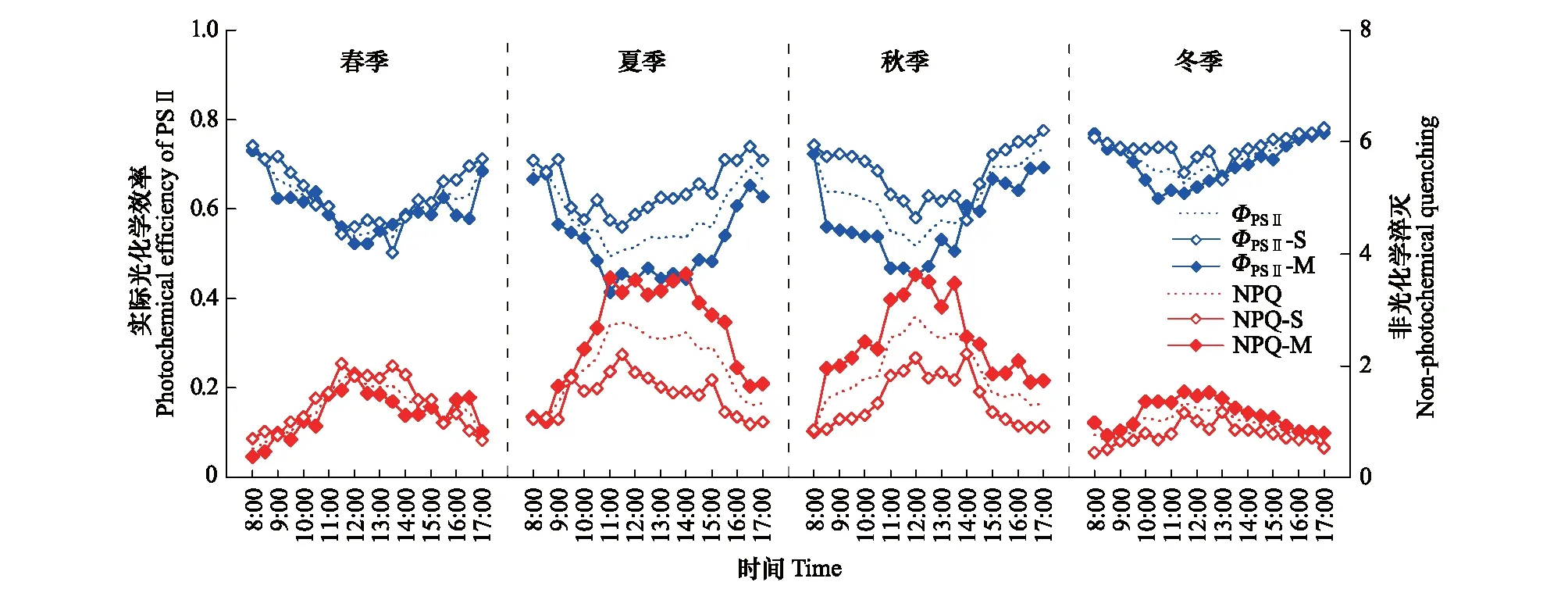
图1 湿地松和马尾松叶片ΦPSⅡ和NPQ不同季节的日变化Fig.1 Duirnal changes of ΦPSⅡ and NPQ in Pinus elliotii and Pinus massoniana leaves in different seasonsΦPSⅡ:实际光化学效率 Photochemical efficiency of PSⅡ;NPQ:非光化学淬灭 Non-photochemical quenching;S: 湿地松Pinus elliotii; M: 马尾松Pinus massoniana
同步的环境因子观测结果表明,PAR、Tleaf和RH都存在明显的日变化特征(图2)。PAR从早晨开始上升,午间达到峰值,随后下降,冬季的变化要小于其他3个季节。Tleaf从早晨开始持续上升,达到峰值后略有下降,但下降幅度很小。而RH与前二者相反,整体表现为先降后升,春、夏、秋三季早晨的相对湿度是一天中的最大值,可超过70%,14:00前后降至最低,而冬季的波动幅度很小,且基本维持在80%上下。

图2 湿地松和马尾松叶片环境因子不同季节的日变化Fig.2 Duirnal changes of environmental factors in Pinus elliotii and Pinus massoniana leaves in different seasonsPAR:光合有效辐射 Photosynthetically active radiation;Tleaf:叶片温度 Leaf tempreture;RH:相对湿度 Relative humidity
从ΦPSⅡ和NPQ以及环境因子的总体日变化特征可以看出:在光照较弱的早晨及傍晚,叶片将吸收的光能最大限度地用于光化学反应,随着光强和温度的升高,吸收的光能逐渐更多地以热耗散的形式散失;光强最强、温度最高的中午,转化为热耗散的能量比例最多,而用于光化学反应的能量比例则最少。
虽然湿地松和马尾松叶片的荧光参数和环境因子整体趋势一致,但仍存在一定差异。在夏秋季节,马尾松叶片荧光参数ΦPSⅡ和NPQ日变化的波动幅度明显大于湿地松,马尾松叶片在午间ΦPSⅡ更低而NPQ更高,其午间的日最大PAR值也要高于湿地松(表1)。湿地松叶片所受光照强度仅在春季高于马尾松,与之相应,春季NPQ湿地松叶片略高,而在夏、秋和冬季均低于马尾松叶片(图1,表1)。此外,在夏季,湿地松和马尾松叶片所受光照都是11:00前后达到最高值而后持续降低,然而,湿地松叶片的ΦPSⅡ在11:00前后达到最低值后几乎持续上升,马尾松叶片的ΦPSⅡ最低值维持至14:00前后才恢复上升,马尾松叶片的NPQ在11:00至14:00间也几乎为持续的高值。而夏季马尾松叶片日均温高达33℃,也是唯一Tleaf高于湿地松的季节(表1)。
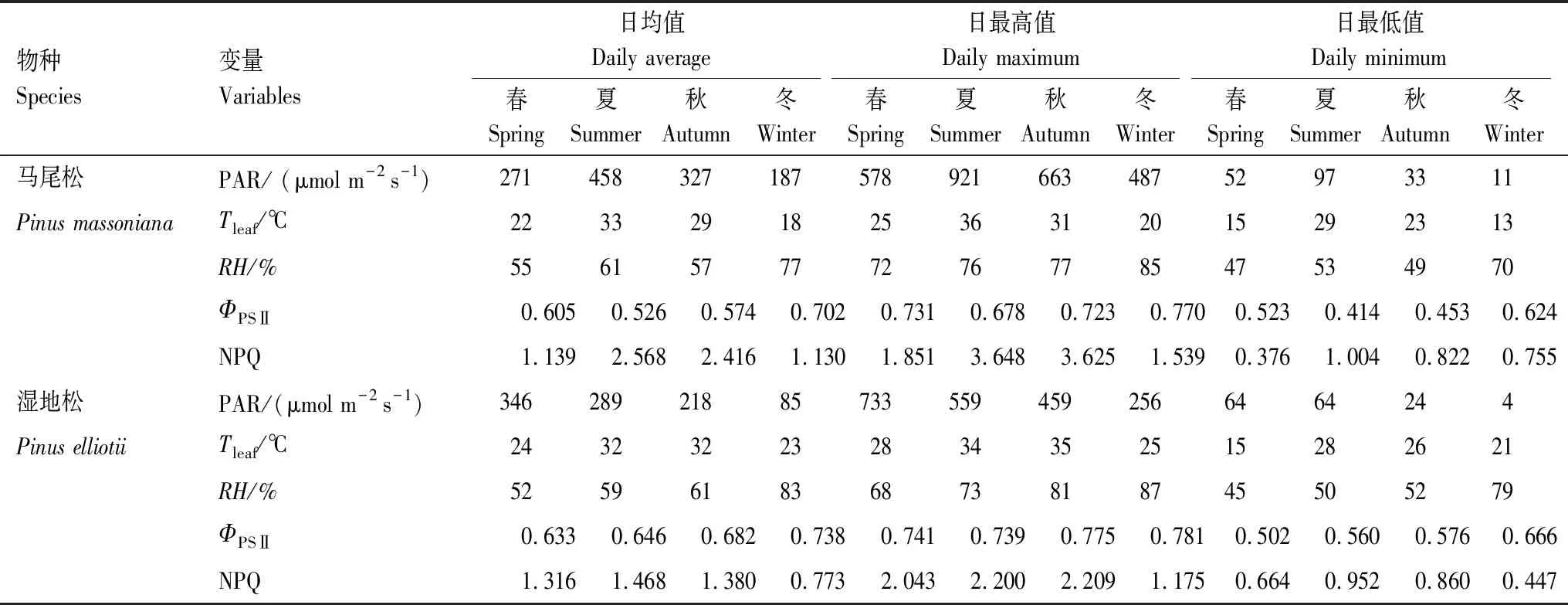
表1 不同季节湿地松和马尾松叶片环境因子和荧光参数情况
PAR:光合有效辐射 Photosynthetically active radiation;Tleaf:叶片温度 Leaf tempreture;RH:相对湿度 Relative humidity;ΦPSⅡ:实际光化学效率 Photochemical efficiency of PSⅡ;NPQ:非光化学淬灭 Non-photochemical quenching
2.2 环境因子对荧光参数ΦPSⅡ和NPQ的影响
通过增强回归树BRT分析了环境因子PAR、Tleaf和RH对湿地松和马尾松叶片叶绿素荧光参数ΦPSⅡ和NPQ的影响。4个BRT模型在R中均经过超过2500次运算达到最优,训练数据的相关性均高于0.97,交叉验证的相关性均高于0.83,可以认为模拟效果较好。BRT的分析结果如图3的柱状图所示。实际光化学效率ΦPSⅡ主要受到PAR的影响,PAR对湿地松和马尾松叶片ΦPSⅡ日变化的贡献率分别高达83.0%和51.1%。Tleaf对马尾松叶片ΦPSⅡ的影响次之(27.9%),但对湿地松叶片ΦPSⅡ的影响极低。对于非光化学淬灭NPQ而言,3个环境因子中,PAR主导了湿地松叶片NPQ的变化(77.2%),温度次之,但马尾松叶片NPQ主要受温度Tleaf的影响(59.6%),而PAR的贡献率居第二(24.8%)。

图3 环境因子对荧光参数ΦPSⅡ和NPQ的相对贡献Fig.3 The relative contribution of environmental factors on fluorescence parameters ΦPSⅡ and NPQ
图4和图5分别表示了湿地松和马尾松叶片的荧光参数与环境因子的相关分析结果,有相似也有差异。湿地松叶片的ΦPSⅡ与环境因子PAR和Tleaf都存在极显著的负相关关系(P<0.001),RH与湿地松叶片ΦPSⅡ则表现为显著正相关(P<0.001)。3个环境因子中,湿地松叶片ΦPSⅡ与PAR的相关系数最高,达-0.934,因此PAR先升后降的日变化特征主导了ΦPSⅡ的日变化的“U型”特征。湿地松叶片的NPQ与PAR的相关性同样十分显著,二者相关系数高达0.868(P<0.001),表现为良好的线性正相关。3个环境因子之间也存在一定的相关关系,湿地松叶片的RH与PAR、Tleaf均存在极显著的负相关关系(P<0.001)。
马尾松叶片的ΦPSⅡ与PAR和Tleaf同样表现为极显著的负相关关系,PAR依然是与马尾松叶片ΦPSⅡ关系最为密切的环境因子(R=-0.81,P<0.001)。但是,与湿地松叶片相比,马尾松叶片ΦPSⅡ与Tleaf的相关性更高(R=-0.764)。而且,马尾松叶片NPQ与Tleaf的相关性在3个环境因子中最高(R=0.828,P<0.001),与BRT分析的结果一致。可见,马尾松叶片的ΦPSⅡ和NPQ受Tleaf的影响相较于湿地松叶片更大。在夏季午后PAR开始下降后,Tleaf还保持着很高的水平,持续一段时间后才会下降,这二者的共同作用导致了夏季午间马尾松叶片ΦPSⅡ的持续低值和NPQ的持续高值。此外,在马尾松叶片,RH与PAR不再有显著相关关系,但与Tleaf仍然保持显著负相关。

图4 湿地松叶片荧光参数ΦPSⅡ和NPQ与环境因子的相关性Fig.4 Correlation between fluorescence parameters (ΦPSⅡ and NPQ) and environmental factors in Pinus elliotii leaves显著性水平:***,P<0.001;**,P<0.01;*,P<0.05

图5 马尾松叶片荧光参数ΦPSⅡ和NPQ与环境因子的相关性Fig.5 Correlation between fluorescence parameters (ΦPSⅡ and NPQ) and environmental factors in Pinus massoniana leaves显著性水平:***,P<0.001;**,P<0.01;*,P<0.05
2.3 荧光参数ΦPSⅡ和NPQ与植被生产力的关系
不同于叶绿素荧光参数对光合作用的表达原理,基于涡度相关数据计算的GPP和LUE从碳通量的角度表现光合作用在不同季节的日变化特征。与荧光观测同期的千烟洲亚热带人工针叶林的GPP和LUE日变化特征如图6。整体而言,GPP从早晨开始上升,午间达到峰值,随后逐渐降低。植被GPP在夏季最高,且GPP的最大值相比于其他3个季节更早出现,中午存在明显的“午休”现象。LUE的最高值多出现在早晨,随后逐渐降低,下午有所回升。但春、夏、秋三季植被LUE在下午的上升均不明显。虽然GPP在不同季节表现出了比较明显的大小差异,但是LUE在春夏秋三季相差无几,反而在GPP相对较低的冬季更高。

图6 不同季节千烟洲亚热带人工针叶林GPP和LUE的日变化Fig.6 Duirnal changes of GPP and LUE in subtropical coniferous plantation of QYZ in different seasonsGPP:总初级生产力 Gross primary productivity;LUE:光能利用率 Light use efficiency
光能利用率LUE是表征植物固定太阳能效率的指标,代表植物吸收的光能通过光合作用转化为有机物的效率,而ΦPSⅡ代表植物叶片叶绿素吸收的光能用于光化学反应(光合作用)的比例。从相关分析的结果(图7)来看,二者存在极显著的线性正相关关系(R=0.572,P<0.001),植物将吸收的光能用于光化学反应的比例高,进而转化为有机物的效率高。而NPQ作为热耗散的指标,与LUE有很好的线性负相关关系(R=-0.559,P<0.001),植物吸收的光能如果更多地用于光保护机制的热耗散,转化为有机物的效率就会相应的降低。我们也发现,GPP随着ΦPSⅡ的增加而降低(图8),二者显著负相关(R=-0.781,P<0.001)。而NPQ与GPP显著正相关(图8),R=0.788(P<0.001)。

图7 人工针叶林LUE、GPP与实际光化学效率ΦPSⅡ的散点图Fig.7 Scatter plot showing the correlation between LUE, GPP and ΦPSⅡ in artificial coniferous forest

图8 人工针叶林LUE、GPP与非光化学淬灭NPQ的散点图Fig.8 Scatter plot showing the correlation between LUE, GPP and NPQ in artificial coniferous plantation
3 讨论
3.1 荧光参数日变化的生理机制
植物的叶绿素荧光与光合作用中各个反应过程紧密相关,是植物对环境变化的生理响应,包含了丰富和复杂的信号。在本研究中,随着日间光照强度的变化,湿地松和马尾松叶片ΦPSⅡ在午间光强最强时达到最低,而NPQ在此时最高(图1)。表明在强光照下,植物光合机构所吸收的光能超过光合作用所利用的量而导致了吸收的光能用于光化学反应的比例降低,PSⅡ反应中心的光合活性下降,因此过剩的光能依赖于叶黄素循环中紫黄质向玉米黄质转变以热能的形式耗散,形成光保护机制,避免过量光照对植物光合机构的伤害[33- 34]。当午后光照强度逐渐减弱,ΦPSⅡ和NPQ又恢复到原来的水平,可见湿地松和马尾松叶片的光合活性只是受到暂时的抑制,但没有发生不可逆转的损伤,热耗散的光保护机制有效地避免了反应中心遭受过量光强而破坏。此外,本研究通过比较常绿针叶林不同季节的观测结果发现,不同时期ΦPSⅡ和NPQ日变化过程基本一致,但冬季的日变化幅度明显小于其他3个季节。同时,四个季节中冬季ΦPSⅡ最高,NPQ最低(表1),表明在光照相对较弱的冬季,叶片降低热耗散提高光化学效率以维持低光照下的光合作用水平。
3.2 环境因子的影响在不同树种的比较
通过分析环境因子对湿地松和马尾松叶片叶绿素荧光参数ΦPSⅡ和NPQ的影响发现,在观测期内,PAR、Tleaf和RH均对两种松树叶片叶绿素荧光参数有影响(图4,5),当PAR和Tleaf上升,RH下降时,湿地松和马尾松叶片通过增加热耗散降低光化学效率的保护机制来应对强光高温干旱的环境。但整体而言,PAR是影响最大的环境因子。在日变化中,Tleaf和RH的变化都相对滞后于PAR以及叶绿素荧光参数的变化,也可以说明叶绿素荧光参数更容易受到了PAR的影响。光强和温度对荧光的强烈影响同样在温带常绿针叶林被发现[35],研究表明NPQ随着光强的增长而增长,却随着温度的增长而降低,但其温度的调控作用主要在低于7.5℃时体现,而本研究发现在温度较高的亚热带常绿针叶林,NPQ随着叶片温度的增长而增长。也有研究认为,PAR是水分条件适宜时影响ΦPSⅡ降低的首要因子;而当外界可利用水分降低时,RH加剧了ΦPSⅡ的下降[16]。但本研究针对于亚热带季风区植被,观测期内的最低相对湿度仍高于45%(表1),区别于干旱区植被[16],不存在水分匮乏的情况,因此3个环境因子中,RH对荧光参数的影响最小。
然而,即使生境相似,湿地松和马尾松叶片的荧光参数也存在差异。在春季,湿地松叶片的PAR略高于马尾松叶片,湿地松叶片的热耗散略高,但其ΦPSⅡ也高于马尾松(图1,2)。表明即使在更高的光强下,湿地松叶片仍能够将吸收的光能更多的用于光化学反应,这可能与湿地松和马尾松叶片的叶面积和厚度等功能性状有关。湿地松的比叶面积(Specific leaf area,SLA)为4.92 m2/kg,低于马尾松(6.68 m2/kg)[36]。湿地松叶片将更多的干物质来构建保卫结构,增加叶片厚度和叶肉细胞密度,能够容纳更多的叶绿体,从而提高单位面积叶片的光化学效率,更有助于对外界环境变化的适应。也有研究发现相同的立地条件的湿地松叶片光合速率明显高于马尾松[37-38]。从环境因子对湿地松和马尾松叶片荧光参数影响的差异也可以看出,Tleaf对于马尾松叶片荧光参数的影响显著大于湿地松,超过PAR成为马尾松叶片热耗散的主要环境影响因子,而湿地松叶片荧光参数的变化却仅仅受到Tleaf较低程度的影响(图3)。这一结果也侧面说明了引进种湿地松相较于本地种马尾松可能更适应当地的环境。
3.3 叶绿素荧光与植被生产力的分析及展望
植被光合作用的过程中,叶绿素分子吸收的太阳能大部分用于光化学反应以及后来的电子传递,电子最终传递给NADP+(Nicotinamide adenine dinucleotide phosphate,烟酰胺腺嘌呤二核苷酸磷酸)产生NADPH(Reduced nicotinamide adenine dinucleotide phosphate,还原型烟酰胺腺嘌呤二核苷酸磷酸),为CO2的固定提供能量。叶绿素荧光不仅能反映植物光合作用过程中的光能吸收、激发能传递和光化学反应等的光合作用的原初反应过程,同样与电子传递、CO2固定等过程有关。光能利用率LUE被定义为植被将吸收的光能转化成有机干物质的效率,是陆地生态系统生产力模型估算的重要参数。我们的研究尝试将代表植物叶片将吸收的光能用于光化学反应的叶绿素荧光参数ΦPSⅡ与LUE关联,结果表明,ΦPSⅡ与LUE存在显著线性关系(P<0.001),植物叶片吸收的光能越多地用于光化学反应,也相应有越多的吸收光能被转化为有机物存储。但是,仍然存在ΦPSⅡ高而LUE低的情况。可见虽然植被叶片将吸收的光能很大程度用于进行光化学反应,CO2的固定阶段的能量转化效率可能并不高。光照是植物光合作用的主要驱动力,对于未受到胁迫的健康植物,高光照往往更有利于光合作用有机物的合成,从而对应高的GPP,而在高光照植物叶片吸收的光能用于光化学反应的比例会降低而热耗散的比例升高,因而GPP与ΦPSⅡ和NPQ分别表现为显著负相关和显著正相关。
此外,虽然我们将两种优势树种的多束叶片观测到的叶绿素荧光参数综合考虑,但基于通量数据获取的LUE和GPP更多地是反映生态系统或者植被群落的光能利用效率和光合速率情况,植被冠层结构同样可能对它们的关系有所影响[39-41]。然而,叶绿素荧光技术仍局限于叶片“点”是接触测量[42],要开展生态系统水平的观测存在一定难度,但目前利用荧光光谱探测的陆地植被的荧光遥感发展迅速,提升荧光测量尺度,为宏观、动态地了解和评价植物光合作用提供了契机[43-44]。
4 结论
通过对亚热带人工针叶林叶绿素荧光参数不同季节日变化特征及其环境因子的分析,以及对荧光参数与植被生产力关系的探讨,我们获得了以下主要结论:
(1)随着日间光照强度的变化,湿地松和马尾松叶片ΦPSⅡ(实际光化学效率)的日变化呈“U型”,在午间光强最强时ΦPSⅡ最低,而NPQ(非光化学淬灭)在此时达到峰值,与ΦPSⅡ的日变化趋势相反,表明强光照下,过剩的光能通过热耗散的形式散失以避免光合机构受到损伤。
(2)ΦPSⅡ主要受到PAR(光合有效辐射)的影响,ΦPSⅡ随着PAR的升高而降低,PAR对湿地松和马尾松叶片ΦPSⅡ日变化的相对贡献率分别高达83.0%和51.1%。NPQ随着PAR和Tleaf(叶片温度)的升高而升高,PAR主导了湿地松叶片NPQ的变化(77.2%),温度次之,但马尾松叶片NPQ主要受Tleaf的影响(59.6%),而PAR的贡献率居第二(24.8%)。
(3)GPP(总初级生产力)与ΦPSⅡ显著负相关,而与NPQ显著正相关,主要是由于光照的影响。ΦPSⅡ与LUE(光能利用率)表现为显著的线性正相关关系,但仍存在ΦPSⅡ高而LUE低的情况。NPQ与LUE显著负相关,热耗散的比例越高,转化为有机物的效率就会相应的降低。
猜你喜欢
农业与技术(2021年6期)2021-12-12
现代盐化工(2021年3期)2021-08-27
农家科技中旬版(2020年7期)2020-08-10
林业与生态(2020年2期)2020-04-16
食品与生物技术学报(2020年4期)2020-01-06
科学咨询(2020年36期)2020-01-06
中国新技术新产品(2019年21期)2019-01-20
电子技术与软件工程(2017年23期)2018-01-17
科学家(2016年12期)2017-09-29
中国市场(2017年13期)2017-06-03
