
红松半同胞家系变异分析及选择研究
2019-07-19 10:30张秦徽王洪武姜国云王连奎李焱龙王立祥李月季赵曦阳
植物研究 2019年4期
张秦徽 王洪武 姜国云 沈 光 王连奎 李焱龙 王 雷 王立祥 李月季 李 蕊 赵曦阳*
(1.林木遗传育种国家重点实验室东北林业大学,哈尔滨 150040; 2.吉林省三岔子林业局林木良种基地管理中心,白山 134702; 3.白城市国有林场总场,白城 137000)
红松(Pinuskoraiensis)系五针松,为松科(Pinaceae)松属(Pinus)常绿乔木,国家二级重点保护野生植物[1],中国优质用材树种,主要分布于长白山及其北部的张广才岭、老爷岭、完达山和小兴安岭[2]。红松树形高大,干形通直,材质优良,易加工,广泛应用于家具制造、建筑等[3],是东北地区重要的用材树种;松籽味美,富含人体所需的多种营养物质及维生素,具有较高的食用和药用价值[4],因此也是我国重要的经济树种[5]。自“六五”开始,红松就被列为国家科技攻关计划课题支持树种,科研工作者们在优良种源选择[6]、林分选择[7]、无性繁殖[8]、种子园营建[9]、光合生理[10]、分子标记[11]等方面进行了广泛研究,并取得一定进展。
林木种子园是为生产优良种子、按人为设计要求所营建的特种人工林,最早可追溯到1880年[12]。我国红松种子园营建工作始于20世纪80年代,按种源区划选择种源或优树,以穗条嫁接的方式在红松分布区内营建了20多个红松无性系种子园,为后期红松的遗传改良提供了物质基础[13]。由于红松生长相对较慢,这些种子园目前大都处在初级种子园阶段,存在遗传增益低、种子产量不稳定等问题,不能满足东北林区生产用种的实际需要[14],因而需进行子代测定以及优良家系和单株选择来建立改良代种子园和二代种子园,以实现对红松的遗传改良。
研究生长性状的遗传变异是筛选优良群体及个体的重要内容。林木树种的生长性状研究较多,国内外均有报道。如Doran等人[15]用17个杨树无性系进行对比试验,对4年、6年和8年生的树高、胸径和材积进行了分析,估算遗传参数,进而筛选出应用前景较好的杨树无性系;张正刚等人[16]对7年生日本落叶松自由授粉家系的树高、胸径和冠幅等生长性状进行了测定,研究不同家系遗传变异,筛选出二代优树群体,作为日本落叶松高世代种子园的建园材料。本研究以吉林省三岔子林业局国家红松良种基地的53个28年生红松半同胞家系为材料,对其生长性状和形质性状的遗传变异进行分析评价,并选出优良家系和单株,以期为改良代种子园的营改建提供理论依据及造林生产应用提供优良种质材料,也为红松育种群体的建立及推广提供参考。
1 材料与方法
1.1 试验地点与材料
子代测定林位于吉林省三岔子林业局国家红松良种基地(E126°56′,N41°95′),该地海拔520 m,年均温2.5℃,年降水量725.5 mm,年日照时数2 300 h,无霜期110 d。试验材料共53个家系,其中52个家系来源于吉林省露水河林业局天然林母树林中的红松优树种子,1990年春季育苗,培育4年,苗龄型为S2-2,1995年春季选择试验地营建子代测定林。利用当地种子作为对照,具体编号见表1。试验地采用完全随机区组设计,6株小区,5个区组,株行距为2 m×3 m,周边设置保护行。
1.2 试验方法
于2017年10月对28年生的红松半同胞子代测定林的树高、地径、胸径、三米径、枝下高和第六轮枝高等生长性状及通直度、分枝角和轮枝数等形质性状进行全林调查。利用Vertex Ⅳ测高仪测量树高、枝下高和第六轮枝高;利用测树钢围尺测量地径、胸径和三米径;利用量角器测量分枝角;利用分级法[17]确定通直度。
利用实验形数法[18]计算材积(V):
V=(H+3)g1,3f
(1)
式中:红松平均实验形数f为0.33,H为树高,d为胸径,g1.3为胸高处横断面积。
方差分析线性模型为:
Xi=μ+Bi+F+Fiji+Meiji
(2)
式中:μ为总体平均值,Bi为区组效应,F为家系效应,Fiji为区组和家系的交互作用,Meiji为机误。
遗传力采用公式[19~20]:
家系遗传力(H2):
(3)
(4)

变异系数采用公式[21]:
表型变异系数(PCV):
(5)
遗传变异系数(GCV):
(6)

采用布雷金多性状综合评价法[22]对家系及家系内单株进行综合评定,采用公式:
(7)
式中:ai=Xij/Xjmax,Xij为某一性状的平均值,Xjmax为某一性状的最优值。
家系现实增益估算公式[23]:
(8)
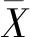
单株遗传增益估算公式[24]:
(9)
一般配合力采用公式[25]:
g=x-μ
(10)
式中,g为亲本的一般配合力,x为亲本的某个交配组合在某个性状的子代平均值,μ为这个性状所有组合的子代总平均值。
所有数据利用SPSS19.0软件进行分析[26]。
2 结果与分析
2.1 各性状方差分析
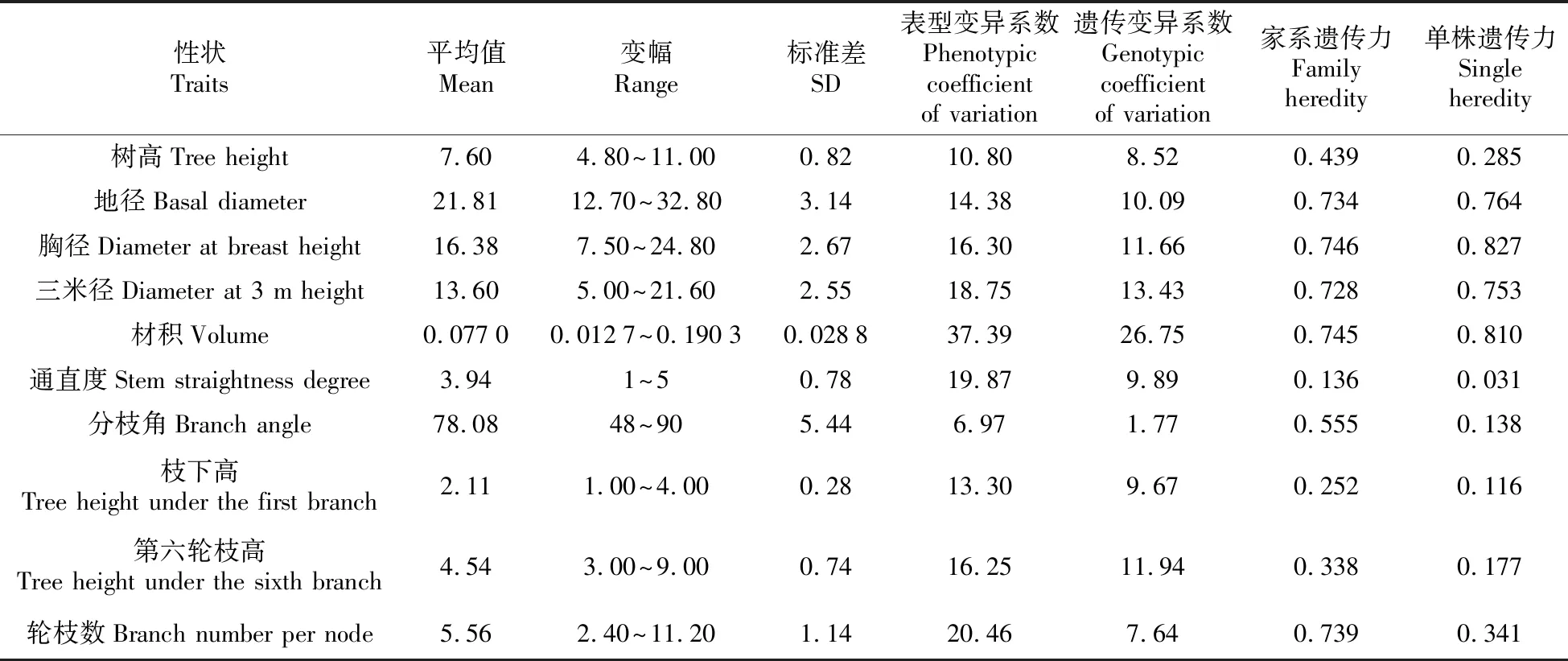
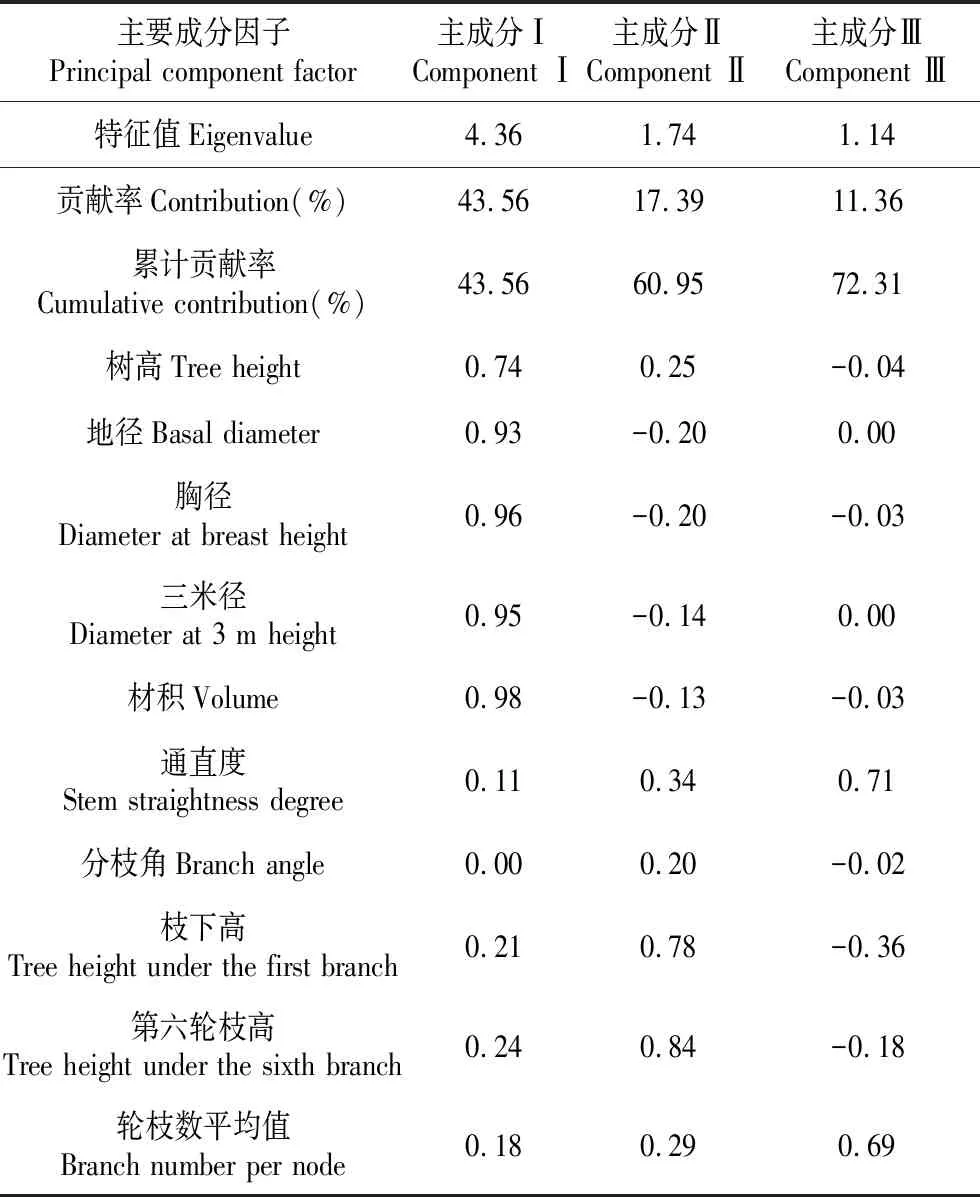
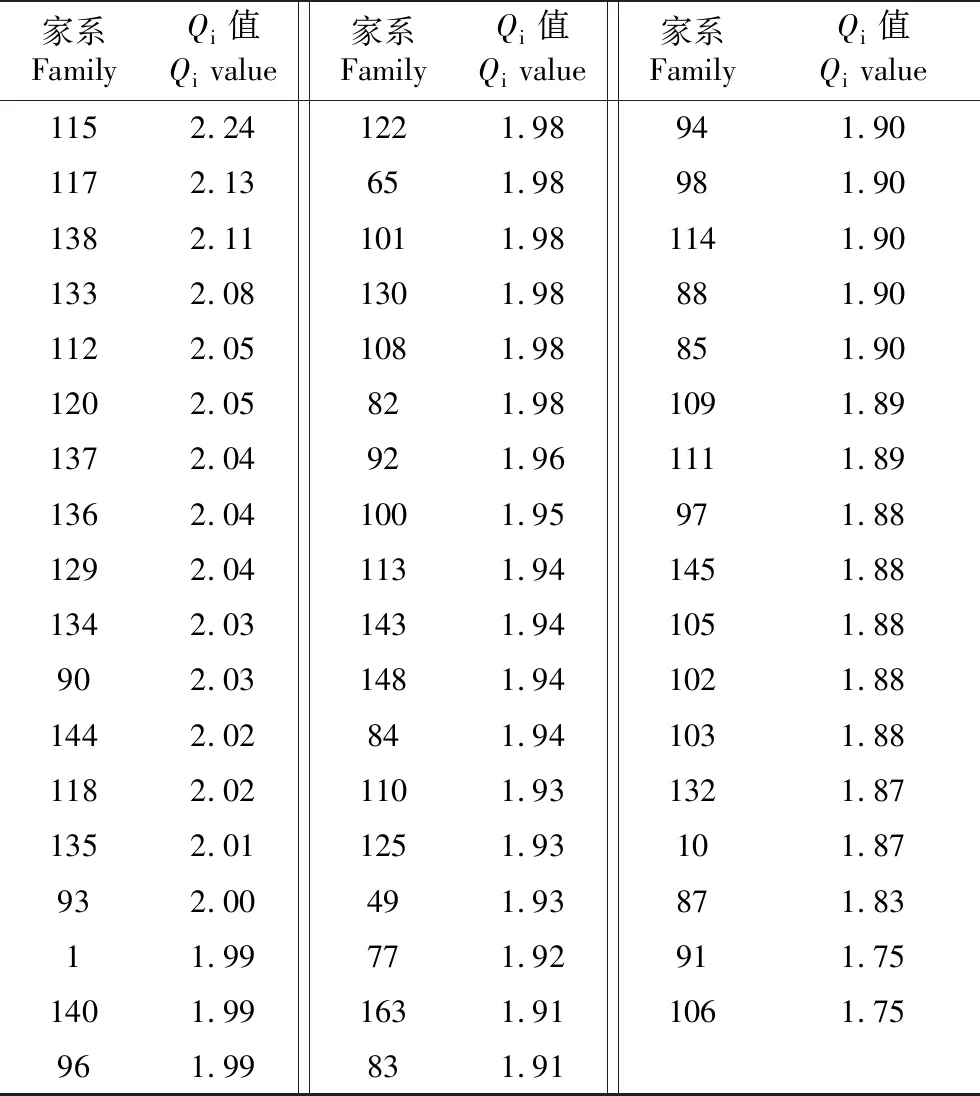
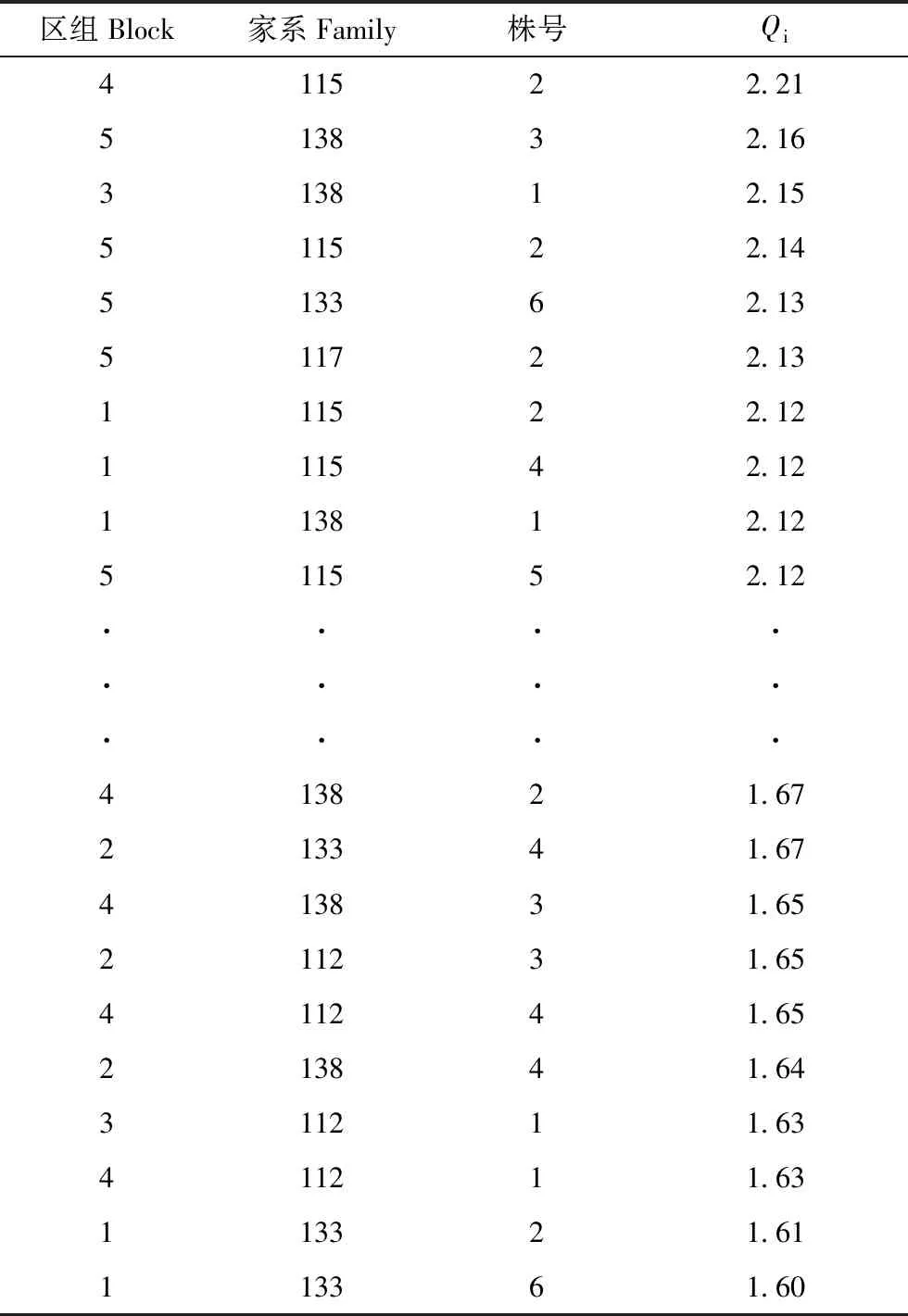
53个家系各性状方差分析结果见表2。结果显示,通直度在家系间、分枝角在区组间及家系和区组的交互作用间、枝下高在家系间以及轮枝数在区组间的差异未达显著水平(P>0.05);第六轮枝高在家系间以及轮枝数在家系和区组的交互作用间的差异显著(0.01 表2 不同家系各性状方差分析表 表3 不同家系各性状变异系数 53个家系树高、地径和胸径等5个生长性状及通直度、分枝角和轮枝数等形质性状变异参数见表3。所有家系树高平均值为7.60 m,变幅为4.80~11.00 m;地径的平均值为21.81 cm,变幅为12.70~32.80 cm;胸径的平均值为16.38 cm,变幅为7.50~24.80 cm;三米径的平均值为13.60 cm,变幅为5.00~21.60 cm;材积的平均值为0.077 0 m3,变幅为0.0127~0.1903 m3;通直度的平均值为3.94,变幅为1~5;分枝角平均值为78.08°,变幅为48°~90°;枝下高平均值为2.11 m,变幅为1.00~4.00 m;第六轮枝高平均值为4.54 m,变幅为3.00~9.00 m;轮枝数的平均值为5.56个,变幅为2.40~11.20个。 各性状表型变异系数变化范围为6.97%~37.39%,除分枝角外,其余各性状表型变异系数均超过10%;其中材积的表型变异系数最大,为37.39%。遗传变异系数变化范围为1.77%~26.75%。除通直度、枝下高和第六轮枝高外,其余各性状的家系遗传力较高,部分性状遗传力超过0.7。从单株遗传力来看,地径、胸径、三米径和材积的单株遗传力较大,均超过0.7。 各性状间相关系数见表4。由表4可知,从生长性状来看,树高、地径、胸径、三米径和材积各性状之间均达极显著正相关水平,其中地径、胸径、三米径和材积各性状间相关性较高,相关系数均超过0.9。从形质性状来看,通直度与分枝角达正相关(0.010),与轮枝数达极显著正相关(0.219);分枝角和轮枝数呈正相关(0.036)。对比生长性状和形质性状,通直度与树高、地径、胸径、三米径和材积均呈正相关关系,其中与树高和三米径达极显著正相关水平,与材积达显著正相关水平(0.054);分枝角与地径、胸径、三米径和材积表现为负相关,与树高表现为正相关(0.006);轮枝数与树高、地径、胸径、三米径和材积均表现为极显著正相关水平。 各性状一般配合力见表5。由表5可知,树高一般配合力变化范围为-0.598 6~0.971 4,地径一般配合力变化范围为-4.462 5~5.267 5,胸径一般配合力变化范围为-3.466 4~4.414 0,三米径一般配合力变化范围为-3.283 6~4.066 4,材积一般配合力变化范围为-0.030 6~0.054 7,通直度一般配合力变化范围为-0.602 8~0.497 2,分枝角一般配合力变化范围为-6.201 9~6.498 1,枝下高一般配合力变化范围为-0.167 2~0.269 6,第六轮枝高一般配合力变化范围为-0.398 9~0.907 8,轮枝数平均值一般配合力变化范围为-1.095 7~1.017 6。其中家系115各性状一般配合力均较高,其亲本可选作优良亲本材料。 各性状主成分分析结果见表6。由表6可知,主成分Ⅰ特征值为4.36,贡献率为43.56%,树高、地径、胸径、三米径和材积等性状特征值较高,分别为0.74、0.93、0.96、0.95和0.98;主成分Ⅱ特征值为1.74,贡献率为17.39%,枝下高和第六轮枝高等性状特征值较高,分别为0.78和0.84;主成分Ⅲ特征值为0.14,贡献率为11.36%,通直度和轮枝数平均值等性状特征值较高,分别为0.71和0.69。三个主成分的累计贡献率为72.31%,包含了供试家系生长及形质性状的大部分信息。 表4 不同家系各性状相关性分析 注:**极显著相关水平(P<0.01);*显著相关水平(P<0.05) Note:**means correlation is significant at the 0.01 level;*means correlation is significant at the 0.05 level 表6 各性状主成分分析 表7 不同家系Qi值 依据主成分分析结果,以树高、地径、胸径、三米径和材积等生长性状为选择指标对53个半同胞家系进行综合评价,各家系Qi值见表7。按家系材积现实遗传增益在35%以上,兼顾树高、地径、胸径和三米径对家系进行评价选择,家系115、117、138、133和112入选,入选率为10%,入选家系树高、地径、胸径、三米径和材积等性状平均值分别为7.90 m、25.02 cm、19.21 cm、16.23 cm和0.107 4 kg·m-3,现实增益分别为3.94%、14.71%、17.26%、19.34%和39.48%。 依据主成分分析结果,以树高、地径、胸径、三米径和材积等生长性状为选择指标对优良家系内的单株进行评价选择,各单株Qi值见表8。以单株材积遗传增益超过100%为标准,兼顾树高、地径、胸径和三米径对优良家系内单株进行评价选择,共获得6株优良单株,入选率为6%,入选优良单株树高、地径、胸径、三米径和材积平均值比总平均值分别高1.60 m、8.54 cm、7.04 cm、6.69 cm和0.097 kg·m-3,单株遗传增益分别为5.99%、29.92%、35.53%、37.06%和102.04%。 表8 不同单株Qi值 遗传和变异是林木育种研究的主要内容[27],对变异来源、特点和规律的认识是进行树种改良的重要基础[28]。本研究对28年生的红松优树子代半同胞家系的方差分析结果显示,家系间大部分性状均达极显著差异水平,表明母树林后代具有较大的分化[29],与王顺安等[30]对日本落叶松的子代分析结果一致,有必要进一步进行优良家系的评价选择。 变异系数是衡量群体变异程度大小的指标,它能反映树体各性状的遗传变异能力,包括表型变异系数和遗传变异系数[21]。变异程度高,有利于进行优良家系选择[31]。本研究中材积表型变异系数最大,达到37.39%;胸径表型变异系数为16.30%;树高的表型变异系数较小,为10.80%,这与李自敬对23年生长白落叶松研究结果较为相似[32],表明以材积作为选择指标进行家系选择更具有选择潜力。三个性状的遗传变异系数占表型变异系数的比例较大,均超过70%,比马尾松(Pinusmassoniana)研究结果高[33],说明家系间变异受遗传因素的影响较大,为优良家系的选择提供了基础。 遗传力表示亲本某一性状遗传给子代能力的大小,是估算遗传增益的重要参数[34]。本研究中树高、胸径和材积家系遗传力均高于宋云平等人对27年生红松的研究结果[35],部分性状的家系遗传力超过0.7,属高遗传力,表明所选材料各性状能较为稳定遗传,有利于进行优良家系的评价选择[36]。本研究中各性状的单株遗传力变化范围为0.031~0.827,其中树高单株遗传力为0.285,与张振等对25年和27年生的红松研究结果相似[37];胸径和材积的单株遗传力达到0.8以上,比侯义梅对15年生的日本落叶松研究结果偏高[38]。单株遗传力较高,可在家系内进行优良单株的筛选,为二代种子园的营建提供建园材料。本研究中各性状家系遗传力均高于单株遗传力,比Magnussen对白云杉(Piceaglauca)的研究结果偏高[39],表明各性状在单株水平上的遗传能力弱于家系,家系在各性状的遗传差异具有更稳定的遗传能力。 林木改良过程中,通常希望综合性状得到改良,所以性状之间相关性至关重要[40],相关性分析可以反映各性状之间存在的联系,为多性状联合控制育种提供参考[41]。本研究中的相关性分析结果显示,从生长性状来看,树高、胸径和材积等性状之间均达极显著正相关水平,这与Liang等对32年红松无性系研究结果相似[42],其中胸径与材积相关性较大,相关系数超过0.9,表明树高和胸径不是独立遗传的两个性状。从形质性状来看,通直度与分枝角呈正相关,表明树干的生长势强于树枝,树枝侧向生长。结合生长性状和形质性状,通直度与树高、三米径达极显著正相关水平,与材积达显著正相关水平,为长势和干型优良的优树选择提供依据。 配合力是林木育种工作的研究内容之一,其大小与具体性状有关,反映亲本优良性状传递给子代的相对能力[43]。本研究对53个半同胞家系各性状进行一般配合力分析,除家系115各性状一般配合力均较高,其亲本可作为优良亲本材料外,不同性状一般配合力高的家系差异较大,难以进行联合筛选,因而需结合主成分分析进一步对家系进行评价选择。 育种目标决定育种方向。红松是东北地区特有的经济用材树种,但因其生长缓慢,使得红松种子园升级换代较慢。为选育高生长、高出材量的优质红松资源,本研究通过主成分分析研究发现主成分Ⅰ中的树高、地径、胸径、三米径和材积等生长性状特征值较高,贡献率较大,且各性状间相关性较高,因此可作为筛选优质红松资源的评价指标。在进行家系选择时,入选率越大,遗传多样性丰富度越高,但遗传增益越小[44]。本研究以家系材积现实遗传增益在35%以上为期望目标,对家系进行评价选择,入选率为10%,入选家系树高、地径、胸径、三米径和材积等性状现实增益分别为3.94%、14.71%、17.26%、19.34%和39.48%,与梁德洋等人的研究结果相似[45],表明利用表型选择对红松进行遗传改良具有较好的效果,入选的家系在生长上存在明显的优势,若用优良家系进行生产造林,可减少林分工艺成熟时间,进而提高经济效益[41]。另一方面,筛选出的优良家系同时反映出对应亲本的优良特性,可进一步筛选出优良亲本,为1.5代种子园的营建提供理论支持。在优良家系选择的同时,还可进行优良单株的筛选。利用多性状综合评价法对优良家系内单株进行评价选择,按单株期望遗传增益在100%以上为标准对家系内单株进行筛选,获得6株优良单株,入选率为4%,入选单株树高、地径、胸径、三米径和材积等性状的平均值分别比总平均值高1.60 m、8.54 cm、7.04 cm、6.69 cm和0.097 kg·m-3,遗传增益分别为5.99%、29.92%、35.53%、37.06%和102.04%,高于张振对红松[36]和林能庆对马尾松[46]的研究结果,具有较大的遗传改良潜力,可作为红松优良种质材料推广使用。 随着林木改良工作日渐深入,在大力营建高世代种子园的同时,对初级种子园的改良和升级亦在进行。本研究的材料存在丰富的变异,所选的5个优良家系和6个优良单株生长及干型优势明显,各性状相关性强,遗传力高,家系间和家系内的个体间有较大的选择潜力,有利于红松的良种选育,为推广造林应用提供物质基础;根据子代测定结果可对亲本无性系进行重新选择,为初级种子园的去劣疏伐、1.5代种子园的改良营建提供理论参考;在优良家系内选出优良单株,可为二代种子园的营建提供建园材料;通过筛选得到的一般配合力高的亲本若进行杂交实验,还会提高子代杂种优势的机率,优良的个体也可用于无性化推广应用。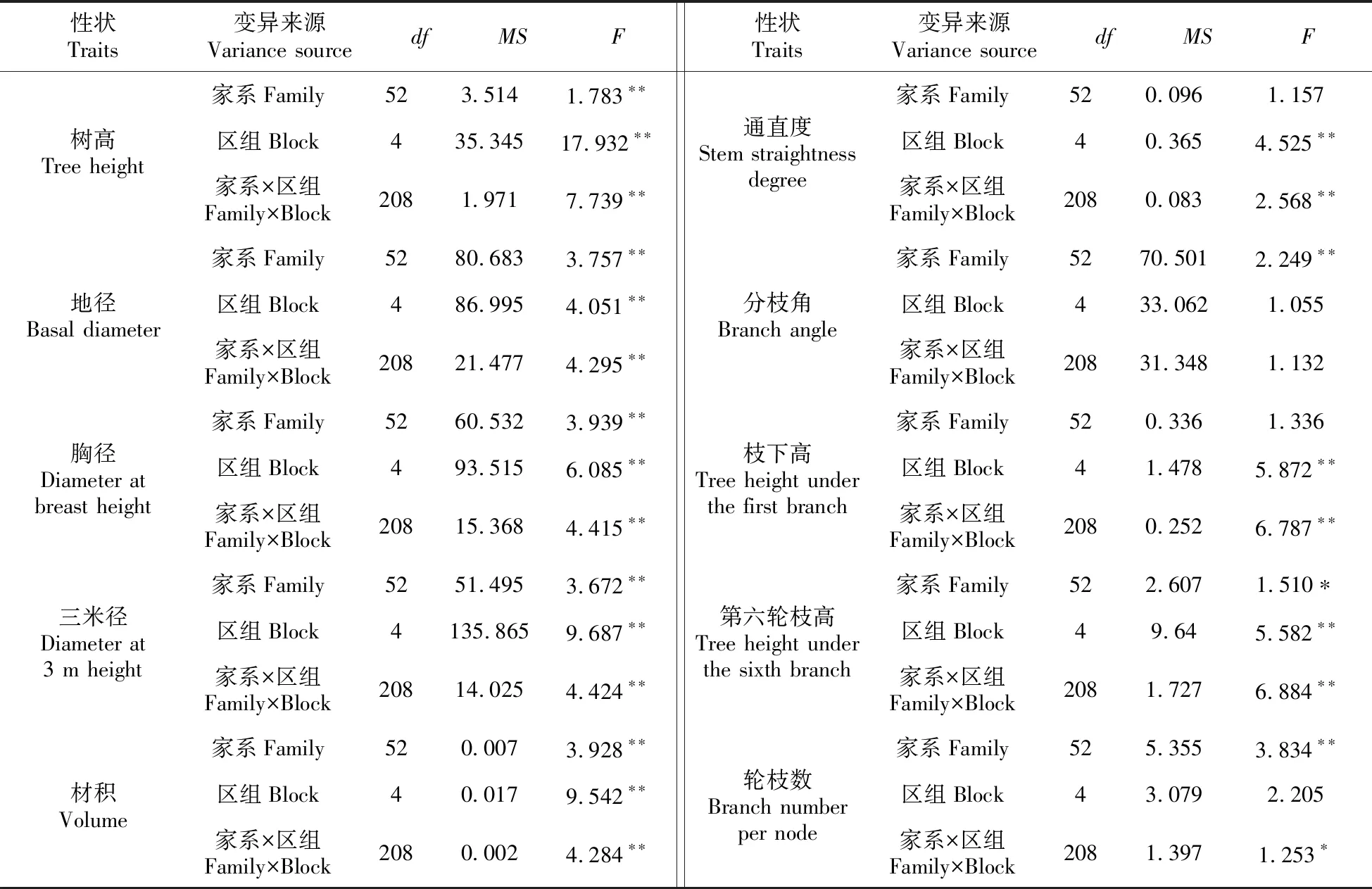
2.2 各性状遗传变异分析
2.3 各性状相关性分析
2.4 各性状一般配合力分析
2.5 各性状主成分分析

2.6 优良家系选择
2.7 单株选择
3 讨论
4 结论
猜你喜欢
浙江林业科技(2022年4期)2022-07-16
广西林业科学(2022年3期)2022-07-08
养殖与饲料(2021年11期)2021-11-15
园艺与种苗(2021年8期)2021-09-23
辽宁林业科技(2021年3期)2021-07-08
山西水土保持科技(2020年2期)2020-12-22
绿色科技(2016年23期)2017-03-15
园艺与种苗(2015年8期)2015-12-28
浙江林业(2015年4期)2015-12-21
卷宗(2012年5期)2012-10-21
