
藏北高寒区越冬期间野生早熟禾根系生理特征及抗寒性比较
2019-09-03 07:50周娟娟秦爱琼黄利春汪丽军
草业科学 2019年8期
周娟娟,魏 巍,桑 旦,秦爱琼,黄利春,汪丽军
(1. 青稞和牦牛种质资源与遗传改良国家重点实验室 / 西藏自治区农牧科学院草业科学研究所,西藏 拉萨 850000;2. 内蒙古自治区科尔沁右翼前旗农业技术推广中心,内蒙古 科尔沁 137713)
藏北地区位于青藏高原腹地,被称为“世界屋脊的屋脊”,平均海拔4 500 m以上,气候寒冷干旱,自然条件极为严酷[1],却肩负着国家生态安全屏障的使命,也承担着西藏乃至全国畜牧业基地的重任。近年来,在藏北草原受生态保护和发展畜牧双重压力作用,草地生产力下降和草畜矛盾尖锐成为该区面临的主要问题[2]。因此,开展栽培草地种植是保护生态、缓解高寒草地压力、解决牲畜缺草问题的有效途径。西藏自20世纪70年代开始引种工作,引进品种不下百余种,但在高寒特殊的气候条件下,绝大多数品种不能安全越冬,很少能完成生育周期[3]。藏北地区越冬期低温是多年生牧草生产面临的瓶颈问题之一,筛选生态适应能力强的乡土草种成为解决上述问题的关键。早熟禾属(Poa)植物资源丰富,全球有500余种,广泛分布在凉冷湿润区[4],西藏记载有31种和1变种[5],不同资源生长生境存在很大差异。因此,开展早熟禾属资源开发利用对高寒区退化草地改良、缓解草畜压力具有重要的实际意义。
许多研究表明,植物生理指标是评价抗逆性强弱的主要参数。张尚雄等[6]利用生理指标确定不同地区野生披碱草属(Elymus)苗期抗寒性强弱,魏双霞等[7]发现,与暖温半干旱气候区相比,种植在高寒区的紫花苜蓿(Medicago sativa)的可溶性糖含量、可溶性蛋白含量、超氧化物歧化酶活性等均有不同程度增加。吕优伟等[8]发现,低温胁迫下草地早熟禾(Poa pratensis)苗期可溶性蛋白含量、可溶性糖含量、抗氧化酶活性与其耐寒能力正相关。那曲地区冬季漫长寒冷,低温冻害时有发生,为驯化材料抗寒性研究提供了有利的环境条件。本研究以西藏本地不同海拔的早熟禾属材料为研究对象,测定越冬前后其生理指标的变化规律,并计算各指标抗寒系数、隶属函数值来综合评价抗寒性强弱,以期为西藏高寒区多年生抗寒种质资源的筛选提供方法,为早熟禾属牧草的抗寒育种以及栽培草地建植提供技术和理论依据。
1 材料与方法
1.1 试验地概况
试验区位于那曲市那曲镇的“藏北高寒草地生态科技园”内,坐标 31°26' N,92°01' E,海拔 4 512 m。该区年平均气温为-2.2 ℃,极低温低于-25 ℃,≥ 5 ℃ 年积温为 940.6 ℃·d(1990-1999 年)[9]。年降水量为406.2 mm,年蒸发量为1 811.2 mm,年日照时数为2 789.9 h(1955-2011年)[10]。气候干燥、多风。雨热同季,降水主要集中在5-9月,无绝对无霜期,植物5月底返青,9月中旬枯黄,青草期100 d左右。研究区属高原亚寒带季风半湿润气候,土壤类型为高寒草原土,土壤有机质缺乏,仅0.24%为矿质土壤。土壤全氮、全磷和全钾含量分别为2.37、0.68和3.91 g·kg-1,土壤碱解氮、速效磷和速效钾含量分别为78.52、19.14和217.81 mg·kg-1,土壤pH 7.85。2016年入冬前(9月)至翌年返青期(5月),月均气温与土壤温度均在1月出现最低值,土壤0 - 10 cm土层温度为-6.6 ℃,近地气温为-9.8 ℃(图 1)。
1.2 供试材料
供试材料为藏北不同海拔采集的早熟禾属野生种(表1);对照(CK)是青海冷地早熟禾(栽培品种),由青海畜牧兽医科学院提供。
1.3 试验设计
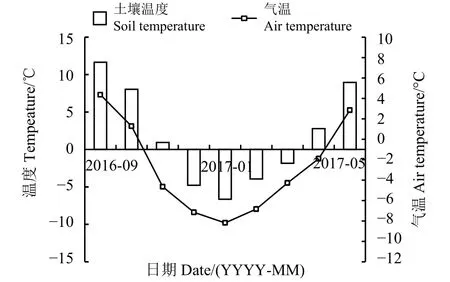
图 1 2016年9月至2017年5月试验区气温、土壤温度变化Figure 1 Variation in air temperature and soil temperature in the test area from 2016 to 2017
试验于2015年6月播种,播种时试验小区施底肥,有机肥500 g·m-2,试验小区采用随机区组设计,小区面积3 m × 5 m = 15 m2,3次重复,人工开沟条播,播深3~5 cm,行距35 cm,播量1.5 g·m-2,小区之间起垄相隔,垄宽0.5 m。进行常规田间管理,播种第1年苗期进行人工拔除杂草。
1.4 测定项目和方法
1.4.1 越冬率
每个小区选择1 m样段分别于植物枯黄前(2016年8月31日)和翌年返青后(2017年6月2日)统计植株存活株数,计算越冬率(%),越冬率 = (冬后返青苗数/入冬前植株数) × 100%。
1.4.2 生理指标
于入冬前(2016年9月10日)、最冷月(2017年1月15日)、返青期(2017年5月20日) 3个时期挖根采样,室内将根洗净,取根颈下端5 cm根段放入-80 ℃ 冰箱保存,进行生理指标的测定,重复3次。
根系中的可溶性糖含量、可溶性蛋白含量、丙二醛(MDA)含量、超氧化物歧化酶(SOD)活性和过氧化氢酶(CAT)活性分别采用蒽酮比色法、考马斯亮蓝染色法、TBA法、黄嘌呤氧化酶法和钼酸铵比色法测定[11]。
1.5 分析方法
以文献[12-13]分析方法为依据,通过4个材料越冬前后丙二醛、可溶性糖、可溶性蛋白和超氧化物歧化酶等指标的变化,进行抗寒性评价。计算各指标抗寒系数(cold-resistance coefficients,PI)、综合抗寒系数(comprehensive cold resistance coefficient,RI)、隶属函数值μ(x),计算公式如下:
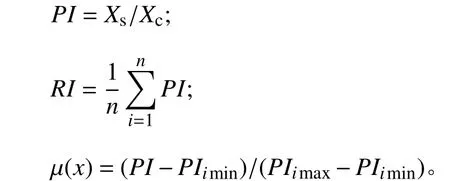
式中:Xs和Xc分别为越冬后和对照下各供试材料各指标的测定值,Xs为该指标在寒冷胁迫下的平均值,PIimin、PIimax为各性状抗寒系数的最小值和最大值。

表 1 试验材料采集地概况Table 1 Profile of seed collecting plots
1.6 数据处理
利用Excel 2007对试验数据进行整理及图表的绘制,利用SPSS 16.0统计软件中Duncan检验对不同时期、不同材料间各项生理指标进行显著性(0.05水平)方差分析。
2 结果与分析
2.1 早熟禾材料的越冬率
4个早熟禾材料越冬率均超过了80%,但不同材料之间越冬率存在明显差异,由高到低依次为中亚早熟禾(P-1) > 渐尖早熟禾(P-2) > 冷地早熟禾(P-3) > 青海冷地早熟禾(CK),分别为88.50%、85.85%、84.54%和81.46%(图2)。3个野生种的越冬率均显著高于栽培品种 (P < 0.05)。
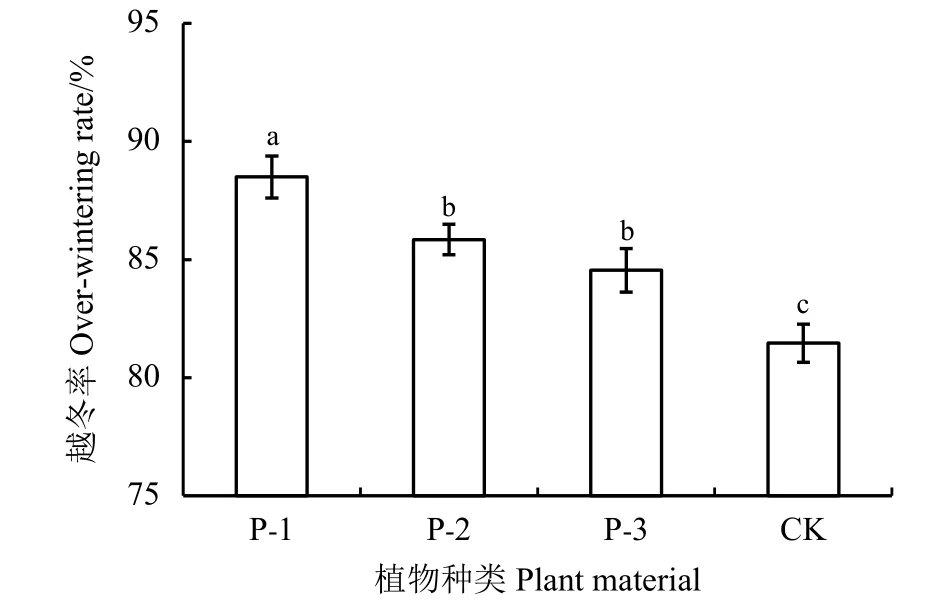
图 2 不同供试材料越冬状况Figure 2 Wintering rate of four tested materials材料代号对应的材料名称如表1所列,下同。不同小写字母表示不同材料间差异显著(P < 0.05)。The material codes are the same as those in Table 1. Difference lowercase letters indicate significant difference between different materials at the 0.05 level.
2.2 越冬期间根系生理指标变化
2.2.1 根系可溶性糖含量变化
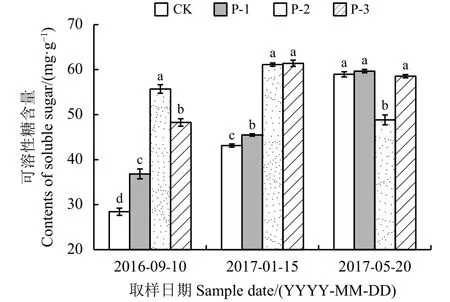
图 3 不同供试材料根系可溶性糖含量变化Figure 3 Change in soluble sugar content in roots of four tested materials柱状图上不同的小写字母表示同一时期不同材料间的差异显著性 (P < 0. 05)。下同。Different lowercase letters on the bar chart indicate significance differences among different materials at the same time at the 0.05 level;similarly for the following figures.
CK、P-1、P-2和P-3可溶性糖含量变幅分别为30.53、22.85、12.28 和 13.04 mg·g-1(图 3)。整个越冬期内,CK、P-1可溶性糖含量随时间推移逐渐上升,最大值出现在返青期(5月20日);P-2和P-3可溶性糖呈现先增加后降低的趋势,与气温变化相一致,最冷月可溶性糖含量高于入冬前与返青期。同一时期不同材料比较发现,入冬前4个材料根系可溶性糖含量存在显著差异(P < 0.05),P-2含量最高;最冷月P-2和P-3的可溶性糖含量显著高于CK和P-1 (P < 0.05),返青期P-2的含量显著低于其他 3 个材料 (P < 0.05)。
2.2.2 根系可溶性蛋白含量变化
整个越冬期内,4个材料根系可溶性蛋白含量变化一致,均呈持续增加的变化趋势,返青期与入冬前相比,CK、P-1、P-2和P-3根系可溶性蛋白含量分别提高了 0.73、0.84、0.96和 1.11 mg·g-1(图4)。最冷月P-1、CK可溶性蛋白含量显著低于P-2和P-3(P < 0.05);返青前P-1含量显著低于P-3(P < 0.05)。
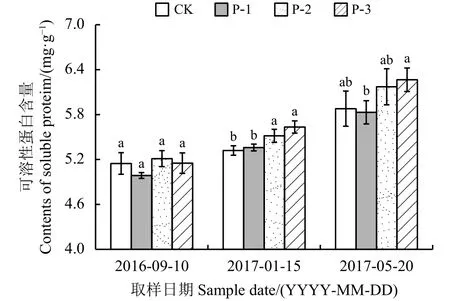
图 4 不同供试材料根系可溶性蛋白含量变化Figure 4 Change in SP content in the roots of four tested materials
2.2.3 根系丙二醛含量变化
整个越冬期内,4种材料根系中丙二醛(MDA)的变化呈现先增加后降低的趋势,与气温、表土温度变化一致,均在最冷月达到最大值(图5)。入冬前-最冷月MDA含量急剧增加,CK、P-1、P-2和P-3分别增加了21.16、13.62、15.40 和17.77 nmol·g-1;最冷月-返青期CK、P-1和P-2有小幅度降低,返青期MDA含量明显高于入冬前。同一时期不同材料比较发现,入冬前不同材料MDA含量差异不明显,最冷月和返青期,P-1的MDA含量最低。
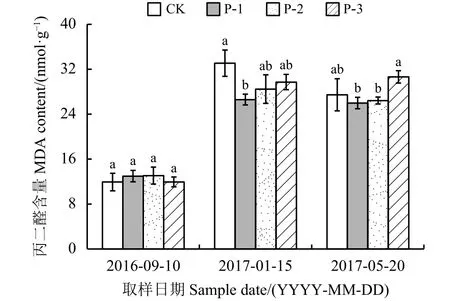
图 5 不同供试材料根系丙二醛含量变化Figure 5 Change in MDA content in the roots of the four tested materials
2.2.4 根系超氧化物歧化酶活性变化
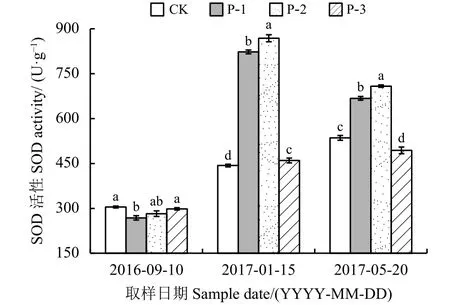
图 6 不同供试材料根系SOD活性变化Figure 6 Change in SOD activity in the roots of the four tested materials
整个越冬期内,4种材料根系中超氧化物歧化酶(SOD)活性呈现不同的变化规律,CK和P-3从入冬前到返青期呈现升高的趋势(图6);P-1和P-2呈现先升高后降低的现象,在最冷月活性最高,分别为822.68和868.21 U·g-1。入冬前-最冷月,P-1和P-2活性升高迅速,分别升高554.26和585.92 U·g-1。同一时期不同材料比较发现,4种材料根系超氧化物歧化酶活性有显著差异(P < 0.05),在最冷月、返青期P-1、P-2活性均显著高于CK和P-3 (P < 0.05)。
2.2.5 根系过氧化氢酶活性变化
4种材料根系过氧化氢酶(CAT)活性随越冬期间气温变化呈现不同的变化趋势,入冬前-最冷月,随气温降低,P-1、P-2材料CAT活性迅速升高,升高幅度分别为231.98和140.56 U·g-1(图7)。最冷月-返青期,随气温上升,P-1、P-2的CAT活性缓慢降低,降幅为49.14和39.18 U·g-1。而在整个越冬期内,CK的CAT活性持续下降,降幅为23.86 U·g-1;P-3变化趋势为持续升高,升高幅度为10.06 U·g-1。同一时期不同材料比较发现,在最冷月和返青期,4种材料根系过氧化氢酶活性差异显著(P < 0.05),均以P-1材料的活性为最高。

图 7 不同供试材料根系CAT活性变化Figure 7 Change in CAT activity in the roots of the four tested materials
2.3 供试早熟禾材料抗寒性评价
应用单项指标进行抗寒性评定不能准确评价牧草抗寒等级。本研究以入冬前4种材料各性状指标的测定值为对照值,计算单项指标的抗寒系数。通过综合抗寒系数和隶属函数法进行4种材料抗寒性等级评定。结果表明(表2),综合抗寒系数和隶属函数法计算值,均表现出P-1的评价值为最大,值分别为2.164和为0.604,抗寒性最强。其次为P-2,值分别为2.114和0.501。抗寒性评定较差的材料为青海冷地早熟禾(栽培品种)。
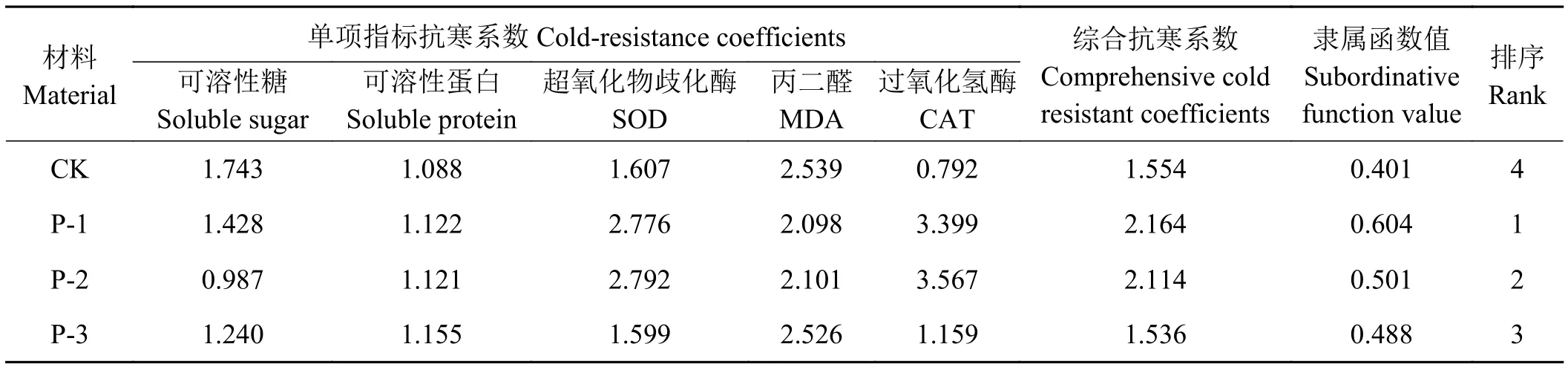
表 2 供试材料抗寒系数Table 2 Cold-resistance coefficients of tested materials
参照连续变数的次数分布统计方法[13],将4种材料各性状的抗寒系数以组距0.5分成7个组区间,制作成次数分布表(表3)。结果表明,在同一组区间各性状的抗寒系数分布次数相差较大,在4种材料中,100%的可溶性蛋白含量的抗寒系数PI [1.0,1.5),50%的SOD活性的抗寒系数PI ≥ 2.5,50%的丙二醛的抗寒系数PI < 2.5,50%的可溶性糖含量的抗寒系数PI [1.0,1.5)。说明野生材料和其他栽培作物一样,不同性状对寒冷胁迫响应的敏感程度不同,可溶性蛋白含量的反应非常敏感,SOD活性、可溶性糖和丙二醛反应较敏感。
3 讨论与结论
3.1 渗透物调节物对抗寒性的影响
研究表明,可溶性糖是植物抗寒的一种保护物质,也是植物安全越冬和翌年再生的物质和能量来源,不同植物可溶性糖的含量不尽相同。当植物受到寒冷胁迫时,其含量的增加能够提高细胞内溶质的浓度,降低冰点,提高细胞的持水力,进而提高植物抗寒能力[14]。有研究表明,紫花苜蓿主根中可溶性糖在越冬前期随越冬温度下降明显增加,在翌年返青含量又随之降低[15]。低温胁迫,早熟禾可溶性糖含量随温度下降呈现增加的趋势[8]。本研究中,4个材料在入冬前(9月)至最冷月(1月)随温度降低,根系中可溶性糖含量呈现不同程度的增加,表明低温胁迫过程,早熟禾根系通过增加可溶性糖积累,来降低植物保水能力,防止膜质氧化,进而对根系起到保护作用。在最冷月至翌年返青期(5月)随温度回升,青海冷地早熟禾(栽培品种)和中亚早熟禾可溶性糖含量继续升高。李轶冰等[16]在对美国引进的高山早熟禾和爱达荷冰草研究中也得出了相同的结论(即植物经过冷冻处理后,在温度回升的过程中植物可溶性糖含量是持续升高的)。而渐尖早熟禾和冷地早熟禾(野生种)有小幅度的下降,可能原因是此时供试材料已开始返青萌动,根系糖类物质由保护物质转化为能量物质,少量可溶性糖被消耗利用。
可溶性蛋白含量的变化与植物抗寒性的关系密切,植物受低温胁迫时,通过增加蛋白来束缚更多的自由水,降低渗透势,从而减少原生质因结冰而伤害致死的机率[17],提高植物抗寒性。南丽丽等[18]研究紫花苜蓿越冬期生理变化发现,根系中可溶性蛋白含量随气温的降低而增加,随翌年气温的回升又呈现小幅度降低。也有研究显示,可溶性蛋白与温度无相关性[19]。本研究,入冬前至返青期可溶性蛋白含量持续增加。这种相悖的研究结果可能与不同材料返青恢复时间有关,关于返青多长时间后才能消除寒冷对可溶性蛋白积累的滞后效应还需进一步研究。
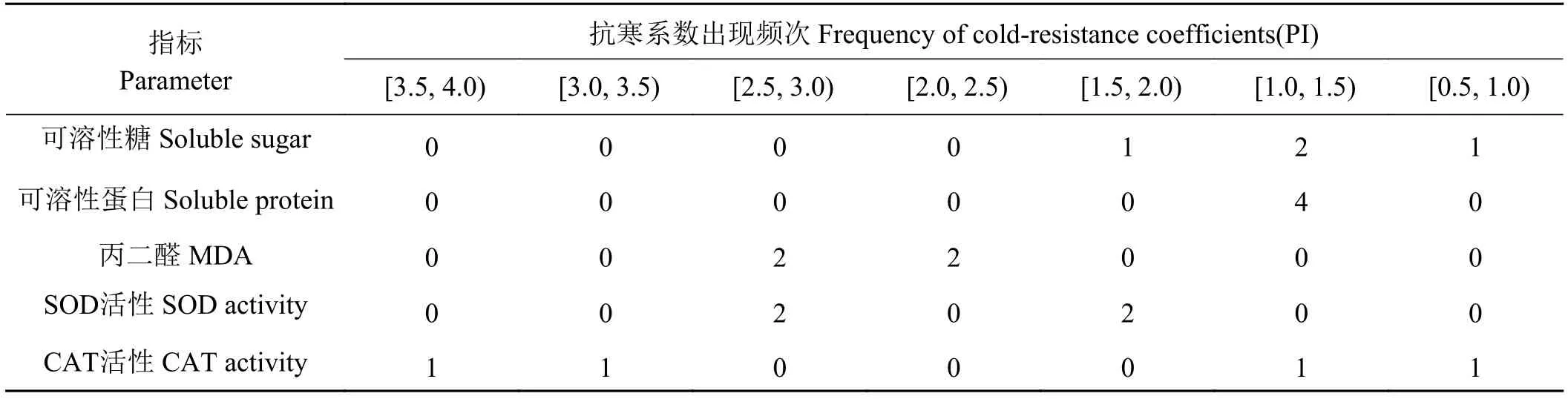
表 3 供试材料各性状指标的抗寒系数在不同区间的分布Table 3 Different distribution of cold-resistance coefficients of tested varieties
3.2 丙二醛和抗氧化酶对抗寒性的影响
低温胁迫能诱导植物产生氧自由基,高浓度的氧自由基会迫害植物细胞膜质,造成细胞死亡[20],SOD和CAT作为抗氧化酶防御系统的主要酶系,可降解细胞内有害自由基[21],对增强植物抗寒性有重要作用。本研究中,入冬前至最冷月,随气温降低,中亚早熟禾和渐尖早熟禾根系中SOD和CAT活性迅速升高,表明该时间段酶活性较高可有效清除细胞内自由基,膜质的抗氧化作用较强。最冷月至返青期酶活性降低较为缓慢,说明此阶段该区空气温度、土壤温度仍然处于较低水平,SOD和CAT还能够对早熟禾根系有很好的保护作用。反观野生冷地早熟禾和青海冷地早熟禾,在寒冷胁迫下,SOD和CAT活性提升较为缓慢,抗寒能力相对较弱。
丙二醛是植物遭受低温胁迫氧自由基积累造成细胞膜质过氧化反应的最终产物。其量的大小直接关系到细胞膜被毒害程度。因此,也被作为衡量植物抗寒性的重要参数[22-23]。本研究中,4个材料根系丙二醛含量水平偏低,表明它们对寒冷环境都有较好的适应性。随越冬期间温度的降低丙二醛含量总体呈上升的趋势,说明植物在寒冷胁迫下,不同材料细胞膜质过氧化有一定程度增加。中亚早熟禾和渐尖早熟禾升高幅度较慢,最冷月至返青期含量逐渐趋于缓和,表明这两种早熟禾在寒冷环境下膜质过氧化程度较低,具有较强的清除氧自由基的能力,Bonnecarrere等[24]在水稻(Oryza sativa)的研究中也有相同结果。
3.3 早熟禾抗寒评价
植物越冬期抗寒能力是其生理生化特性综合作用的结果,利用单一的指标很难判定植物对寒冷的适应能力。Fuzzy数学隶属函数法被广泛运用在紫花苜蓿[18]和禾本科牧草[16]越冬及混播草地建植[25]的抗寒性评判中,并得到了较为满意结果。本研究利用隶属函数法和综合抗寒系数对早熟禾植物根系中可溶性糖、可溶性蛋白、丙二醛、过氧化氢酶和超氧化物歧化酶5项指标进行综合分析,隶属函数值和综合抗寒系数均表现出相同的指示结果,抗寒性顺序为中亚早熟禾 > 渐尖早熟禾 > 冷地早熟禾(野生种) > 青海冷地早熟禾(栽培种)。
4 结论
藏北高寒区常年低温,不同野生早熟禾的抗寒性不同。越冬期参试早熟禾材料根系的SOD活性和CAT活性均随气温下降而升高,进入最冷月开始下降;而可溶性蛋白含量随气温下降而上升,翌年返青后继续随气温升高持续升高。采用综合抗寒系数、隶属函数对4个供试早熟禾的生理指标进行综合分析,抗寒性强弱依次为中亚早熟禾、渐尖早熟禾、冷地早熟禾和青海冷地早熟禾,与越冬率的表现一致。
猜你喜欢
蔬菜(2022年10期)2022-11-10
当代水产(2022年7期)2022-09-20
热带作物学报(2022年5期)2022-06-01
今日农业(2021年21期)2021-11-26
今日农业(2021年20期)2021-11-26
河北果树(2020年4期)2020-11-26
河北果树(2020年1期)2020-02-09
现代园艺(2017年11期)2017-06-28
中国农业文摘-农业工程(2016年5期)2016-04-12
中国果菜(2016年9期)2016-03-01
