
膜荚黄芪毛蕊异黄酮葡萄糖苷积累规律研究
2019-09-02 14:01吕爽包蕊刘俊频袁元全雪丽吴松权
江苏农业科学 2019年5期
关键词:病虫害
吕爽 包蕊 刘俊频 袁元 全雪丽 吴松权
摘要:为了解膜荚黄芪毛蕊异黄酮葡萄糖苷的积累规律,为膜荚黄芪的种植和采收提供理论依据,测定了自然环境条件下膜荚黄芪生长中的生物量,毛蕊异黄酮葡萄糖苷含量,土壤温度和湿度,并分析了毛蕊异黄酮葡萄糖苷积累变化规律。总体而言,随着生物量的增加,毛蕊异黄酮葡萄糖苷含量在叶中呈下降趋势,在茎中呈上升趋势,而在根中表现出先下降再上升的趋势、且与土壤温度和水分呈显著负相关,病虫害的发生则明显降低了生物量和毛蕊异黄酮苷的积累。研究得出的结论是膜荚黄芪毛蕊异黄酮葡萄糖苷的积累具有组织特异性,其在地上部和地下部的含量受不同机制的调控,适当的低温和水分胁迫有利于根中毛蕊异黄酮糖苷的积累。
关键词:膜荚黄芪;毛蕊异黄酮葡萄糖苷;积累规律;病虫害
中图分类号:S567.23+9.01 文献标志码: A 文章编号:1002-1302(2019)05-0103-03
收稿日期:2018-06-07
基金项目:国家自然科学基金(编号:21462044、30860036)。
作者简介:吕 爽(1993—),女,吉林松原人,硕士研究生,研究方向为特种植物资源。E-mail:893226771@qq.com。
通信作者:全雪丽,博士,教授,硕士生导师,研究方向为特种植物资源,E-mail:qxl2000@aliyun.com;吴松权,博士,副教授,硕士生导师,研究方向为特种植物资源,E-mail:arswsq@ybu.edu.cn。
黄芪为豆科(Leguminosae)植物蒙古黄芪[Astragalus membranaceus (Fisch.)Bge. var. mongholicus(Bge.)Hsiao]或膜荚黄芪[Astragalus membranaceus (Fisch.)Bge.]的干燥根,具有补气升阳、固表止汗、利水消肿、生津养血、行滞通痹、脱毒排脓的功效[1]。黄芪含有多种化学成分,主要有皂苷类、多糖类和异黄酮类等[2-3]。其中毛蕊异黄酮葡萄糖苷是黄芪中主要的异黄酮类活性成分,具有抗炎、抗病毒、抗菌、清除超氧阴离子、保护肝细胞、增加缺血再灌注大鼠脑细胞中膜的流动性[4]、预防骨质疏松、增强骨质再生等作用[5],并且对糖尿病肾病的发展也具有潜在的调控作用[6]。2015年版《中华人民共和国药典》中规定毛蕊异黄酮葡萄糖苷是评价黄芪药材质量的“标记化合物”之一[1]。目前,普遍采用高效液相色谱法(high performance liquid chromatography,简称HPLC)来检测黄芪中毛蕊异黄酮葡萄糖苷含量,此法具有准确可靠、操作简便、重复性好的优点[1,7-8]。
黄芪是名贵中药材,由于长期大量采挖导致野生资源急剧减少,野生黄芪已被列为国家3级保护植物[9]。膜荚黄芪主产于黑龙江、吉林、内蒙古、河北、山西等地[3]。《药物出产辨》(1920年)记载:“正芪产区有3处:1关东,2宁古塔,3卜奎”,其中宁古塔为黑龙江省宁安和吉林省延吉一带(延边朝鲜族自治州,即延边地区)。但在延边地区这一优质的道地药材由于野生资源被过度利用而不能满足人们的需求,且人工栽培黃芪还不能代替传统的野生黄芪[10]。延边地区位于吉林省长白山麓,属中温带湿润季风气候,春季干燥多风,夏季温热多雨,秋季凉爽少雨,冬季寒冷漫长。由于其独特的地理环境和气候条件,形成了复杂的植被类型和丰富的野生植物资源,特别适合发展中草药产业[11-12]。本研究的目的是以膜荚黄芪为植物材料,研究自然生长条件下根、茎和叶中毛蕊异黄酮葡萄糖苷的积累规律,为延边地区膜荚黄芪栽培和利用提供理论依据。
1 材料与方法
1.1 植物材料
黄芪种子和植株采自吉林省延吉市,经延边大学农学院植物学教研室石铁源教授鉴定为膜荚黄芪。凭证标本留存于延边大学农学院植物学教研室。
1.2 试剂和主要仪器
对照品毛蕊异黄酮葡萄糖苷(批号:110907)购自上海融禾医药科技有限公司,质量分数大于98%;色谱级乙腈购于美国Fisher公司;色谱级甲醇购于美国迈瑞达科技有限公司;水为超纯水;其他试剂均为分析纯。
岛津高效液相色谱仪(LC-10ATVP Plus),包括二元高压梯度泵(LC-10ADVP)、紫外可见双波长检测器(SPD-10AVP Plus)、自动进样器(SIL-10A)、柱温箱(CTO-10ASVP Plus);C18反向色谱柱(华谱科仪科技有限公司);RE-2000B型旋转蒸发仪(上海亚荣生化仪器厂);DLSB-5/20型低温冷却液循环泵(上海豫康科教仪器设备有限公司);RE-501型恒温水浴锅(上海豫康科教仪器设备有限公司);KQ-500D型数控超声波清洗器(昆山市超速仪器有限公司);精密鼓风干燥箱(上海施都凯仪器设备有限公司);RHP-600型高速多功能粉碎机(浙江荣浩工贸有限公司);土壤温湿度测定仪(顺科达)。
1.3 方法
1.3.1 黄芪种植和采收 于延边大学农学院实验基地(吉林省延吉市)进行。2015年4月10日播种,6月10日后现真叶,10月枯萎;越冬后,次年4月下旬开始返青,10月枯萎。由此确定1年生采收期从7月开始,2、3年生采收期从5月开始,截止至10月,具体采样日期为每月10日。每次采样时取9株植株,3株植株为1次重复。采样后,将样品于50 ℃烘干箱中烘干至恒质量,测定生物量。将干燥样品磨至粉末状,过60目筛,用于毛蕊异黄酮葡萄糖苷的提取。
1.3.2 毛蕊异黄酮葡萄糖苷含量的测定
1.3..2.1 供试品溶液 参考2015年版《中华人民共和国药典》方法[1]。精确称取黄芪样品粉末0.5 g,放入圆底烧瓶中,加入分析级甲醇50 mL,于80 ℃水浴锅中加热回流4 h,冷却至室温,用沃特曼(Whatman)中速滤纸过滤后,在50 ℃下旋转蒸发浓缩至干,用5 mL色谱级甲醇溶解,0.45 μm微孔滤膜过滤,转移到5 mL离心管中,密封保存,待HPLC分析。每个处理重复3次。
1.3..2.2 HPLC分析 采用Kim等的方法[13-14]。利用反向C18色谱柱(5 μm,100A,4.6 mm×250 mm)进行试验,流动相A为水,B为乙腈;流速为0.8 mL/min;检测波长为 230 nm,柱温为30 ℃,进样体积为20 μL。二元梯度洗脱条件:B泵浓度在0~30 min为15%~55%,30~35 min为55%~100%,35~40 min为100%~15%,40~45 min为15%。
1.3.2.3 毛蕊异黄酮葡萄糖苷产量 毛蕊异黄酮葡萄糖苷产量=毛蕊异黄酮葡萄糖苷含量×生物量。
1.4 土壤温度和湿度
用土壤温湿度测定仪测定,探针深度为10 cm,取月平均值代表土壤温度和湿度。
1.5 统计分析
数据分析采用SPSS 19.0软件包,2个样本之间显著性差异利用独立样本t检验(P<0.05),相关性分析利用双变量相关分析中的皮尔森相关系数(Pearson correlation coefficient,简称Pearson)。
2 结果与分析
2.1 不同生长时期膜荚黄芪的生物量
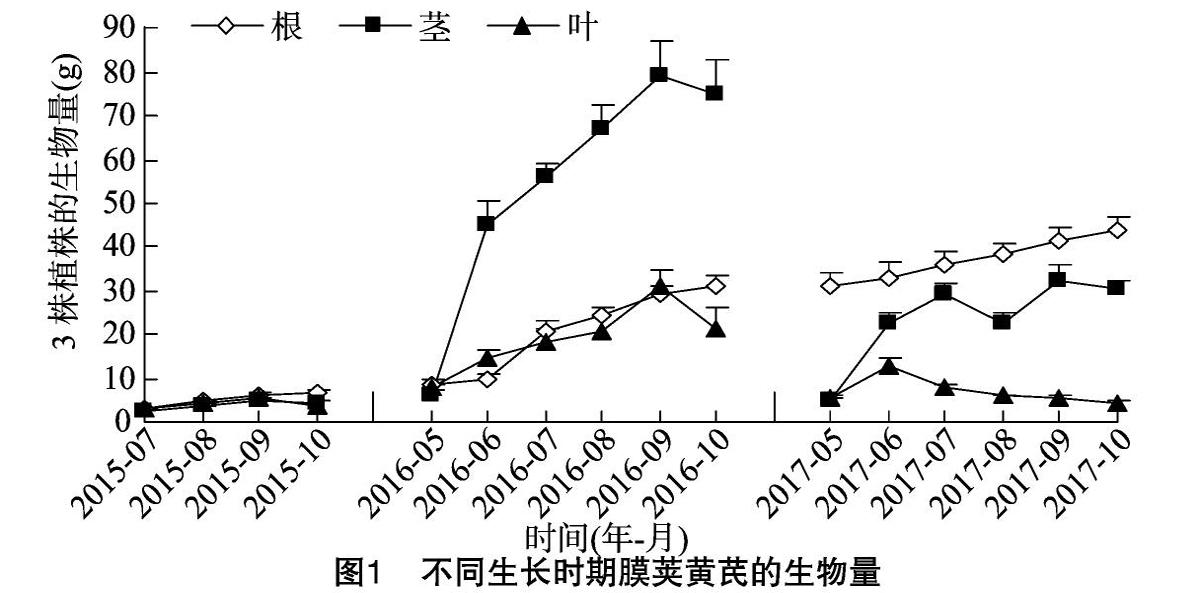
在本研究过程中,随着膜荚黄芪的自然生长,1、2年生黄芪几乎没有发生病虫害,而3年生黄芪在5月末受到蚜虫侵害,6月初发生白粉病,8月初开始受根腐病危害,其生物量的动态变化如图1所示。随着植株生长,根生物量呈持续增长趋势,1、2年生积累速度较快,3年生积累速度开始减缓,这可能与3年生黄芪发生病虫害有关;1、2年生茎生物量至9月份为止呈上升趋势,10月份地上部开始枯萎时出现下降现象,而3年生茎生物量至9月份为止呈波动升高,随后降低;1、2年生叶生物量积累与茎相似,3年生叶生物量则在6月份明显增加后,一直呈降低趋势。2、3年生茎和叶的生物量在6月份快速积累,表明植株开始进入旺盛生长期。总体而言,1、2年生黄芪的生物量随着植株生长逐渐积累,而病虫害的发生严重限制了3年生黄芪生物量的积累。
2.2 毛蕊异黄酮葡萄糖苷含量
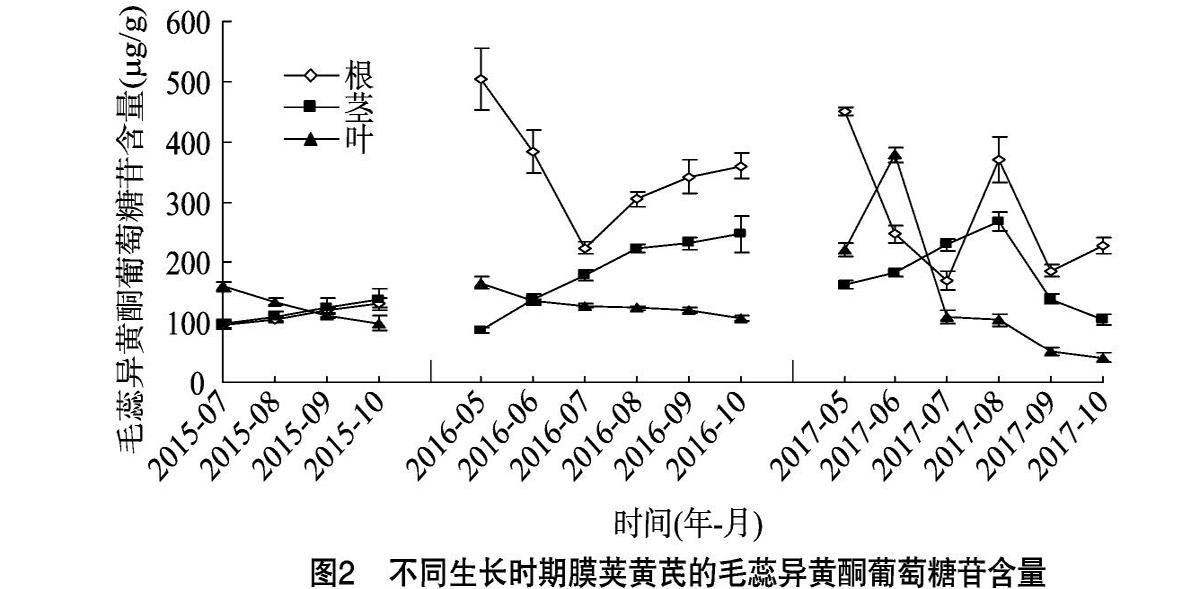
毛蕊异黄酮葡萄糖苷在膜荚黄芪自然生长中的各个时期、各个部位(根、茎和叶)均有分布(图2)。其含量在1年生根中呈增长趋势(7—10月),在2年生根中呈先降低(5—7月)后升高(8—10月)趋势,在3年生根中除8月明显增加之外,变化趋势与2年生相似,这可能与3年生根在8月受根腐病刺激有关;1、2年生茎中含量一直保持上升趋势,3年生茎中含量到8月为止也一直呈上升趋势,之后开始下降;1、2年生叶中含量呈持续下降趋势,3年生叶中除6月急剧增加之外,也一直保持下降趋势。总而言之,1、2年生黄芪中毛蕊异黄酮葡萄糖苷积累规律相似,而病虫害的发生干扰了3年生黄芪中较为规律的积累模式。
2.3 土壤温度和湿度对根毛蕊异黄酮葡萄糖苷含量的影响
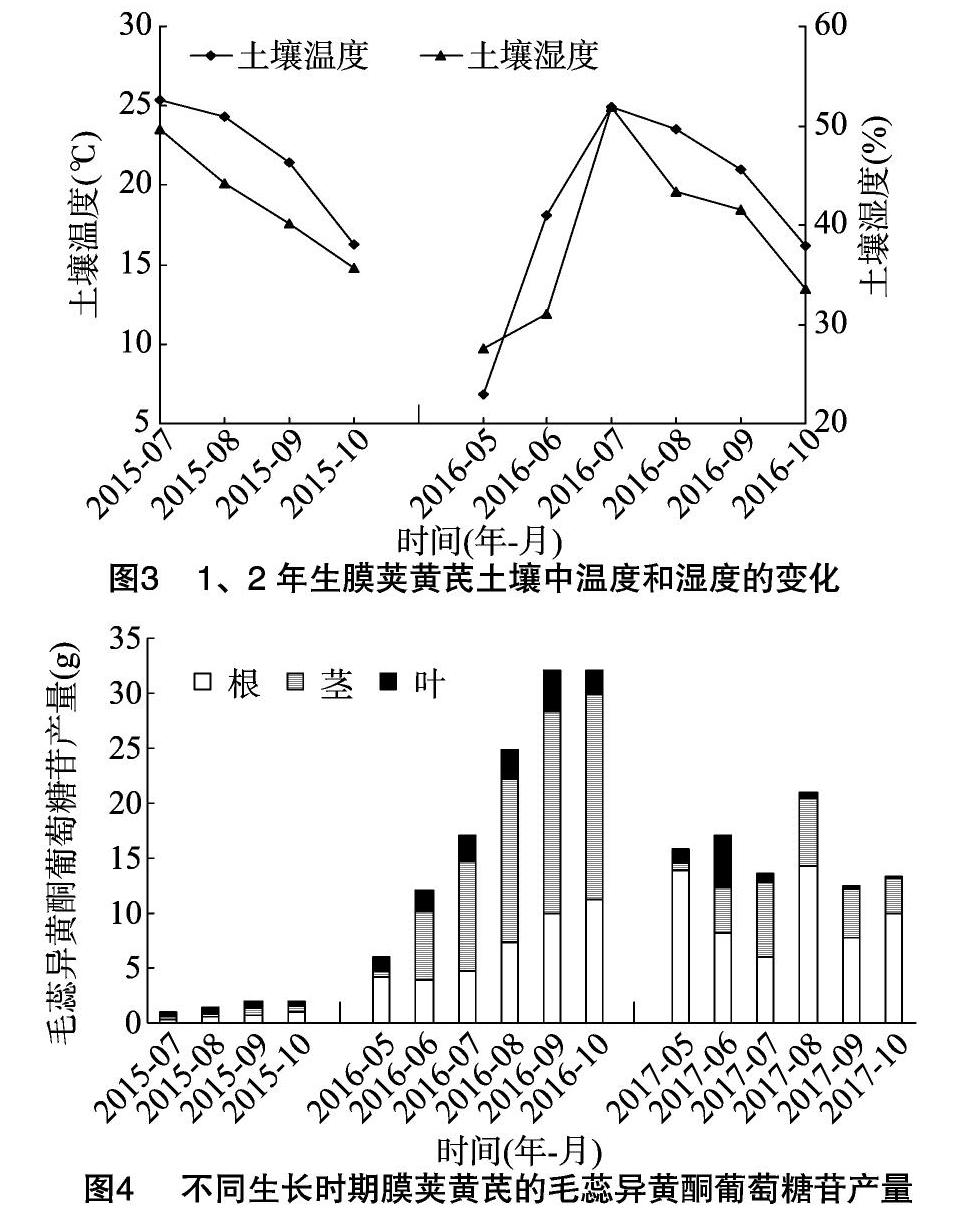
由于3年生黄芪受病虫危害,本研究只分析了土壤温度和湿度对1、2年生黄芪根中毛蕊异黄酮葡萄糖苷含量的影响。由图3可知,黄芪土壤温度和湿度变化趋势与根中毛蕊异黄酮葡萄糖苷的含量变化趋势(图2)相反。统计分析结果显示,土壤溫度和湿度与1年生根中毛蕊异黄酮葡萄糖苷含量之间的Pearson相关系数分别为-0.974(P<0.05)和 -0.988(P<0.05),与2年生根中毛蕊异黄酮葡萄糖苷含量之间的Pearson相关系数分别为-0.952(P<0.01)和 -0.931(P<0.01),这表明土壤温度和湿度与毛蕊异黄酮葡萄糖苷含量呈显著负相关。
2.4 毛蕊异黄酮的葡萄糖苷产量
由图4可见,毛蕊异黄酮葡萄糖苷总产量(包括根、茎和叶)在1、2年生黄芪中整体呈上升趋势,10月时最高,而病虫害的发生则抑制了3年生黄芪总产量增长。值得一提的是,除5月外,2年生茎中的产量明显高于同时期根和叶中的产量。
3 讨论
毛蕊异黄酮葡萄糖苷是黄芪中主要的活性成分之一,由本研究结果可知,其组成型表达发生在膜荚黄芪生长的各个部位和各个时期, 这表明它在膜荚黄芪的生长发育过程中起重要的作用。在不发生病虫害时,无论是1年生还是2年生黄芪,随着植株的生长和生物量的积累,毛蕊异黄酮葡萄糖苷含量在茎中呈上升趋势,在叶中则保持下降趋势,说明地上部毛蕊异黄酮葡萄糖苷含量受植物本身生长发育水平的调控,
在嫩叶和成熟茎中含量较高。特别是2年生成熟茎(8、9、10月)中毛蕊异黄酮葡萄糖苷含量符合《中华人民共和国药典》规定的标准[1](>0.02%,即>200 μg/g),产量高,可成为获取毛蕊异黄酮葡萄糖苷的有效途径。
根中毛蕊异黄酮葡萄糖苷的含量与土壤温度和湿度呈显著负相关,且地上部枯萎后,土壤中的根经过寒冷漫长的冬季和寒冷干燥的春季,其毛蕊异黄酮葡萄糖苷的含量(2、3年生5月)与上一年(1、2年生10月)相比明显增加。这些结果表明,根中毛蕊异黄酮葡萄糖苷含量的积累与黄酮类化合物在植物的抗寒和抗旱过程中的保护作用密切相关[15],即适当的低温和水分胁迫促进了毛蕊异黄酮葡萄糖苷的积累。Pan等也报道,低温(2 ℃)刺激了蒙古黄芪幼苗根中毛蕊异黄酮葡萄糖苷的积累[7],并且这种积累提高了蒙古黄芪幼苗的耐冻性[16]。还有报道表明,适当的干旱胁迫促进了3年生蒙古黄芪根中毛蕊异黄酮葡萄糖苷的积累[17]。
在黄芪植株生长过程中,病虫害的发生是不可避免的。由本研究结果可知,进入快速生长期之后,病虫害的发生严重限制了3年生黄芪生物量的积累,这与杨春清等报道的黄芪白粉病、根腐病和蚜虫等病虫害的发生会造成黄芪大幅度减产的结果[18]相一致。异黄酮是豆科植物中的植保素[19],异黄酮的含量与大豆抗蚜虫[20]、红三叶抗白粉病[21]和大豆抗根腐病[19]的能力密切相关。与此相似,在本研究中蚜虫与白粉病(6月的叶)、根腐病的发生之初(8月的根)分别诱导了黄芪叶和根中毛蕊异黄酮葡萄糖苷的明显积累,但是长期的病虫危害则降低了毛蕊异黄酮葡萄糖苷的含量和产量。
根是黄芪主要的入药部位,在本试验中,根中毛蕊异黄酮葡萄糖苷的积累是动态的,与非生物和生物胁迫密切相关,同时也具有时间特异性,这可能与膜荚黄芪长期适应延边地区的生长环境有关。因此,在比较不同地区膜荚黄芪毛蕊异黄酮葡萄糖苷含量时,应充分了解各自地区毛蕊异黄酮葡萄糖苷积累的差异性,以增加可比性。
综上,本试验结果表明,2年生膜荚黄芪根中毛蕊异黄酮葡萄糖苷含量均符合《中华人民共和国药典》规定的标准,可进行适时采收;成熟茎中含量和产量都较高,可用于开发饲料;嫩叶中含量也不低,适合作茶饮,这些结果将有助于指导延边地区黄芪的栽培和生产。
参考文献:
[1]国家药典委员会.中华人民共和国药(一部)[M]. 北京:中国医药科技出版社,2015.
[2]张亚洲,徐 风,梁 静,等. 蒙古黄芪中异黄酮类化学成分研究[J]. 中国中药杂志,2012,37(21):3243-3247.
[3]李延勋,栗章彭,颜世利,等. 膜荚黄芪化学成分研究[J]. 中草药,2017,48(13):409-411.
[4]Ma C H,Wang R R,Tian R R,et al. Calycosin 7-O-β-D-glucopyranoside,an anti-HIV agent from the roots of Astragalus membranaceus var. mongholicus[J]. Chem Nat Compd,2009,45(2):282-285.
[5]Jian J,Sun L,Cheng X,et al. Calycosin 7-O-β-D-glucopyranoside stimulates osteoblast differentiation through regulating the BMP/WNT signaling pathways[J]. Acta Pharm Sin B,2015,5(5):454-460.
[6]Tang D,He B,Zheng Z G,et al. Inhibitory effects of two major isoflavonoids in Radix Astragali on high glucose-induced mesangial cells proliferation and AGEs-induced endothelial cells apoptosis [J]. Planta Medica,2011,77(7):729-732.
[7]Pan H,Fang C,Zhou T,et al. Accumulation of calycosin and its 7-O-β-D-glucoside and related gene expression in seedlings of Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao induced by low temperature stress[J]. Plant Cell Reports,2007,26:1111-1120.
[8]宋成英,封加福. HPLC同時测定黄芪药材中毛蕊异黄酮葡萄糖苷和黄芪甲苷[J]. 中国实验方剂学杂志,2013,19(11):115-117.
[9]张贺廷,王 健,程铭恩,等. 蒙古黄芪主产区栽培及商品规格等级调查[J]. 中药材,2015,38(12):2487-2492.
[10]秦雪梅,何 盼,李震宇,等. 黄芪的名称考证[J]. 中药材,2014,37(6):2487-2492.
[11]张艺莲. 延边中草药材的资源及其利用情况[J]. 中国民族民间医药杂志,2001(52):285-289.
[12]李 波,肖井雷. 延边朝鲜族自治州中药资源调查研究[J]. 吉林中医药,2015,35(8):823-825.
[13]Kim G S,Lee D Y,Lee S E,et al. Evaluation on extraction conditions and HPLC analysis method for bioactive compounds of astragali radix[J]. Korean J Medicinal Crop Science,2013,21(6):486-492.
[14]李子羊,刘 佳,孙海燕,等. IBA浓度对膜荚黄芪不定根生物量、毛蕊异黄酮及毛蕊异黄酮葡萄糖苷积累的影响[J]. 延边大学农学学报,2016,38(3):209-213.
[15]Akula R,Ravishankar G A. Influence of abiotic stress signals on secondary metabolites in plants[J]. Plant Signal Behavior,2014,6(11):1720-1731.
[16]Pan H,Li X,Cheng X,et al. Evidence of calycosin-7-O-β-D-glucosides role as a major antioxidant molecule of Astragalus membranaceus Bge. var. mongholicus (Bge.) Hsiao plants under freezing stress[J]. Environmental& Experimental Botany,2015,109:1-11.
[17]李光跃,罗晓雅,孙窗舒,等. 干旱胁迫对黄芪植株生长中黄酮类成分积累的影响[J]. 西北植物学报,2017,37(1):138-143.
[18]杨春清,孙明舒,丁万隆. 黄芪病虫害种类及为害情况调查[J]. 中国中药杂志,2004,29(12):67-70.
[19]张淑珍,王维峰,西 芳,等. 杨庆凯大豆抗疫霉根腐病机制的研究进展[J]. 大豆科学,2001,20(4):290-294.
[20]李 娜,于希森,李 琪,等. 大豆异黄酮合成关键酶基因对蚜虫取食的防御响应分析[J]. 大豆科学,2016,35(5):800-804.
[21]宋 超,靳晓丽,田新会,等. 不同红三叶品种生产性能及异黄酮含量的比较[J]. 草原与草坪,2012,32(5):47-52.张梓豪,李 明,郑扬波,等. 硝酸铜对白木香细胞生长及抗氧化酶活性的影响[J]. 江苏农业科学,2019,47(5):106-109.
猜你喜欢
今日农业(2021年7期)2021-11-27
今日农业(2021年12期)2021-10-14
中老年保健(2021年5期)2021-08-24
天津农林科技(2020年3期)2020-08-13
河北果树(2020年1期)2020-02-09
今日农业(2019年15期)2019-09-03
现代园艺(2017年22期)2018-01-19
现代园艺(2017年13期)2018-01-19
现代园艺(2017年11期)2017-06-28
中国农业文摘-农业工程(2016年5期)2016-04-12
