
山杏苗木对干旱胁迫的生理响应
2019-09-04 03:15:00谢东锋王华田张光灿
中国水土保持科学 2019年4期
谢东锋, 王华田, 张光灿†, 张 才
(1.山东农业大学林学院,271018,山东泰安;2.临沂大学农林科学学院,276000,山东临沂;3.山东省乳山市干果站,264500,山东威海)
干旱胁迫严重影响植物生存,干旱缺水对植物造成的伤害超过了其他逆境因子的总和[1]。近几十年来随着全球温室效应和水资源不足等环境问题日益严重,旱灾的频率、持续时间及严重程度不断增加[2],对干旱胁迫下植物生理的研究成为科研人员的研究重点[3-7]。目前我国存在着许多立地条件和水分环境极为恶劣的荒山和矿区,而植被恢复是根本性的水土保持措施,是改善这些恶劣生态环境的重要手段[8]。山杏(Prunussibirica)是我国优良的水土保持植物,根系发达,喜光耐寒,耐干旱瘠薄[9],研究山杏在干旱胁迫下的生理响应,对深入了解山杏的抗旱机制,制订科学的栽培措施,以及利用山杏进行荒山绿化、矿区植被恢复和水土保持具有重要的理论与实践价值。现有报道对山杏的光合特性[3-4]、渗透调节物质[5-6]、栽培技术[10]等方面进行研究;但很多研究是基于少数几个土壤干旱程度(如轻度、中度和重度)下的试验结果[3-4],难以明确山杏生理生化指标与土壤水分的定量关系,对于山杏土壤-植物-大气连续体(soil-plant-atomsphere continuum)水势随干旱胁迫动态变化规律的研究,还未见报道。土壤水势是土壤质地和土壤水分共同作用的函数,在描述土壤干旱程度及土壤水分对植物有效性方面,土壤水势比土壤水分更具代表性[11]。笔者采用盆栽试验和自然耗水的方法,测定土壤水势、山杏叶水势、蒸腾和光合参数的日变化特征,探讨山杏在SPAC水势动态变化下的生理响应,为山杏在生态恢复和水土保持中的应用提供参考和理论依据。
1 材料与方法
1.1 试验地概况
试验地位于山东农业大学林学试验站,地理位置为E 117°08′,N 36°11′,海拔150 m,属暖温带季风大陆性气候,年均气温为12.8 ℃,极端最高气温为40 ℃,极端最低气温为-22 ℃,无霜期186.6 d,年均相对湿度65%,年降水量600~800 mm,降水多集中于7—8月份,降水量占全年降水量的53%,而6—9月份占74%,因此春秋季严重干旱。
1.2 试验材料和测定方法
2013年7月上旬于泰安市虎山公园内1个健壮山杏母树上采集约90枚成熟种子(半同胞家系),经低温沙藏处理后于2014年4月上旬播种育苗,株行距约为15 cm×40 cm。2015年4月上旬,选择36株生长健壮、无病虫害、株高和地径相同(平均株高40 cm,地径0.9 cm)的山杏1年生苗木作试验材料,将主根统一修剪为15 cm后进行盆栽,所用花盆高20 cm、口径20 cm,每盆1株,装土10 kg,为防止根系通过排水孔插入地面,花盆下放入塑料托盘。试验用土为鲁东山丘区(淄博)露天采石场废弃地自然堆积土壤,密度为1.29 g/cm3(环刀法),田间持水量为29.8%(烘干法),pH值8.1,在4、5月份缓苗期进行常规苗木管理。
采用随机区组试验设计,分为3区组(重复),每区组12株苗木,其中6株为干旱处理试验组,6株为对照组,试验小区为长3 m、宽2 m的长方形,以避免边际效用。于6月3日20:00将苗木充分浇水至饱和(每盆浇水约0.5 L,直至水分从花盆泄水孔流出),然后自然耗水,试验组不再补充水分,对照组每天20:00浇水至饱和,采用露天试验,不进行遮阴处理(在测定期没有降雨,仅6月9日出现阴天)。胁迫处理1 d后,从6月5日开始测量数据,每天08:00—18:00期间每隔2 h使用露点水势仪(HR-33T型,美国WESCOR公司)测定各组土壤水势(Ψs)和叶水势(Ψl),连续测定7 d。Ψs的测定是每次每盆重复3次,取平均值;Ψl的测定是每次从每区组6株苗木中选2株健壮苗木,选取主干顶端向下第4枚成熟叶片进行测定,每次重复3次。在测定山杏Ψl的同时,使用光合作用测定系统(CIRAS-2型,美国PPS公司)测定所选山杏(与测定Ψl的苗木相同)气体交换参数,选择主干顶端往下第5枚健壮叶片标记,然后夹入叶室进行活体测定,重复测定3次。由于在6月8日时试验组山杏开始暂时萎蔫,除06:00外,其他时间的净光合速率(Pn)都为负值,所以光合作用气体交换参数日变化连续测定了4 d。测定参数包括Pn、蒸腾速率(Tr)、胞间CO2的摩尔分数(Ci)、大气温度(ta)、大气相对湿度(RH)、光合有效辐射强度(PAR)等。
大气水势的计算[12]:
Ψa=4.628×105Tkln(RH/100)。
式中:Ψa为大气水势,MPa;Tk为大气热力学温度,K;RH为大气相对湿度,%。
气孔限制值计算[13]:
Ls=1-Ci/Ca。
式中:LS为气孔限制值;Ci为胞间CO2的摩尔分数,μmol/mol;Ca为大气CO2的摩尔分数,μmol/mol。
1.3 数据处理
用Excel 2007和SPSS 19.0软件进行分析和作图,用单因素方差分析(One-way ANOVA)和最小显著差异法(least significant difference, LSD)在5%水平上进行差异显著性检验,图中数据为平均值±标准误。
2 结果与分析
2.1 干旱胁迫下山杏土壤-植物-大气连续体(SPAC)水势连日变化特征
在正常水分条件下,树木净光合速率最大值一般出现在09:00—11:00,笔者选择连续7个观测日10:00时对山杏SPAC水势值进行观测(图1),发现Ψa在-244 MPa上下波动,前4 d变化不显著,第5天是阴天,Ψa显著升高,5~7 d变化不明显,但显著高于前4 d(P<0.05);对照组Ψs、Ψl变化幅度很小;试验组Ψs随干旱持续呈下降趋势,第1~4天Ψs下降幅度小,第4天降至-1.33 MPa,接近大部分植物永久萎蔫点-1.5 MPa[14],此时山杏出现暂时萎蔫现象,第4~6天,Ψs则近乎以线性下降,第6天和第7天Ψs没有明显差异,但显著低于前5 d(P<0.05),第6天Ψs(-4.93 MPa)是7 d内的最小值,比最大值(第1天)降低了1 750%,而Ψl呈波动性下降趋势,第7天Ψl显著高于该日Ψs及第5、6天Ψl(P<0.05),Ψl最小值(第6天)比最大值(第1天)降低了94%,可见Ψl降低幅度显著小于Ψs。
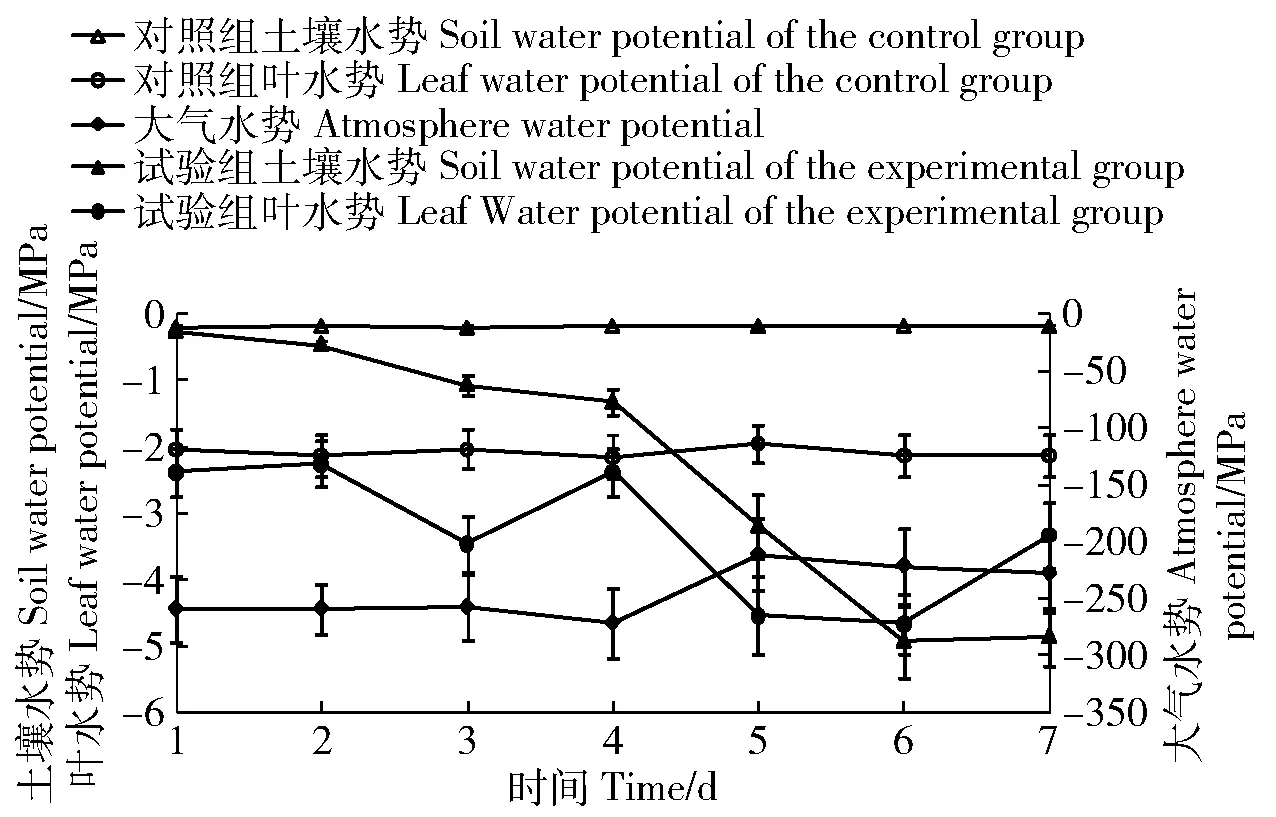
图1 连续7个观测日10:00时山杏土壤-植物-大气连续体水势变化Fig.1 Changes of water potential of the Soil—Plant—Atmosphere Continuum (SPAC) of Prunus. sibirica at 10:00 of consecutive 7 days
2.2 干旱胁迫下山杏蒸腾作用和光合作用的日变化特征

图2 山杏连续4个观测日对照组(图(a)、(b)、(c))和试验组(图(d)、(e)、(f))蒸腾速率、净光合速率和胞间CO2浓度的日变化Fig.2 Diurnal changes of transpiration rate, net photosynthetic rate and intercellular CO2 concentration of Prunus. sibirica of thecontrol group (Figure (a), (b) and (c)) and the experimental group (Figure (d), (e) and (f)) in consecutive 4 days
在山杏气体交换参数观测期间,白天ta最高37 ℃,最低23 ℃,平均31 ℃,RH最高20%,最低8%,平均14%,试验地白天出现极端高温低湿的大气干旱现象。连续4个观测日PAR、ta和RH日均值没有显著差异(P>0.05),表明不同观测日大气环境因子极为相似,影响试验组山杏苗木生理参数发生显著变化的外因主要是土壤水分的减少。对照组Tr、Pn连续4个观测日日变化均呈双峰型(图2(a),(b)),由于受到极端干热天气的影响,相对于正常水分条件的植物而言,Tr、Pn达到峰值的时间大大提前,在08:00就分别达到最高值3.63、4.90 μmol/(m2·s),然后随PAR、ta的升高和Ψs、RH的降低而逐渐下降,在中午表现出明显的光合午休现象,下午Tr、Pn峰值出现在14:00,低于上午的峰值;Ci日变化曲线与Pn相反,呈现“W”形,两低谷出现在08:00和14:00(图2(c))。Tr、Pn和Ci连续4个观测日均值变化很小,依次为(1.71±0.27)、(1.78±0.28)、(1.72±0.34)、(1.73±0.27);(1.59±0.22)、(1.53±0.31)、(1.60±0.35)、(1.50±0.20)和(325±19)、(319±16)、(316±15)、(310±15) μmol/(m2·s),差异不显著。
试验组山杏干旱胁迫第1、2天Tr和Pn日变化呈双峰型,在08:00达到峰值,最高值分别为2.23、4.80 μmol/(m2·s),然后随PAR、ta的升高和Ψs、RH的降低而逐渐下降,中午有光合午休现象,下午虽有回升,但都低于上午的峰值,Pn最小值是负值;干旱胁迫第3天Tr、Pn日变化也呈双峰型,上午峰值都提前到06:00,经过光合午休,下午峰值在14:00,但下午Tr峰值高于上午,增加了65%,Pn峰值低于上午,降低了67%,可见经过干旱胁迫和光合午休后山杏下午蒸腾作用的恢复远比光合作用迅速;干旱胁迫第4天Tr日变化近似单峰型,最大值推迟到16:00出现,说明随着水分亏缺加重,山杏需要更多的时间来恢复水分平衡,蒸腾作用最强烈的时间越来越靠近傍晚,而Pn日变化呈不规则下降趋势,在06:00达到峰值后下降,下午Pn虽有起伏,但都是负值(图2(d)和(e)),光合作用在早晨达到最大值,是植物在干旱条件下,充分利用光能进行光合作用,又尽量减少水分消耗的适应性结果;4个观测日Ci日变化有明显差异,第1、2天和对照组相似,呈“W”形,第3天呈不规则波动形,第4天呈单峰型(图2(f))。Tr和Pn连续4个观测日均值随干旱胁迫下降显著(P<0.05),4 d分别为(1.72±0.24)、(1.28±0.24)、(0.68±0.19)、(0.47±0.18)和(1.70±0.45)、(0.63±0.35)、(-0.20±0.45)、(-0.81±0.30) μmol/(m2·s),与第1天相比,第2~4天Tr和Pn日均值依次降低了26%、61%、72%和63%、112%、148%,可见干旱胁迫严重抑制了山杏的蒸腾和光合,但对光合的抑制更明显;Ci日均值随着干旱胁迫的持续显著上升(P<0.05),4 d分别为(361±20)、(395±26)、(408±29)、(434±35) μmol/mol,与第1天相比,第2~4天依次增加了9%、13%和20%。
2.3 干旱胁迫下山杏土壤水势(Ψs)和叶水势(Ψl)的日变化特征
对照组山杏每天Ψs和Ψl日变化相似,相同时间点Ψs和Ψl变化幅度很小(图1),笔者重点观测了试验组Ψs和Ψl日变化规律。连续4个观测日试验组苗木Ψs和Ψl都是从清晨开始降低,在12:00左右降到最低值后回升,呈现早晚高、中午低的“V”型曲线(图3),这是因为夜间土壤水分蒸发和植株蒸腾小,Ψs和Ψl随土壤深层水补充及山杏吸收水分而升高,白天土壤水分蒸发和植株蒸腾高于土壤内水分的移动及植株对水分的吸收,Ψs和Ψl随之降低。
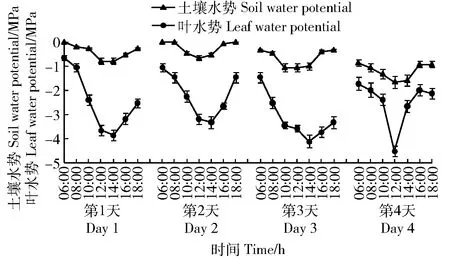
图3 连续4个观测日试验组山杏土壤水势(Ψs)和叶水势(Ψl)的日变化Fig.3 Diurnal changes of soil water potential (Ψs) and leaf water potential (Ψl) of Prunus sibirica of the experimental group in consecutive 4 days
根据气孔限制理论,植物受干旱胁迫后,Ci随Pn降低而降低,是气孔限制;Ci随Pn降低而升高,是非气孔限制[15]。在干旱胁迫第1、2天08:00—10:00,试验组山杏Ci随Pn下降而降低,Pn下降是因为气孔限制,此时Ψs范围分别为-0.20~-0.27 MPa、0~-0.47 MPa,在10:00—12:00,Ci随Pn下降而升高,Pn下降是因为非气孔限制,Ψs范围分别为-0.27~-0.80 MPa、-0.47~-0.67 MPa;在干旱胁迫第3、4天06:00—12:00,Ci随Pn下降而升高,Pn下降是因为非气孔限制,Ψs范围分别为-0.33~-1.07 MPa、-0.87~-1.67 MPa(图2(e)~(f)、图3)。可见光合作用降低的过程是从气孔限制过渡到非气孔限制,土壤越干旱,过渡到非气孔限制的时间越早,光合能力下降越快;山杏光合作用从气孔限制过渡到非气孔限制时的Ψs阈值为-0.27 MPa,低于此值时,光合作用处于非气孔限制状态,高于此值时,光合作用处于气孔限制。
水分从高水势向低水势流动,植物能否从土壤中吸取水分,取决于Ψs与Ψl间的水势梯度,在不考虑自身输导能力的情况下(如木质部导水率和栓塞脆弱性等),可将Ψs与Ψl之间的差值表示为植物对土壤水分的吸收能力,简称为水吸力(ΔΨs-l)。试验组山杏4个观测日ΔΨs-l日变化都呈单峰型,呈现早晚低,中午高的趋势(图4)。因为从凌晨开始随着土壤水分蒸发和山杏蒸腾,土壤和植株失水,Ψs、Ψl降低,Ψl下降更快(图3),因此ΔΨs-l上升,植株提高了对土壤水分的吸收;当山杏蒸腾减弱,根系吸水超过蒸腾失水时,植株开始充水,山杏叶片水分状况转好,Ψl升高,由于土壤深层水补充,Ψs也升高,但Ψl上升更快(图3),ΔΨs-l降低,这种与ΔΨs-l相关的植物充放水特性称为植物水容(C)。山杏不仅通过增强ΔΨs-l来维持对土壤水分的吸收,还能通过水容来调节自身水分平衡,增强抗旱能力。如图3所示,将试验组Ψs日变化以12:00为转折点分为06:00—12:00和12:00—18:00两个阶段,在1~3天两阶段Ψs相同时,由于水容调节,Ψl并不相同,如第2、3天10:00和14:00的Ψs大致相同,但14:00的Ψl明显低于10:00的Ψl,Ψl最小值出现的时间总迟于Ψs;但是当Ψs太低,如第4天12:00时Ψs降为-1.67 MPa,低于永久萎蔫点时,山杏很难从土壤中吸收水分,Ψl和Ψs日变化保持同步。

图4 连续4个观测日试验组山杏水吸力(ΔΨs-l)的日变化Fig.4 Diurnal changes of leaf water suction (ΔΨs-l) of Prunus sibirica of the experimental group in consecutive 4 days
3 结论和讨论
干旱是限制植物生长、分布和生存的主要环境因子之一,而矿区的立地条件和水分环境尤为恶劣,抗旱性是筛选矿区植被恢复和水土保持植物的首要条件。植物在干旱环境维持体内水分平衡的方式除了增强对环境水分的吸收(开源),就是减少自身蒸腾(节流)。Tr作为表征植物蒸腾作用强弱的参数,能够影响植物体内水分平衡,反映植物适应干旱环境的能力。笔者发现,随着干旱持续,4个观测日试验组Tr日均值显著下降,到第4日降为(0.47±0.18) μmol/(m2·s),日变化为单峰型,最高峰推迟到16:00才出现(图2(f)),表明经过4天干旱胁迫后,山杏蒸腾作用已经受到了严重抑制,山杏再通过减少自身蒸腾来节流的潜力已极低。
一般而言,植物通过2种方式吸收水分,一是通过地下部分吸收土壤水分,二是通过地上部分吸收雨、露、雾等大气凝结水。植物为了维持蒸腾拉力,Ψl要远低于Ψs,以提高ΔΨs-l吸收土壤水分;但本研究发现,干旱胁迫7天内Ψl降低幅度显著小于Ψs,到了第7天山杏Ψl显著高于Ψs,此时Ψs低于-3.1 MPa(图1),土壤水变为束缚水[14],山杏无法从土壤中吸收水分,Ψl高于Ψs显然是山杏枝叶在凌晨吸收了大气凝结水,导致叶片水分增加,Ψl上升。这也可以从解剖学上得到证明,山杏叶片蜡质和角质层在干燥时具有相对不透水性,但湿润时却更容易使水分通过[16],而旱生叶表皮细胞角化层分为内外两层,内层具有纤维素及果胶质微通道,能将水分由表面送到内部[17]。山杏通过吸收大气凝结水,缓解水分匮乏对植株的影响,是其适应干旱环境的重要机理。其实植物吸收大气凝结水是普遍存在的现象,Limm等[18]的研究表明,在夏季降雨少的美国西海岸的Polystichummunitum,其叶片吸水能力在不同区域存在明显差异[;刘志东[19]的研究表明,对凝结水利用率较高的为浅根系灌木植物和一年生草本植物;王恒方[20]的研究表明,凝结水可提高乔灌木和草本的净光合速率。
Pn是表示植物光合作用强弱的指标,能反映植物潜在生长能力的大小。本研究发现,在干旱胁迫第4日试验组山杏Pn日均值降为负值,除清晨6点为正值外,其他时间都为负值,说明在持续干旱下,山杏正常生长受到严重影响,植株光合作用明显下降。这并不代表山杏抗旱性弱,因为山杏是深根性树种[9],主要以长根系深入地下吸收水分来抗旱,但在盆栽环境下山杏吸水受到限制,因此过早显现暂时萎蔫现象。在如此干旱情况下山杏能够继续保持蒸腾,反而表明其适应干旱,维持水分平衡的能力强。在水分胁迫环境下,植物可通过合理协调碳同化速率和蒸腾耗水速率间关系,以保持生长与水分消耗的平衡,其中气孔起着重要的调节作用[21]。Schulze等[22]研究表明,杏树的气孔调节以前馈式反应为主,在大气湿度开始下降而土壤水势尚未改变前就开始调节气孔以减少水分散失。可见,山杏光合作用之所以下降迅速,是因为当环境变得干旱时,山杏会灵敏地关闭气孔,降低蒸腾,保持体内水分,以更高的水分利用效率适应干旱。张淑勇等对黄土丘陵区山杏、柠条(Caraganakorshinskii)、黄刺玫(Rosaxanthina)和山桃(Prunusdavidiana)的光合生理研究发现,山杏蒸腾速率、暗呼吸速率显著低于柠条、黄刺玫、山桃,水分利用效率明显高于柠条、黄刺玫,仅低于山桃,说明山杏对土壤水分的消耗小,干物质积累高,在黄土丘陵沟壑区有较高的发展潜力[3]。可见,灵敏地进行气孔调节,以降低蒸腾,提高水分利用效率,是山杏适应干旱的重要机制。
山杏具有耐旱型特征,当受到干旱胁迫时,山杏体内脯氨酸、可溶性糖等渗透调节物质数量急剧升高[5-6],不仅能降低Ψl,提高渗透调节能力,还能防止细胞在干旱环境下脱水,维持光合呼吸等生理作用稳定性。本文发现,山杏能够通过降低Ψl,提高ΔΨs-l来增强对土壤水分的吸收,提高耐旱能力。山杏增加ΔΨs-l可能与渗透调节物质相关,今后应加强两者的相关性研究。水容在树木耗水过程中起着极其重要的作用,能够极大的缓解蒸腾过程对树木根系吸水的压力[23]。山杏水容充放水特性不仅与植株本身的生物学特性有关,与环境水分的多少也紧密联系。本文发现,当Ψs高于永久萎蔫点时,Ψl的变化总是滞后于Ψs的变化,这是因为山杏植株水容具有放水功能,山杏还能通过提高ΔΨs-l增强水分吸收,使体内水分变化较缓,Ψl不会随Ψs降低而迅速下降;当Ψs低于永久萎蔫点时,Ψl和Ψs日变化保持同步,是因为山杏根系能吸收的土壤水分减少,ΔΨs-l升高也无法吸收更多的水分,而水容充放水具有一定的阈值,经过连日消耗,山杏体内水分大幅度降低,水容调节能力下降,最终导致Ψl与Ψs同步降到最低。根据植物水容充放水特性,可将植物体内水分日变化分为2个阶段,即植物水容以放水为主的阶段(从凌晨蒸腾失水量大于植物吸水量时开始到下午蒸腾失水量等于植物吸水量为止)和植物水容以充水为主的阶段(从下午蒸腾失水量小于植物吸水量开始到次日凌晨蒸腾失水量等于植物吸水量为止)。据此可将山杏水容日变化分为2阶段,在第1~3天,06:00—14:00属于水容放水阶段,14:00—18:00属于充水阶段;在第4天,06:00—12:00属于水容放水阶段,12:00—18:00属于充水阶段。在水容充放水不同阶段山杏生理活动存在差异,是因为山杏能通过水容调节自身生理,在不超出放水阈值时,能够保持正常的生理活动,所以说,水容调节是山杏能够防御干旱的重要生理机制。在自然条件下,植物充放水是同时进行的,只是一个阶段以充水为主,另一阶段以放水为主,在今后的工作中应对水容这一动态变化进行进一步的模型构建和量化研究。
植物抗旱性是一个涉及其形态解剖结构、水分生理生态特征、光合生理生化反应的综合过程,这些生理过程常是同时发挥作用的。从本文研究可知,山杏通过吸收大气凝结水、水容调节、提高水吸力及气孔调节来减少体内水分流失,维持水分平衡以抵抗干旱胁迫,因此可将山杏作为矿区植被恢复和水土保持的优良植物材料。但是在应用中需要注意的是,山杏是深根性树种,在土层深厚的地方生长良好,应与长柄扁桃(Amygdaluspedunculata)等浅根性植物配合使用,或作为先锋树种,在退化生态系统恢复中促进群落的正向演替。
猜你喜欢
现代农村科技(2022年11期)2022-12-16 19:35:59
现代农村科技(2021年3期)2021-04-11 10:43:28
辽宁林业科技(2017年4期)2017-06-22 10:23:36
高原山地气象研究(2016年2期)2016-11-10 06:06:27
水利科技与经济(2016年3期)2016-04-09 13:57:20
水利科技与经济(2016年4期)2016-04-09 03:49:18
水利科技与经济(2016年2期)2016-04-09 02:09:13
湖南安全与防灾(2015年10期)2015-03-03 06:02:10
塔里木大学学报(2014年3期)2014-03-11 18:47:27
自然资源遥感(2014年2期)2014-02-27 11:56:12
