
两种地衣共生藻对铜和锌的耐受性及吸附特性研究
2019-07-26 09:10吉米拉木加马力美合日班阿不力米提古海尼沙买买提艾尼瓦尔吐米尔
生物技术通报 2019年6期
吉米拉木·加马力 美合日班·阿不力米提 古海尼沙·买买提艾尼瓦尔·吐米尔
(新疆大学生命科学与技术学院,乌鲁木齐 830046)
自然环境中铜和锌元素对生物起着双重作用。锌是各种藻类生长代谢的重要微量元素[1-2],但是高浓度的锌离子通过抑制细胞分裂、影响光合作用等方式抑制藻类的生长,从而使得叶绿素含量下降[1]。锌离子在水溶液中的浓度达到1.0-1.4 mg/L时,导致藻类的直接死亡[3]。低浓度铜是藻类代谢过程中必不可少的,但高浓度铜对藻类具有毒害性。铜对细胞代谢具有极强的抑制作用,故在生产生活中常把硫酸铜配成杀藻剂,有效控制水环境藻类的大量生长[4]。
随着农业、工业等的不断发展,汞、镉、铅、镍、铜、锌等重金属污染对水生态环境、生物生存及人类健康造成威胁,已成为人类面临解决的迫切难题之一。如今,多种生物体包括细菌、真菌、酵母、藻类以及大型植物等作为生物吸附剂,被用来吸附水环境中的重金属。与其他生物吸附剂相比,藻类对诸多重金属具有很好的吸附能力,资源丰富,不仅是水体重金属污染检测剂,更重要的作用是作为生物吸附剂,可以广泛地应用在已被重金属污染并且其他的生物都很难以生存的水体的改造。1949年Ruchhoft等首次提出了“生物吸附”的概念[5];1988年,Gekeler等[6]研究指出,藻类能够通过合成螯合肽复合物与重金属结合,并发现小球藻合成的螯合肽复合物可以结合Cu、Pb、Zn、Ag和Hg等多种重金属;2001年,浩云涛等[7]通过分离筛选得到的具高重金属抗性椭圆小球藻(Chlorellae elipsoidea)来研究其对不同浓度的Cu2+、Cd2+、Ni2+和Zn2+的吸收富集能力,结果发现,小球藻对4种重金属有较高的去除能力,去除率分别达到40.93%、98.33%、97.62%和86.88%;2010年,吴海一等研究指出:鼠尾藻对Zn2+和Cd2+的富集能力较强,修复重金属污染的海洋水环境时,尤其是锌的污染,可以利用鼠尾藻[8]。2011年,薛培英等[9]以黑藻为吸附剂,对重金属进行吸附研究,结果显示黑藻对Cu2+的最大吸附容量为42.86 mg/g,去除率达70%;2011年,Zakhama等[10]研究报道,石莼对Cu2+、Pb2+、Cd2+和Ni2+的吸附量分别为112 mg/g、230 mg/g、127 mg/g和 67 mg/g,石莼能有效地去除水环境中部分重金属离子。在国内,2015 年热依拉等[11]研究指出,不同浓度(1-4 mmol/L)Cu2+处理对地卷共生藻细胞结构无明显的影响,在所有处理条件下,地卷共生藻细胞壁、细胞膜均完整,且大多数共生藻细胞处于分裂状态。有些地卷共生藻及其内生真菌可能本身就具有对Cu2+、Zn2+等重金属离子的耐性,在地衣共生藻抗重金属Cu2+、Zn2+胁迫中与共生菌可能同等重要[12-15]。
为了深入了解地卷共生藻响应重金属Cu2+、Zn2+胁迫的耐受性吸附特性和地衣共生藻在水体重金属污染修复中是否存在一定的优势,本研究以2种地衣共生藻为研究对象,在 Cu2+、Zn2+胁迫下,研究地衣共生藻对Cu2+、Zn2+的吸附性,为进一步探讨地衣共生内藻在水体中重金属污染生物修复中的实际应用提供理论依据。
1 材料与方法
1.1 材料
从两种地衣体即平盘软地卷(Peltigera elisabethae),犬地卷(Peltigera canina)内分离培养的地衣共生藻(2种杆裂丝藻)分别命名为:B和P.E。
地衣标本:2014 年 10 月采集于乌鲁木齐米泉林场哈熊沟,海拔高度为 1 960-2 100 m,-20℃保存。
1.2 方法
以分离纯化的两种地衣共生藻(B、P.E)为实验材料,配制含重金属BG-11液体培养基及藻株悬浮液,Cu2+(20、40、60、80、100 mg/L),Zn2+(100、200、300、400、500 mg/L)对2种地衣共生藻进行培养12 d,隔3 d对各种指标(细胞活率,重金属吸附特性)进行一次测定。
1.2.1 相关实验指标的测定 细胞存活率的测定:采用伊文斯蓝染色法计算细胞存活率[16],进一步用半数致死浓度计算方法[17]来计算并分析细胞半致死浓度(LC50)。
Cu2+、Zn2+的吸附特性:配制含重金属BG-11液体培养基及藻株悬浮液,对5种地衣体藻进行培养6 d,并通过离心(8 000 r/min,10 min),去除上清液用于测定培养液重金属含量。将收集的藻体用5 mL 的 20 mmol/L Na2-EDTA 反复冲洗 10 min,然后用 5 mL的去离子水冲洗10 min,2次冲洗的溶液混合在一起用吸附于细胞表面的重金属含量测定。采用BCO分光光度法测定铜含量[18],双硫腙分光光度法测定锌含量[19]。,并按照公式计算藻细胞对重金属的去除率:Q=(C0- C1-C2)/C0×100%。
公式中:Q为藻细胞对重金属的去除率,C0为培养液中的重金属初始浓度,C1为培养期结束后培养液中的重金属含量,C2为细胞表面冲洗取得重金属含量。
1.2.2 数据处理 采用 Excel 2010 及 SPSS 22.0 软件进行数据处理与方差分析和多重比较(0.05 和 0.01水平)。
2 结果
2.1 Cu2+胁迫对2种地衣共生藻细胞活率的影响
通过伊文思蓝染色剂检测地衣共生藻的死亡,死亡细胞着色深蓝色,而活细胞不着色(绿色),可明显地区别活细胞和死亡细胞,是一种简便、直观的检测方法(图1)。

图1 两种地衣共生藻伊文斯兰染色结果图(10×40)
如图2所示,在不同浓度胁迫下(0-100 mg/L),2种地衣共生藻(B、P.E),随着 Cu2+浓度的提高及培养时间的延长(0,3,6,9,12 d),2种地衣共生藻B与P.E细胞存活率均逐步下降(P<0.01),但具有一定的差异。
较低的Cu2+胁迫下(≤60 mg/L时),2种地衣共生藻均保留着较高的细胞活力,培养至12d时,仍然存活一半多的细胞,Cu2+胁迫对2种地衣共生藻的毒性作用较低;随着Cu2+浓度的加剧,共生藻细胞的活力明显梯度式降低,当Cu2+胁迫100 mg/L时,地衣共生藻P.E培养至第6d,56.66%的藻细胞可存活,但是培养至9 d、12 d时,藻细胞基本都死亡(32.48%、10.43%);地衣共生藻B培养初期(3 d)细胞活力较明显的下降,达12d时,只有 6.43%细胞可存活,几乎没有生存的细胞。结果表明,较高浓度Cu2+胁迫(80-100 mg/L),对2种地衣共生藻的影响均较显著,但是各浓度胁迫下,地衣共生藻P.E的细胞活力均高于B,对Cu2+胁迫有较高的耐性。

图2 不同浓度Cu2+胁迫对2种藻细胞活率的影响
2.2 Zn2+胁迫对两种地衣共生藻细胞活率的影响
如图3所示,不同浓度的Zn2+(0-500 mg/L)胁迫下,2种共生藻B和P.E的细胞存活率与随着胁迫浓度及培养时间呈反相关,即随着胁迫浓度的提高及培养时间的延长呈逐步下降趋势(P<0.01),但变化趋势存在一定的差异。
较低浓度Zn2+(≤200 mg/L)胁迫下,2种地衣共生藻细胞均保留着较高的活力,并且较稳定(细胞活力>80%);当Zn2+浓度300 mg/L培养第12天时,地衣共生藻B细胞存活率为61.49%,仍然保留着较高的细胞活力,P.E 的细胞活率分为49.99%(接近50%);较高浓度的Zn2+胁迫(>300 mg/L)对两种藻的影响较大,均出现明显的梯度式下降;当Zn2+浓度500 mg/L培养第12天时,2种共生藻B和P.E的细胞存活率分别为38.25%和26.47%,细胞基本死亡。
计算半致死浓度(LC50)来确定对重金属的耐受性是一种可靠并且方便快捷的方法。为了比较2种地卷共生藻对Cu2+、Zn2+的耐受性,基于2种重金属胁迫对细胞存活率的影响,进一步计算并分析3种地衣共生藻在不同浓度Cu2+、Zn2+胁迫下的半致死浓度。结果(表1)表明,Cu2+、Zn2+胁迫下,2种地衣共生藻半致死浓度均随着培养时间的延长而逐步下降(P<0.01);不同重金属胁迫下,2种地衣共生藻对Zn2+的LC50均高于Cu2+,说明2种地衣共生藻对Zn2+的耐性均高于Cu2+;在Cu2+胁迫下,地衣共生藻P.E各培养时期的LC50均高于B,地衣共生藻P.E对铜的耐受性高于B;Zn2+胁迫时,前段培养时间(3-9 d),地衣共生藻P.E对Zn2+胁迫有较高的耐性,但培养12 d时,半致死浓度急剧下降并低于B,时间对衣共生藻P.E的影响较大。
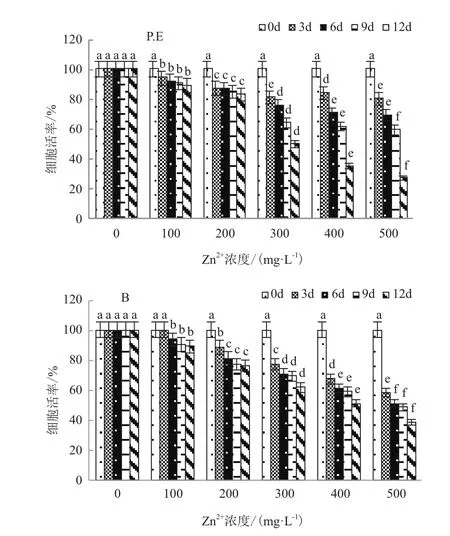
图3 不同浓度Zn2+胁迫对2种藻细胞活率的影响
2.3 两种地衣共生藻对Cu2+吸附特性研究
双环己酮草酰二腙(BCO)溶液中Cu2+浓度在波长(λ610nm)处与吸光度有良好的线性关系,以Cu2+浓度为横坐标,吸光度为纵坐标,绘制标准工作曲线,其标准方程为y=0.0955x-0.0065,相关系数为0.9995。
细胞内(表2),在不同浓度Cu2+胁迫下,2种地衣共生藻所吸附的铜含量与Cu2+胁迫浓度呈正相关,并随着时间的延长而逐步增加(P<0.01),但吸附量有所差异;各浓度Cu2+胁迫下(0-120 mg/L),地衣共生藻P.E在不同时间内所吸附的铜含量均高于B。

表1 两种地衣共生藻在Cu2+,Zn2+胁迫的半致死浓度(LC50)(mg·L-1)

表2 不同Cu2+浓度胁迫下2种地衣共生藻细胞内铜含量的变化
细胞外(表3),在不同浓度Cu2+胁迫下,藻培养液的铜含量变化有所差异却不同;当Cu2+胁迫下培养3 d时,2种地衣共生藻B和P.E培养液的铜含量随着胁迫浓度的提高呈先增加后下降趋势(P<0.01),当Cu2+浓度100 mg/L时,两种地衣共生藻对培养液Cu2+的去除率分别为88.61%和92.35%;培养第6天时,随着胁迫浓度的提高而增加(P<0.01),但对Cu2+去除率有所下降,当Cu2+浓度100 mg/L时,B和P.E对Cu2+去除率为85.67%,84.23%。

表3 不同Cu2+浓度胁迫下2种地衣共生藻培养液铜含量的变化
2.4 两种地衣共生藻对Zn2+吸附特性研究
双硫腙-吐温80溶液中波长(λ525nm)处的吸光度与Zn2+浓度具有良好的线性关系,其标准方程为y=0.0516x-0.0015,相关系数为0.9979。
细胞内(表4),不同浓度的Zn2+胁迫下,随着胁迫浓度的提高,2种地衣共生藻细胞所吸附的锌含量逐步增加(P<0.01);各浓度Zn2+胁迫下(0-500 mg/L),地衣共生藻P.E在不同时间内所吸附的铜含量均高于B,但地衣共生藻B所吸附的锌含量随着时间的延长而增加,地衣共生藻P.E则相反,随培养时间的延长而下降,时间对衣共生藻P.E的影响较大。
细胞外(表5),Zn2+胁迫下培养3d时,随着胁迫浓度的增加,培养液锌含量及2种地衣共生藻对Zn2+的去除率逐步增加(P<0.01),去除率分别为95.11%,95.05%,几乎相等;随着培养时间的延长(6d时),Zn2+浓度≤300 mg/L时,培养液的锌含量逐步增加,≥400 mg/L时差异不显著(P>0.05),当Zn2+浓度 500 mg/L,B和P.E对Zn2+的去除率为95.10%,92.23%,随着时间的延长地衣共生藻P.E对Zn2+吸附量有所下降,与地衣共生藻B几乎相等。
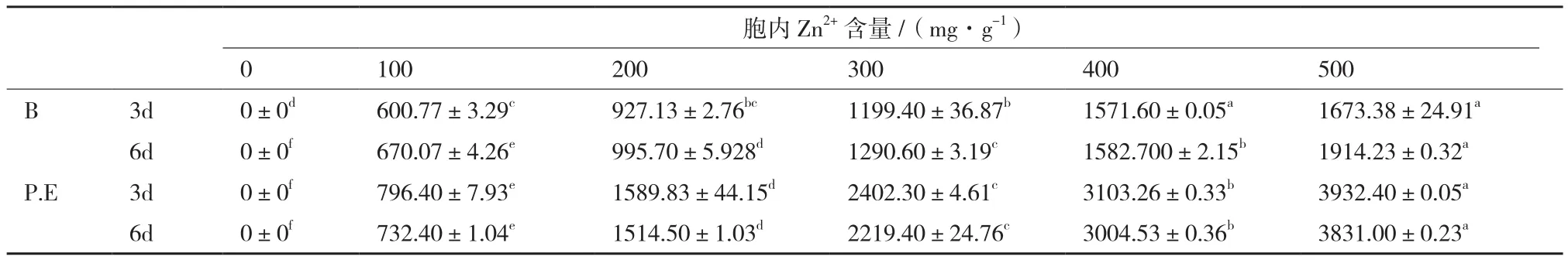
表4 不同Zn2+浓度胁迫下2种地衣共生藻细胞内锌含量的变化
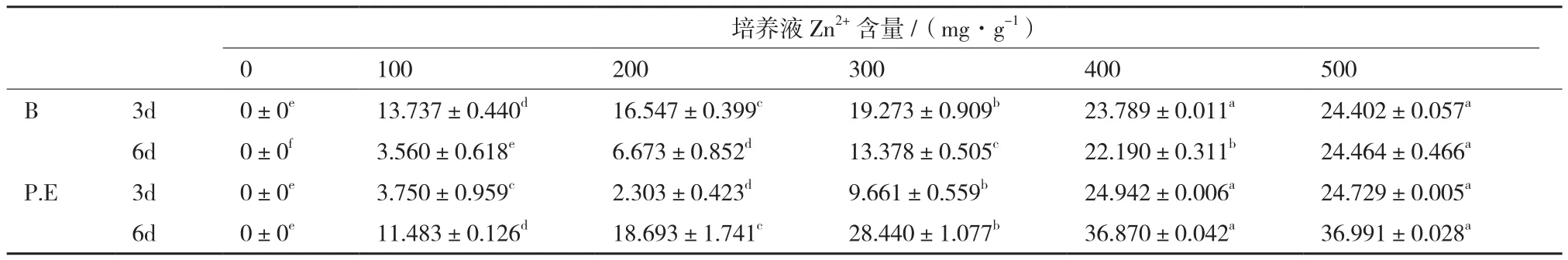
表5 不同Zn2+浓度胁迫下2种地衣共生藻培养液锌含量的变化
3 讨论
近年来有很多研究者利用水生生物来测定污染物在水中对生命体的毒性,而利用自由生长藻类来研究污染物的毒性及污染物的吸附性。藻中所含重金属的水平反映了藻类对水环境中重金属的生物利用率和对重金属的耐受性及吸附能力[20]。藻类与重金属离子有极强的亲和性,大多数研究聚焦于藻类对重金属离子的吸附特性[21-26],其机理包括络合、螯合、离子交换、转化、吸收和微沉淀等;金属阳离子可被细胞表面的负电荷位点所吸附细胞原生质膜外有明显的细胞壁,其在微生物吸附重金属离子的过程中起着重要作用,它是阻止金属离子进入细胞产生毒害的第一道屏障,并且不同藻类相对于不同重金属离子具有不同的耐性及吸附性。许多研究表明一些来自地衣体的藻类对重金属具有较高的耐受性及吸附特性。例如,2003年,Bačkor 等发现分离培养的地衣共生藻在4 mmol/L Cu2+胁迫下仍能正常生长,对铜具有较高的耐性[27];Álvarez等[28]也发现,分离培养的地衣共生藻Trebouxia在100 μmol/L Pb2+胁迫下,对铅有较高的吸附性。
为了探讨来自地衣体的藻类对不同重金属的耐受性及吸附特性,本研究以来自不同地卷属地衣的两种共生绿藻为研究对象,在不同浓度的铜和锌胁迫下处理不同时间,研究吸附于地衣共生藻细胞内外所吸收的铜、锌含量与细胞活力的影响,以确定2种地衣共生藻对铜和锌的耐受性与其铜吸附性差异及相关性。研究结果表明,2种地衣共生藻虽然属于同一个属,但对不同重金属的耐性及吸附性是有差异的;Cu2+对2种地衣共生藻的毒性作用高于Zn2+,对Zn2+的吸附性均高于 Cu2+,并且当不同浓度的Cu2+、Zn2+胁迫(3 d,6 d)时,地衣共生藻P.E的细胞活力及吸附率均高于B;可见地衣共生藻细胞活力与吸附性是有联系的,并呈正相关性。因此,研究2种地衣共生藻吸附于细胞内和细胞外Cu2+、Zn2+含量的变化及吸附能力的差异,可为解释地衣共生藻在响应重金属Cu2+、Zn2+胁迫时各自的作用或协同作用及其机制提供可参考的宝贵参考资料。
4 结论
(1)Cu2+、Zn2+是植物生长的必需元素,但在高胁迫浓度下,2种地衣共生藻对Zn2+耐受性及吸附性明显高于Cu2+,而且2种地衣共生藻对重金属的耐受性与对重金属的吸附性呈正相关性。
(2)来自菌藻共生的特殊生物-地卷属地衣的2种共生藻,相比一些自由生长藻类对铜和锌具有较高的耐受性和吸附特性,在水体重金属污染修复中是存在一定的优势,可进一步做一个良好的生物吸附剂材料。因此,本研究结果加深探讨环境重金属Cu2+、Zn2+污染的生物修复中地衣共生藻作为候选生物资源应用的可行性。
猜你喜欢
中国资源综合利用(2022年9期)2022-10-13
现代矿业(2022年3期)2022-04-09
当代水产(2021年8期)2021-11-04
潍坊学院学报(2020年2期)2021-01-18
军事文摘(2020年20期)2020-11-16
园林科技(2020年2期)2020-01-18
制造技术与机床(2019年9期)2019-09-10
今日农业(2019年10期)2019-01-04
天津科技大学学报(2016年1期)2016-02-28
中国医学科学院学报(2015年5期)2015-03-01
